
Abstract
Objectives:
Bacteremia due to invasive Salmonella enterica has been reported earlier in children in Nigeria. This study aimed to detect the virulence and antibiotic resistance genes of invasive Salmonella enterica from children with bacteremia in north-central Nigeria.
Method:
From June 2015 to June 2018, 4163 blood cultures yielded 83 Salmonella isolates. This is a secondary cross-sectional analysis of the Salmonella isolates. The Salmonella enterica were isolated and identified using standard bacteriology protocol. Biochemical identifications of the Salmonella enterica were made by Phoenix MD 50 identification system. Further identification and confirmation were done with polyvalent antisera O and invA gene. Antimicrobial susceptibility testing was done following clinical and laboratory standard institute guidelines. Resistant genes and virulence genes were determined using a real-time polymerase chain reaction.
Result:
Salmonella typhi 51 (61.4%) was the most prevalent serovar, followed by Salmonella species 13 (15.7%), choleraesuis 8 (9.6%), enteritidis 6 (7.2%), and typhimurium 5 (6.1%). Fifty-one (61.4%) of 83 Salmonella enterica were typhoidal, while 32 (38.6%) were not. Sixty-five (78.3%) of the 83 Salmonella enterica isolates were resistant to ampicillin and trimethoprim-sulfamethoxazole, followed by chloramphenicol 39 (46.7%), tetracycline 41 (41.4%), piperacillin 33 (33.9%), amoxicillin-clavulanate, and streptomycin 21 (25.3%), while cephalothin was 19 (22.9%). Thirty-nine (46.9%) of the 83 Salmonella enterica isolates were multi-drug resistant, and none were extensive drug resistant or pan-drug resistant. A blaTEM 42 (50.6%), floR 32 (38.6%), qnrA 24 (28.9%), tetB 20 (20.1%), tetA 10 (10.0%), and tetG 5 (6.0%) were the antibiotic resistance genes detected. There were perfect agreement between phenotypic and genotypic detection of antimicrobial resistance in tetracycline, ciprofloxacin, and chloramphenicol, while beta-lactam showed κ = 0.60 agreement. All of the Salmonella enterica isolates had the virulence genes invA, sopB, mgtC, and sip4D, while 33 (39.8%), 45 (51.8%), and 2 (2.4%) had ssaQ, spvC, and ljsGI-1, respectively.
Conclusion:
Our findings showed multi-drug resistant Salmonella enterica in children with bacteremia in northern Nigeria. In addition, significant virulence and antimicrobial resistance genes were found in invasive Salmonella enterica in northern Nigeria. Thus, our study emphasizes the need to monitor antimicrobial resistance in Salmonella enterica from invasive sources in Nigeria and supports antibiotic prudence.
Introduction
The type of infection that results from Salmonella enterica is determined by the virulence factors of the bacterium and the host’s factors.1–3 Salmonella infection could manifest clinically as gastroenteritis (diarrhea, abdominal cramps, and fever) or a fatal febrile systemic infection (typhoid) that needs to be treated with antibiotics.4–7 Focal conditions and asymptomatic carriers are possible and are significant sources of continued infection transmission. 8 Salmonella is a gram-negative, flagellated, with O, H, and Vi antigens. More than 1800 Salmonella serovars are known.2,9 Salmonellae infection is possible when the bacteria gets through the gastric acid barrier, into the mucosa of the small and large intestines, and makes toxins. 10 Invasion of the epithelial cells causes the release of cytokines that cause inflammation. 11 It results in diarrhea, leading to ulceration and the destruction of mucosal cells. Also, if this spread in the intestines persists, it could result in systemic infection. 11
Horizontal gene transfers shape bacterial genomic diversity. 12 Some pathogens have genomic islands or islets (GI) with functionally linked genes.13,14 Salmonella pathogenic islands (SPIs) contain virulence genes, distinguishing them from nonpathogenic types.15–18 Virulence factors determine a host’s pathogenicity. 19 The adhesion and invasion of host cells by pathogenic S. enterica are aided by pef, spv, invA, and fim genes, respectively. Magnesium transport C (mgtC), Salmonella toxin (stn), and pip A, B, and D help the bacterium survive in the host system.20,21
The burden of invasive bloodstream infection due to S. enterica is increasing, especially in developing countries. 5 Globally, 1.2 million deaths attributable to S. enterica are recorded annually, with the vast majority occurring in resource-limited countries. 22 In resource-limited countries, non-typhoidal Salmonella infections cause bacteremia in immune-compromised and malnourished adults and children. 16 Most morality from Salmonella infection is connected to poor diagnostic infrastructures leading to misdiagnosis and drug misuse. 23 It is established that most pathogenic bacteria are acquired from the environment, food, and water sources.24,25 Although typhoidal Salmonella is a human host-adapted strain, recent literature has found typhoidal Salmonella in the food chain and water sources.26–30
The worldwide rise of multi-drug resistance is a major health concern. 31 It is becoming increasingly important to routinely apply antimicrobial susceptibility testing to select the antibiotic of choice and to screen for emerging multi-drug resistant (MDR) strains, 32 as several recent studies have reported the emergence of multi-drug resistant Salmonella pathogens from various origins, including humans, 33 birds, 34 cattle, 31 and fish. 35
Pathogens’ high antimicrobial resistance has been attributed to antimicrobial misuse in human and animal husbandry.36–39 Comparative genomic data from invasive Salmonella data and those from the environment and food chain have shown relatedness between clinical isolates and other sources.10,11,40 It has caused great concern as clinical S. enterica are resistant to the commonly used antibiotic, and some have been found to harbor extended-spectrum beta-lactamases (ESBL) genes which could make treatment difficult.41–46 Recent data from surveillance has found non-typhoidal Salmonellae to be highly resistant to antimicrobials.4,47–49 Thus, clinical care for individuals with invasive Salmonella infection is expensive, increases their hospital stay, and burdens them financially.50,51 Studies have been carried out on Salmonella virulence factors, but information on invasive isolates is scarce, especially from the pediatric population. This study aims to investigate the virulence and antimicrobial resistance genes pattern of S. enterica from invasive bloodstream infection in children from north-central Nigeria.
Materials and methods
Study design
This is a secondary cross-sectional analysis of isolated S. enterica in children with bloodstream infection in the Federal Capital Territory (FCT) and Nasarawa State, Nigeria.
Collection of Salmonella isolates analyzed in the study
Eighty-three gram-negative bacilli isolates were collected from blood cultures recovered from the study conducted at seven hospitals in FCT and Nasarawa State, Nigeria. Presumptively identified Salmonella isolates from four thousand and sixteen blood cultures were processed from June 2015 to June 2018. The study was part of Community-Acquired Bacteremia Syndrome in Young Nigeria Children conducted in north-central Nigeria from 2008 to 2018 by the International Foundation Against Infectious Diseases in Nigeria. The outcomes from 2008 to 2015 had been previously reported by Obaro et al., 5 and those previously reported isolates were excluded from this study.
Isolation of S. enterica of positive blood culture
Bacterial analysis, including gram staining and biochemical analysis using the analytical profile index (API20E) (Biomerieux, SA Lyon, France), was used to identify the Salmonella pathogens. Obaro et al. 5 have previously described the protocol used to culture and isolate the S. enterica used in this work. Briefly, all positive bottles were subcultured onto MacConkey agar (Oxoid, London, UK) and Salmonella shigella agar (Oxoid) plates and then incubated at 36°C for 24 h. The isolates were frozen at −80°C in 10% skim milk glycerol (Hardy Diagnostics, Santa Maria, California, USA) until used. 52 In conducting the study, the previously collected isolates were grown onto S. shigella agar (Oxoid) and incubated at 37°C for 24 h. In checking for Salmonella isolates, colonies were looked for on the plates. The morphological traits and characteristics of Salmonella species were selected, and gram staining of the selected colonies from each plate was examined. 52 Biochemical assays, including reactions on triple sugar iron agar, lysine iron agar, indole synthesis in tryptone broth, and urea splitting ability, were then conducted using the Phoenix MD (Beckon-Dickson systems, San Jose, California, USA). Molecular invA gene detection was used to validate the authenticity of the isolates. Polyvalent Salmonella antisera, A-G, A-S surface antigen, flagellar H (phase 1 and phase 2) (Beckon-Dickson systems) according to Kauffman-White Scheme 53 were used in the serotyping of the Salmonella isolates.
By Antimicrobial Susceptibility Testing and Multiple Antimicrobial Resistance (MAR) Index according to Clinical and Laboratory Standards Institute recommendations, 54 the antibiotic susceptibility of the isolates was determined. The antibiotic discs (ampicillin (10 μg), amoxicillin-clavulanate (30/10 μg), piperacillin (30 μg), piperacillin-tazobactam (30/10 μg), streptomycin (10 μg), trimethoprim-sulfamethoxazole (10/25 μg), chloramphenicol (30 μg), tetracycline (30 μg), aztreonam (30 μg), gentamicin (10 μg), amikacin (30 μg), cephalothin (30 μg), cefuroxime (30 μg), cefotaxime (30 μg), ceftazidime (30 μg), cefepime (30 μg), ceftriaxone (30 μg), levofloxacin (5 μg), meropenem (10 μg), imipenem (10 μg), tigecycline (25 μg), cefotaxime-clavulanate (30/10 μg), and ceftazidime-clavulanate (30/10 μg) that were utilized in a disk diffusion assay were from Oxoid. The BD PhoenixTM M50 system (Beckon-Dickson systems) was used for minimal inhibitory concentration (MIC) testing. For ciprofloxacin, MIC values >0.064 g/mL were viewed as reduced susceptibility, while MIC values 4 g/mL were interpreted as resistant; for azithromycin, MIC values >16 g/mL were interpreted as resistant. According to Davis and Brown, 55 the MAR indexes were derived as the ratio of antibiotics to which resistance was demonstrated to the number of antibiotics for which the isolate was screened for susceptibility. According to Algammal et al., 32 the resistance profiles were classified as MDR, extensive drug resistant (XDR), or pan-drug resistant (PDR).
Molecular detection of resistance and virulence genes
Genomic DNA was extracted using the Maxwell 16-cell DNA purification kit (Promega, Madison, Wisconsin, USA) on an automated machine (Maxwell 16 extraction system, Madison, Wisconsin, USA). The real-time polymerase chain reaction (PCR) assays were performed on the AriaMx system (Agilent Inc., Santa Clara, California, USA). Primers and probes were purchased from LGC, Biosearch (Novato, California, USA) for the different genes based on primers and probes used by Ibrahim et al. 56 for invA; Bugarel Weil et al. 57 for sopB, ssaQ, mgtC, spi4D, spvC, and ljsGI-1; Roschanski et al. 58 for blaTEM, blaSHV; Vien et al. 59 for qnrA; Singh and Mustapha 60 for floR and tetG; and Guarddon et al. 61 for tetA and tetB. Supplemental Tables 1 and 2 show the genes sequences and amplification conditions used. A quality-controlled positive and negative internally characterized known resistant and susceptible Salmonella typhi strains from International Typhoid Consortium 62 were used as controls for amplification for detecting the resistance genes and virulence during PCR. Also, no template controls were incorporated into the PCR as an additional method of internal control in the PCR.
A 12.5 μL of Perfecta master mix low ROX kit (Quanta Bioscience Inc., Madison, Wisconsin, USA), 1.0 μL of each 10 mM primers and probes, 7.5 μL of Nuclease free water (Sigma-Aldrich, St Louis, Missouri, USA), and 2.0 μL of DNA template make up a 25 μL PCR reaction mixture. Thermal conditions were those described by the referenced authors (Tables 1 and 2). After the amplification experiments were completed, the cycle thresholds were determined by identifying the fluorescence signal by analyzing the amplification plots in AriaMx system software version 3.1.
Identification of Salmonella by API 20E.

n: number; %: percentage.
The non-susceptibility pattern between invasive typhoidal and non-typhoidal Salmonellae in the study.

n: number; %: percentage; ICS: intermediate ciprofloxacin susceptibility; IAS: intermediate azithromycin susceptibility. p < 0.05 = statistical significance. p > 0.05 = statistical insignificance.
Chi-square statistic.
Fisher’s exact test.
MIC ciprofloxacin>0.064 µg/mL.
MIC azithromycin >16 µg/mL.
Statistical analysis
Data were imputed and validated in Excel 2016. Descriptive statistics were computed for the multiple antibiotic resistance index. Agreement between the values of antimicrobial resistance phenotypes and their corresponding genotypes was established by κ value (coefficient of agreement) according to Jeamsripong et al. 63 Chi-square and Fisher’s exact test were used to test association as appropriate in every case. p < 0.05 was taken as statistically significant. Statistical Package for Social Science Version 20 (IBM, Santa Barbara, California, USA) was used.
Results
Prevalence and phenotypic characteristics of recovered Salmonella species
Table 1 shows the Salmonella serovars found in the study, S. typhi 51 (61.4%) was the most occurring serovar, Salmonella typhimurium 5 (6.1%), Salmonella enteritidis 6 (7.2%), Salmonella choleraesuis 8 (9.6%), and Salmonella species 13 (15.7%) were the other identified serovars in this study. Figure 1 showed a statistically significant (p = 0.02) higher typhoidal Salmonellae 51 (61.4%) than non-typhoidal 32 (38.6%) Salmonellae in the study.

Classification of Salmonella enterica in the study.
Antimicrobial susceptibility testing, antibiotic resistance genes outcomes
Table 2 shows the resistance pattern of the Salmonella isolates. Of the 83 S. enterica isolates, 65 (78.3%) were resistant to ampicillin and trimethoprim-sulfamethoxazole. A significant (p = 0.03) higher resistance was found in typhoidal Salmonellae 44 (86.3%) compared non-typhoidal Salmonellae 21 (65.6%). Resistance to tetracycline 41 (49.4%) was higher, with a statistically significant difference (p = 0.0004) in typhoidal Salmonellae 33 (64.7%) than in non-typhoidal Salmonellae 8 (25.0%).
Resistance to other antimicrobials was as follows: amoxicillin-clavulanate 21 (25.3%), piperacillin 33 (39.8%), cephalothin 19 (22.9%), chloramphenicol 39 (46.9%), streptomycin 21 (25.3%), azithromycin 9 (10.8%) with no statistically significant differences (p > 0.05) in typhoidal Salmonellae and non-typhoidal Salmonella in the study. Intermediate ciprofloxacin susceptibility occurred in 24 (28.9%) Salmonella isolates. Also, intermediate azithromycin susceptibility occurred in nine (10.8%) Salmonella isolates.
MDR was found in 39 (46.9%) of the 83 S. enterica isolated. Of the 39 S. enterica isolates that demonstrated MDR phenotypically, 30 (58.9%) were typhoidal Salmonellae. In comparison, nine (28.1%) of them were non-typhoidal Salmonellae. No XDR and PDR were observed in the study, as shown in Figure 2. Figure 3 shows the MAR indexes of the Salmonella isolates, 50 (60.2%) of the isolates showed higher statistically significant MAR (>0.2) than the other 33 (39.8%) Salmonella isolates with MAR ⩽ 0.2.

Occurrence of MDR of Salmonella enterica isolated.

Multiple antimicrobial resistance index of Salmonella serovars.
Of the resistance genes investigated, the blaTEM gene with 42 (50.5%) was the most common occurrence in the study. The occurrences of the other resistance genes were as follows, floR 32 (38.6%), tetA 10 (12.0%), tetB 20 (24.1%), tetG 5 (6.0%), and qnrA genes 24 (28.9%). The study showed no blaSHV and blaCTX-M, as shown in Figure 4. Table 3 shows the occurrence of the resistance genes in typhoidal and non-typhoidal S. enterica. There was no statistical significance (p > 0.05) differences in the occurrence of the resistance genes in typhoidal and non-typhoidal S. enterica.

Prevalence of resistance genes from the Salmonella enterica isolates.
Occurrences of resistance genes in typhoidal and non-typhoidal Salmonella in the study.

n: number; %: percentage; NA: not applicable. p < 0.05 = statistical significance; p > 0.05 = statistical insignificance.
Chi-square statistic.
Fisher’s exact test.
For even phenotypic resistance recorded, the corresponding gene was determined, κ agreement analysis was done, and the outcomes showed perfect agreement for chloramphenicol (κ = 0.954), tetracycline (κ = 1), and ciprofloxacin (κ = 1), but there was a moderate agreement between phenotypic detection of β-lactam and the genotypic detection with κ = 0.60 as shown in Supplemental Table 3.
PCR-based detection of virulence-determined genes
Of the seven virulence genes examined, SPIs encoding genes (invA, sopB, mgtC, and spi4D) were found in all the Salmonella isolates 83 (100.0%). Gene spvC occurred in 45 (51.8%) S. enterica recovered in the study. In comparison, gene ssaQ occurrence was found in 33 (39.8 %) of the Salmonella isolates, but the ljsGI-1 gene was found in 2 (2.4%) of the Salmonella isolates shown in Figure 5. The prevalence of spvC genes in typhoidal Salmonellae 30 (58.8%) was insignificantly higher than non-typhoidal Salmonellae 16 (57.1%). In contrast, gene ssaQ occurrence in typhoidal Salmonellae 17 (33.3%) is significantly (p = 0.02) lower than in non-typhoidal Salmonellae 16 (57.1%). The ljsGI-1 gene was found only in two (3.9%) typhoidal sub-group, as shown in Table 4.
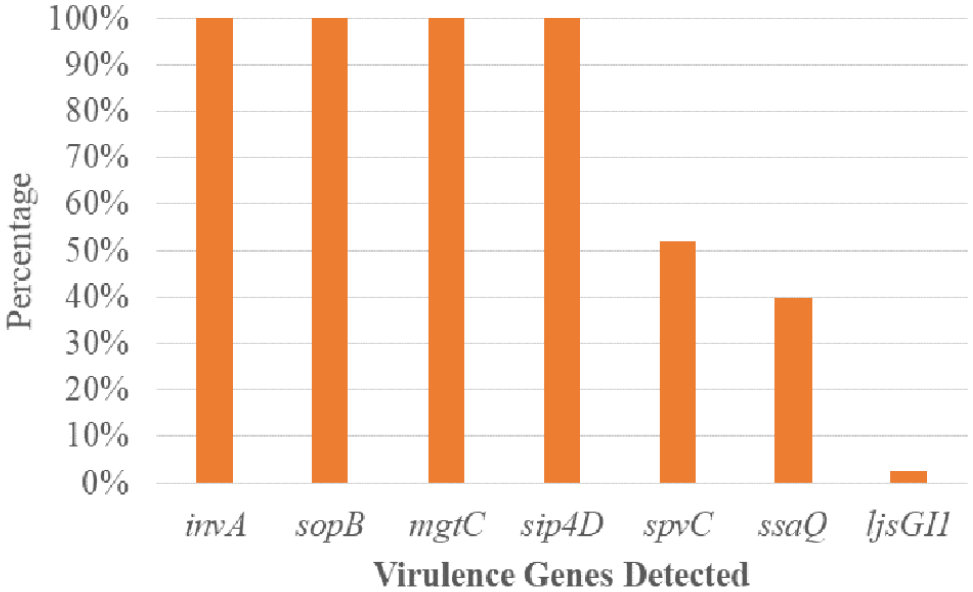
Prevalence of virulence genes from the Salmonella enterica isolates.
Occurrence of virulence genes in invasive typhoidal and non-typhoidal Salmonella.

n: number; %: percentage. p < 0.05 = statistical significance; p > 0.05 = statistical insignificance.
Chi-square statistic.
Fisher’s exact test..
Discussion
Increased resistant S. enterica has continued to pose a significant threat to human health and animal protection, and their spread is being increasingly found in clinical, food, and animal samples. Our study found typhoidal Salmonella serovars and non-typhoidal Salmonella serovars similar to the report by Awol et al. 64 in a multicenter study. Ke et al. 65 and Stanaway et al. 66 show that children in poor and middle-income countries with sub-optimal water, sanitation, and hygiene have a considerable and progressive increase in invasive non-typhoidal Salmonella (iNTS) infection. Invasive typhoid and non-typhoidal Salmonella have been previously reported in Nigerian children.5,8,27,67,68
Regarding the serovars found in this study, S. typhi was the highest, followed by S. choleraesuis, S. enteritidis, and S. typhimurium. Salmonella typhi is the most common serovar in invasive Salmonella infection in children, according to previous studies conducted in Nigeria and elsewhere.5,8,67,69–71 Serovars S. enteritidis and S. typhimurium are not frequently observed in invasive non-typhoidal infections in industrialized countries. Still, in sub-Saharan Africa, they are becoming a reoccurring decimal. Two African authors have previously reported them in invasive Salmonella infection.72,73
High levels of resistance to ampicillin, trimethoprim-sulfamethoxazole, tetracycline, and other routinely used antibiotics were found in our investigation. Salmonella strains isolated from invasive environments resist many of the most widely used antibiotics.5,67,70,71 In particular, multidrug-resistant iNTS caused life-threatening invasive disease outbreaks in children in Nigeria, Rwanda, and Malawi.67,74,75
The antimicrobial resistance of iNTS is a big problem because it can cause bacteremia in immunocompromised people. 76 The high prevalence of antimicrobial-resistant Salmonella is a serious concern for public health. 58 Salmonella typhi was the most common cause of invasive typhoidal Salmonella infection, and its high MDR and low ciprofloxacin susceptibility rates were the most striking findings of our investigation. This result is consistent with patterns seen in Cambodia by Vlieghe et al. 77 when describing MDR in S. typhi. The MDR has been observed in S. enterica in several countries in literature.78–80 Salmonella typhi with multi-drug resistance was found in an assessment of typhoid fever cases in Pakistan, Vietnam, India, China, Indonesia, and Nigeria.73,81–85Our finding regarding MDR in Salmonella isolates in Nigeria agreed with previous reports and the assertion that Nigeria is in the vicinity region referred to as a hotspot for antimicrobial overuse.19,86
The MAR index, a cost-effective and valid method of bacteria origin tracking, was also calculated. It has become possible to tell bacteria apart by their resistance to the most popular antibiotics in human medicine by doing a MAR analysis.87–89 Compared to other methods of bacteria source tracking, such as genotypic characterization, the MAR indexing method is cost-effective, rapid, and easy to perform. 90 MAR index values greater than 0.2 indicate a high-risk source of contamination and index for high antibiotics usage. 88 This study reported a high MAR index >0.2 for S. enterica from invasive sources, which concerns the efficacy of treatment options available. High MAR in S. enterica has been attributed to plasmids containing one or more resistance genes,91–93 each encoding a single antibiotic resistance phenotype. 94 This study did not find XDR and PDR support in literature.91–93
Our results revealed significant positivity of blaTEM in the S. enterica studied. The blaTEM gene found in this study was slightly higher in invasive typhoidal strains than iNTS. The presence of the blaTEM gene in most of the S. enterica in our study supported earlier assertion, which suggested that blaTEM genes code for beta-lactam drug resistance like ampicillin. 95 Beta-lactamase produced by gram-negative bacteria remains the primary mechanism by which they develop resistance to beta-lactam antibiotics. Additionally, ESBLs are increasingly common among S. enterica serovars, and their frequency and prevalence have been reported to rise.96–98 Although blaCTX-M and blaSHV were not found in this study, recent studies from Asia and some parts of Africa have reported them,96,99 specifically reports of the blaSHV gene from clinical S. enterica from India.100–103 Therefore, monitoring the incidence of blaTEM in S. enterica isolates is a crucial public health tool in combatting this threat.
Salmonella enterica isolates with phenotypically intermediate ciprofloxacin susceptibility (ICS) were found to harbor plasmid-mediated quinolone resistance (PMQR) genes (qnrA). This observation agrees with a report from India 104 with observations in human samples (51.4%), food-producing animals (28.6%), environmental samples (11.4%), and animal samples (8.6%), respectively.89,105–107 Ciprofloxacin has become prominent in treating severe infections caused by S. enterica, especially those resistant to nalidixic acid, which has increased significantly in recent years. Still, high resistance levels to ciprofloxacin are rare, but its resistance is foundational for other resistance mechanisms.75,108,109 Mutants resistant to fluoroquinolones are being rapidly selected due to the spread of PMQR genes. 110 Moreover, interactions between mutations in the QRDR and PMQR genes might result in high fluoroquinolones MIC. However, a study 111 speculated that the qnr genes could increase fluoroquinolone resistance.
In literature, tetA, tetB, and tetG were consistently found in S. enterica of human origin. The tetB gene is predominant among the phenotypic tetracycline-resistant strains in the literature. Our findings agreed with two studies that have reported tetA and tetB in S. enterica from a human with gastroenteritis in India and Nigeria.112,113 tetG has also been reported in humans with bacteremia in Nigeria. 113 However, studies examining tetracycline resistance in multiple isolates reported tetA, tetB, and other types in S. enterica isolates from humans and those from animals, environments, and poultries.113,114 Regarding the family Enterobacteriaceae, the tetB and tetA tetracycline resistance determinants have historically been the most prevalent. 112 However, tet (C, D, E, M, O) associated with tetracycline resistance in Salmonella species and other bacteria are less frequently found.
Salmonella isolates that have developed phenotypic resistance to chloramphenicol are strongly linked to the development and expression of efflux that pumps the drug out of the bacteria’s cells,115,116 encoded by floR or cml genes. In our study, all invasive S. enterica phenotypically resistant to chloramphenicol had floR gene. These findings agree with reports of floR gene detection from S. enterica in the literature.117,118 Also, it has been asserted that the floR gene of Salmonella pathogenicity island-1 contributes to S. enterica infectivity. 92 Chloramphenicol was one of Nigeria’s most common drugs of choice in treating Salmonella-related infections. A survey revealed 72.4%–89.2% increased resistance from 1997 to 2007, thus limiting its therapeutic value.117,118 In Iraq, chloramphenicol-associated genes were highly occurring in S. enterica strains isolated from clinical samples.119,120 In literature, chloramphenicol was the first-line drug used to treat typhoid fever, but its recurrent use limits its therapeutic value due to resistance development.119,120
The presence of invA, sopB, spi4D, and mgtC genes in all the tested isolates agreed with the literature’s earlier evidence.13,121,122 Salmonella invasion gene (invA) is involved in the invasion of the intestinal epithelium cells and is found in pathogenic S. enterica. 123 Therefore, for Salmonella infection to occur, invasion of the cells must occur, aided by invA gene.124–126 The invA gene influences the type of Salmonella infection that could result in either systemic or localized. 127 This gene is a transcriptional regulator required to express several genes encoding type III secretion system SPI-1 effector proteins.57,121,128 The invA gene was previously hypothesized to be widely distributed among the S. enterica isolates irrespective of their serovars or source of isolation. Thus, the invA gene is a suitable target for detecting S. enterica from different biological specimens, as documented in the literature.56,60,121,127,129–131
The inositide phosphate phosphatase (sopB) gene is an effector protein that induces macropinocytosis. Gene sopB is an actin-binding protein that interacts with the host cell actin cytoskeleton. It is required for efficient bacterial internalization by the host cell. 132 The sopB gene in all isolates is instructive because sopB has been reportedly involved in micropinocytosis. 133 Macrophages have been considered the main target of Salmonella during infection, and these cells are responsible for bacterial dissemination and control.134–136 In addition to macrophages, other immune system cells are targets of S. enterica pathogens, dendritic cells, and neutrophils.
Furthermore, B cells have also been targeted by S. enterica through the expression of sopB. 137 The sopB genes are necessary for intracellular survival in the host, so the presence of sopB gene is suggested to contribute to the invasiveness of S. enterica pathogens,138,139 found in all the isolates in this study. The sopB gene is also involved in host cell survival by activating the Akt signaling pathway, including activation of the host innate immune system and cell death. 133 The presence of bacterial effector sopB in our study supported the earlier assumption that activation of Akt pathway is mediated through the expression of sopB.
In literature, 126 Salmonella’s SPI-3 island is associated with intra-macrophage invasion, which supports survival when Mg2+, required in the bacteria transported system, is of limited amount. The presence of mgtC gene in all the Salmonella in this study supported the proposition that S. enterica uses the expression of mgtC gene to circumvent the lack of Mg2+ in the bacteria. They, therefore, initiate Mg2+ production without depending on the host for Mg2+. Our findings are supported in earlier literature.13,137,139,140 Salmonella enterica contains several transport systems, both inducible and constitutive. 140 These transport systems have functional complementarities to adjust the Mg2+ concentration in different environmental conditions. In addition, these systems are controlled by transcriptional and post-transcriptional regulatory networks to maintain strict control of the Mg2+ balance. 126 Regarding maintaining Salmonella viability and development in environments with low Mg2+ levels, mgtC appears to be the most crucial SPI-3 component, as reported in some S. enterica isolates 140 and S. typhi. 13 Since mgtC is encoded in a region of SPI-3 that is highly conserved, it plays a crucial role in virulence that is not met by any other factor encoded in SPI-3 or anywhere else on the S. enterica chromosome.121,141
The ssaQ gene was detected in 33 (39.8 %) Salmonella isolates examined. The importance of this gene is relevant in the surveillance of S. enterica, which has been involved in systemic infection in the past. It has been found to produce proteins for the bacteria that bind to and stabilize the larger protein, which is important for the overall efficiency of the secretory system. 142 Essential for virulence in host cells, survival in macrophages, and biofilm development is the ssaQ gene, which codes for proteins in the SPI-2 type III secretion system. 143
The Salmonella plasmid virulence (spvC) gene was significantly higher in iNTS than in typhoid Salmonella isolates. By eliminating their beta-subunits, the spvC gene renders inactive the host’s dual-phosphorylated mitogen-activated protein kinases. It is also hypothesized to play a role in systemic S. enterica infection due to its anti-inflammatory effector effects and attenuation of the intestinal inflammatory response. 144 The spvB gene may collaborate with spvC and other Salmonella effectors to play a role in pathogenesis by triggering apoptosis in human macrophages. The spv genes increase the virulence of non-typhoid Salmonella serovars to induce extra-intestinal illness, as shown by experimental models and human epidemiological data. 145 Intestinal infections caused by non-typhoid Salmonella, typically present as self-limiting gastroenteritis, can be terminated by spv genes. 129 In mice, a study 145 discovered that the spv locus in Salmonella serovars is a crucial distinction in the pathogenesis of typhoid fever compared to that of non-typhoid Salmonella bacteremia.
Study limitations
As this is a further study on Salmonella isolates from an initial isolation process, the sample size was not determined; as such, all the Salmonella isolates recovered from 2015 to 2018 were included in this study. Gene sequencing of the antibiotic resistance and the virulence genes of Salmonella isolates detected were not done to detect mutations that could adversely affect the activity of the antimicrobial agents and their pathogenicity abilities.
Conclusions
The result of our study illustrates the emergence of multi-drug resistant S. enterica from children with bacteremia in north-central Nigeria. The most common antibiotics that S. enterica recovered were resistant to were ampicillin and trimethoprim-sulfamethoxazole. Some recovered S. enterica demonstrated multi-drug resistance to penicillins, first-generation cephalosporin (cephalothin), phenicol, sulfonamide, tetracycline, aminoglycosides, and fluoroquinolone. None of the S. enterica isolates met the criteria required for XDR and PDR designation. The recovered antimicrobial resistance genes (blaTEM, qnrA, floR, tetA, tetB, and tetG) were found. The most prevalent gene was blaTEM, while tetG was the least prevalent resistance gene. The invA, sopB, mgtC, and sip4D were found in all the recovered S. enterica isolates. At the same time, most S. enterica also harbored spvC and ssaQ genes, respectively, with ljsGI-1 gene found in only two S. typhi isolates. Therefore, this study recommends continuous monitoring of antimicrobial resistance patterns of S. enterica from invasive sources in Nigeria and encourages the prudent use of antibiotics and the practices of other infection prevention control measures.
Supplemental Material
sj-docx-1-smo-10.1177_20503121231175322 – Supplemental material for Antimicrobial resistance and virulence genes of invasive Salmonella enterica from children with bacteremia in north-central Nigeria
Supplemental material, sj-docx-1-smo-10.1177_20503121231175322 for Antimicrobial resistance and virulence genes of invasive Salmonella enterica from children with bacteremia in north-central Nigeria by Leonard I Uzairue, Olufunke B Shittu, Olufemi E Ojo, Tolulope M Obuotor, Grace Olanipekun, Theresa Ajose, Ronke Arogbonlo, Nubwa Medugu, Bernard Ebruke and Stephen K Obaro in SAGE Open Medicine
Footnotes
Acknowledgements
We would like to thank the Laboratory staff of the International Foundation Against Infectious Disease in Nigeria for their assistance during the project implementation.
Author contributions
Leonard Uzairue was involved in conceptualization, investigation, methodology, formal analysis, visualization, and writing; Olufunke Shittu was involved in conceptualization, editing, and supervision; Olufemi Ojo and Tolulope M. Obuotor were involved in editing and supervision; Grace Olanipekun, Theresa Ajose, and Ronke Arogbonlo were involved in project administration and supervision. Nubwa Medugu and Bernard Ebruke were involved in methodology, supervision, and editing; and Stephen Obaro was involved in conceptualization, supervision, and editing.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Ethical approval and consent to participate
The research got approvals from the Research Ethics Committees of the Federal Medical Centre Keffi (FMC/KF/HREC/052/15), the Nyanya General Hospital, FCT, Abuja (FCTA/HHSS/HMB/NH/GEN/54/II/128), and the University of Abuja Teaching Hospital (FCT/UATH/HREC/PR/61). Written informed consent was obtained from the parent or guardian of the children. The identities of all data/samples used for this study were removed entirely.
Informed consent
Written informed consent was obtained from the parent or guardian of the children.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.