
Abstract
Objective:
Mast cells are associated in angiogenesis in various human and animal neoplasms. However, association of mast cells with tumor microvessel density in canine lymphoma was not previously documented. The objective of the study is to determine if mast cells are increased in canine nodal lymphomas and to evaluate their correlation with tumor microvessel density and grading of lymphomas.
Methods:
Nodal lymphomas from 33 dogs were studied and compared with nonneoplastic lymph nodes from 6 dogs as control. Mast cell count was made on Toluidine blue stained sections. Immunohistochemistry using antibody against Factor VIII was employed to visualize and determine microvessel density.
Results:
The mast cell count in lymphoma (2.95 ± 2.4) was significantly higher (p < 0.05) than that in the control (0.83 ± 0.3) and was positively correlated with tumor microvessel density (r = 0.44, p = 0.009). Significant difference was not observed in mast cell count and tumor microvessel density among different gradings of lymphomas.
Conclusions:
Mast cells are associated with tumor microvessel density in canine nodal lymphoma with no significant difference among gradings of lymphomas. Mast cells may play an important role in development of canine nodal lymphomas. Further detailed investigation on the role of mast cells as important part of tumor microenvironment in canine nodal lymphomas is recommended.
Introduction
Angiogenesis is an essential process in tumor growth 1 and facilitates tumor development and metastasis. Several mast cell (MC)-associated mediators are known to be angiogenic and regulate endothelial cell proliferation. 2 MCs are reported to play an important role in tumor development and angiogenesis in various neoplasms in humans. Their role in promoting neoplasm angiogenesis and development in human basal cell carcinoma, 3 lung adenocarcinoma, 4 squamous cell carcinoma, 5 plasmacytoma, 6 and lymphomas7,8 has been reported. In contrast to the well-recognized angiogenesis in various types of human malignancies, the role of angiogenesis in neoplasm development is hardly recorded in the veterinary literature. Few reports are available indicating a positive correlation between mast cell count (MCC) and microvessel density (MVD) (angiogenesis) in canine neoplasms. For example, MC-associated angiogenesis that promoted tumor growth and development was reported in canine melanoma, 9 transmissible venereal tumor, 10 hemangioma, hemangiosarcoma (HSA), 11 and mammary carcinoma. 12 Our previous study 13 also reported similar findings in canine cutaneous hemangioma, cutaneous HSA, mammary adenoma, and mammary adenocarcinoma with higher MC numbers recorded in malignant tumors than their benign counterparts. Association of MCs with the angiogenesis in canine nodal lymphomas is not previously documented. The aim of this study was to determine if MCs are increased in canine nodal lymphomas in correlation with MVD. This study, for the first time, documented the association of MCs with MVD in canine nodal lymphoma.
Materials and methods
Tissues
Surgical biopsy specimens from 33 dogs with nodal lymphoma (n = 33), received at the Tifton Veterinary Diagnostic and Investigational Laboratory, College of Veterinary Medicine, The University of Georgia, USA, were used in the study. The nodal lymphomas were graded according to Working Formulation 14 and Kiel 15 classification designed mainly for nodal lymphomas. 16 Canine lymph nodes collected at necropsy in Tifton Veterinary Diagnostic and Investigational Laboratory and having no evidence of inflammation or neoplasia on gross and microscopic examinations were included as control (n = 6). The age of the dogs with lymphoma varied from 2 to 14 years (x- = 8.2 years), while the age of the control dogs varied from 5 to 14 years (x- = 9.0 years). The specimens were fixed in 10% buffered formalin, processed for routine histology, replicate sections cut at 5 µm, stained with hematoxylin-eosin (H&E) and Toluidine blue stains, and examined by light microscopy.
MCC
MCC was made on Toluidine blue (Sigma, St Louis, MO, USA) stained sections. The MCs were independently counted and recounted by two investigators including a board certified pathologist. MCs within the tumor mass and on the periphery in nodal lymphomas and in the control were counted in six high power fields (400×) inside a square reticulum (0.64 mm2) inserted in the ocular piece of a light microscope. 13
MVD
Immunohistochemical staining procedures for Factor VIII (Rabbit Polyclonal; Dakocytomation, Carpinteria, CA, USA) were applied to visualize and identify the blood vessels. The standard streptavidin–peroxidase conjugated method was used as follows. Tissue samples were cut at 4–5 µm sections, deparaffinized by xylene–ethanol sequence, and rehydrated in graded ethanol solutions. Antigen retrieval was made by Heat Induced Epitope Retrieval (HIER) using a 1:10 diluted Target Retrieval Solution, pH 6 (DAKO). Heat was supplied by a Black and Decker Vegetable steamer for 25 min, followed by 10 min in the hot solution on the counter top. Endogenous peroxidase was blocked using 11% hydrogen peroxide in methanol for 20 min. Each step of incubation was followed by a thorough washing of the sections with Tris Buffered Saline (TBS) at pH 7.4. The sections were incubated for 2 h at room temperature with primary antibody against Factor VIII at 1:150 dilutions. The sections were then treated with secondary biotinylated anti-rabbit immunoglobulins in TBS, containing stabilizing protein and 0.015 mol/L sodium azide, for 20 min at room temperature. The sections were washed with TBS, and incubated in streptavidin conjugated horseradish peroxidase in phosphate buffer saline containing stabilizing protein and antimicrobial agents for 20 min at room temperature. Finally, all sections were treated with 3,3′-diaminebenzidine (DAB, DAKO) and counterstained with Gill’s Hematoxylin (Gills III-Formula, Surgipath Richmond, IL, USA). Negative control sections were made by substituting TBS for the primary antibody.
The stained sections were scanned at 100× magnification, and areas of high MVD (hot spots) were identified. The microvessels at hot spots were counted following a previously described method. 17 Briefly, six 400× fields of the sections in each sample were examined within a square reticulum (0.64 mm2) inserted in the ocular piece. Transversally sectioned microvessels (capillaries and small venules), with a single layer of endothelial cells, either with or without a thin basement membrane were selected from all stained vessels. Multilayered vessels or those with muscular wall were excluded. Only those occupying the reticulum points were counted. As the microvessel diameter is smaller than the distance between adjacent points, only one transversally sectioned microvessel could occupy a given point. Microvessels transversally sectioned outside the points and those longitudinally or tangentially sectioned were omitted to ascertain that a given microvessel is counted only once, even in the presence of several of its section planes.
Statistics
The MCC and MVD were calculated and the results were given as mean ± standard deviation (SD). Analysis of variance (ANOVA) was used to analyze the data among the three gradings of lymphomas, and Student’s t-test was employed to compare two groups. Correlation between MCC and MVD was determined using Spearman’s correlation coefficient. In all cases, the results were considered statistically significant if p values were <0.05.
Results
Increased MCC and MVD were observed in the lymphomas than in the control (Figure 1). The MCC in lymphoma (2.95 ± 2.4) was significantly higher (p < 0.05) than that in the control (0.83 ± 0.3). The MCs and microvessels in lymphomas were accentuated on the periphery (invasive edge) of the neoplasm (Figure 1(D)). Scattered MCs were observed in the tumor mass, around blood vessels, and the tumor-supporting stroma. The MCC on the periphery (x- = 2.9) was significantly (p < 0.0001) higher than the MCC within the neoplastic mass (x- = 0.3). In one case of lymphoma, although high MVD (6.67 ± 1.5) was recorded, MCs were not present. The MCs observed in the control were detected in the medullary spaces, and supporting stroma, and were not observed within the lymphoid follicles (Figure 1(A)).

Dog, sections of lymph nodes. (A) Lymph node from a control dog: lymphoid follicle; asterisk (*) shows follicular center; Toluidine blue stain, bar = 50 µm. (B) Nodal lymphoma: several mast cells (arrows) on the periphery of the neoplasm and around blood vessels; Toluidine blue stain, bar = 50 µm. (C) Canine lymph node from a control dog: double arrows show the capsule of the lymph node (note the absence of microvessel on the periphery). Immunohistochemical stain for Factor VIII, bar = 100 µm. (D) Nodal lymphoma: immunohistochemical stain for Factor VIII shows increased vascularization on the invasive edge (periphery) of the neoplasm (arrows); bar = 100 µm.
Overall, there is a significant increase in the number of MCs (2.95 ± 2.4 versus 0.83 ± 0.3; p = 0.0419) and MVD (2.4 ± 1.5 versus 0.36 ± 0.24; p = 0.0023) in the lymphoma than in the control. Statistically significant positive correlation (r = 0.44, p = 0.009) between the MVD (2.45 ± 1.5) and MCC (2.95 ± 2.4) was observed in the lymphoma, but not (r = 0.33, p = 0.519) between the MCC (0.83 ± 0.3) and MVD (0.36 ± 0.2) in the control (Tables 1 and 2).
MCC and MVD (mean ± standard deviation) in canine lymph nodes (control).
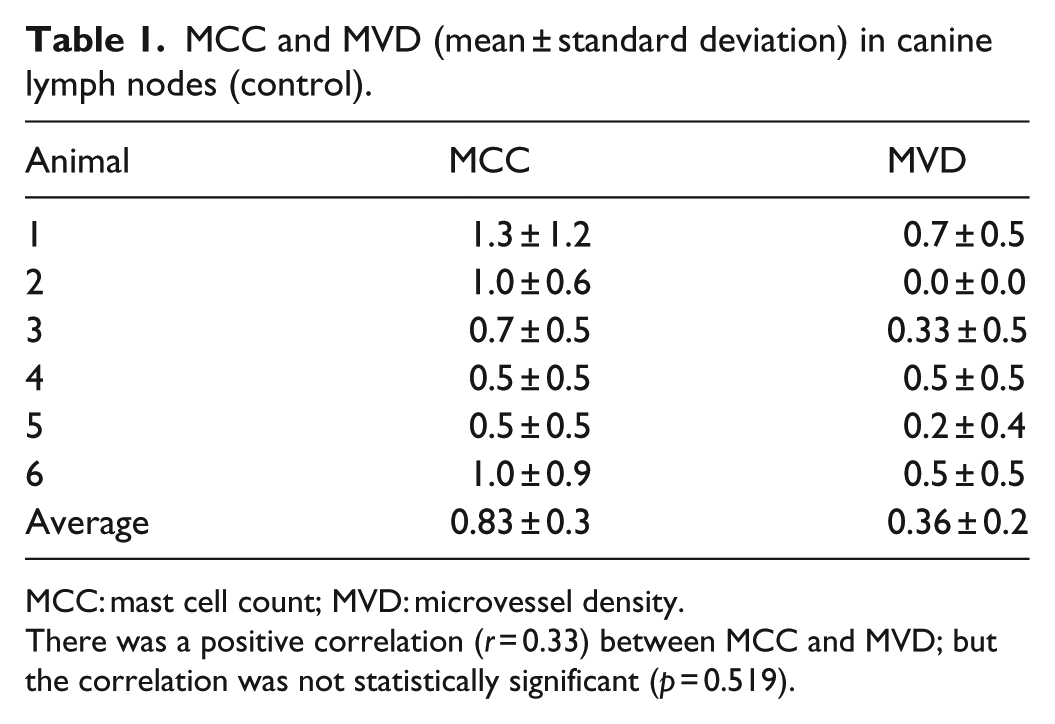
MCC: mast cell count; MVD: microvessel density.
There was a positive correlation (r = 0.33) between MCC and MVD; but the correlation was not statistically significant (p = 0.519).
MCC and MVD (mean ± standard deviation) in canine nodal lymphoma.

MCC: mast cell count; MVD: microvessel density.
H = high-grade; L = low-grade; IM = intermediate-grade.
There is a significant positive correlation between the MCC and MVD (r = 0.44216; p = 0.009).
The lymphomas were classified as high-grade, (13/33, 39.4%), low-grade (6/33, 18.2%) and intermediate-grade (14/33, 42.4%) based on Working Formula classification and high-grade (24/33, 72.73%) and low-grade (9/33, 27.27%) based on Kiel classification. There was no statistically significant difference (p > 0.05) between MCC and MVD among the grades of lymphomas based on Working formulation. The MVD and MCC were positively correlated (low-grade: r = 0.24, p = 0.65; intermediate-grade: r = 0.67, p = 0.008; high-grade: r = 0.17, p = 0.57) in the various gradings of lymphomas although the correlation was statistically significant (p < 0.005) only in intermediate-grade lymphoma. Similarly, there was no statistically significant difference (p > 0.05) between MCC (2.97 versus 2.93) and MVD (2.74 versus 1.98) between the low-grade, and high-grade lymphomas, respectively, based on Kiel classification. The MVD and MCC were positively correlated in both high-grade (r = 0.464, p = 0.022) and low-grade (r = 0.354, p = 0.348) lymphomas with significant correlation (p < 0.05) in high-grade lymphomas, but not in low-grade lymphomas.
Discussion
A significantly higher (p < 0.05) MCC was documented in canine nodal lymphomas than in the control. The MCC in the lymphomas was positively correlated (r = 0.44, p = 0.009) with MVD. The finding in this study is in concurrence with our previous study that reported association of MCs with MVD in canine cutaneous hemangioma, cutaneous HSA, and mammary tumors. 13 Other investigators also reported positive correlation of MCC with MVD in canine melanoma, 9 transmissible venereal tumor, 10 hemangioma, HSA, 11 and mammary tumor. 12 Increased MCC and MVD were associated with tumor development, metastasis, 12 and poor prognosis.9,10 There was no statistically significant difference (p > 0.05) between MCC and MVD among the gradings of lymphomas based on Working formulation 14 as well as Kiel 15 classification system. Two previous reports18,19 on MVD in canine lymphomas reported variable results on MVD in high-grade and low-grade lymphomas based on similar classification systems,14,15 although both studies did not consider MCs and their association of MVD in the studies. No difference was recorded in MVD among gradings of lymphomas based on both classification systems in one study, 18 while a higher MVD in high-grade lymphomas than in low-grade lymphomas was reported in one other study. 19 In humans, similarly, one study 20 reported no correlation between tumor MVD and gradings of malignancy in nodal lymphomas, while another report 21 documented significant difference in MVD among subtypes of lymphomas. The discrepancies are ascribed to different classification systems used, heterogeneity of the patients in different studies, variation in counting and staining methods, and antibodies used to highlight the microvessels. 18 For example, several authors employed Toluidine blue stain for identification of MCs, while others used immunohistochemistry with a primary anti-tryptase antibody.
In this study, a positive correlation was observed between MCC and MVD in all gradings of lymphomas based on both classification systems. However, the correlation was statistically significant in intermediate-grade (Working formulation) and high-grade (Kiel classification) lymphomas. The findings plausibly agree as could be seen from the fact that 78.57% (11/14) intermediate-grade lymphomas classified in the Working formulation belong to the high-grade lymphomas in the Kiel classification system. MC-associated angiogenesis is also reported in human Hodgkin-lymphoma, 7 non-Hodgkin lymphoma,22,23 and cutaneous lymphomas 8 in concurrence with the current study. The authors indicated MCs to be sources of angiogenic factors that promoted angiogenesis.22,23 Furthermore, the significance of MCs in cutaneous lymphoma was demonstrated by an in vitro experiment of a mouse model of human cutaneous lymphoma, in which tumor growth in MC-deficient transgenic mice was significantly decreased. 8 The authors further showed that MCs play a protumorigenic role in the cutaneous lymphomas.
Generally, MCs are involved in tumor angiogenesis due to the release of their various mediators and pro-angiogenic factors2,12 that regulate endothelial cell proliferation. 2 In humans, it is reported that levels of angiogenic factors in tissue reflect the degree of aggressiveness with which tumor cells spread, and thus have predictive value in the identification of the high-risk patients. 24 MCs modify tumor microenvironment; promote marked neovascularization, tumor growth as well as fibrosis; and correlate directly with disease severity in human Hodgkin-lymphoma. 7 Similarly, our previous study 13 recorded higher MCC in canine HSA than in hemangioma and in canine mammary adenocarcinoma than in adenoma indicating an increase in the MCC with malignancy. In concurrence with previous reports on cutaneous lymphomas in humans, 8 and other canine neoplasms, 13 MCs in lymphomas in the current study were accentuated on the invasive edge of the neoplasm associated with blood vessels suggesting aggressive and invasive nature of the tumor.
The distribution pattern of the MCs observed within medullary spaces and perivascular stroma in the control is similar to normal tissue-resident MCs 13 located in the perivascular stroma and close to neurons 25 in various tissues and organs.
MCs were not observed in one case of lymphoma with high MVD (6.67 ± 1.5) on the periphery. In concurrence with this, Sabattini and Bettini 11 also reported infiltrating MCs in the stromal compartment of all canine HSAs except one splenic HSA in which no MCs were detected. The reasoning behind why MCs were absent in rare neoplastic cases, while commonly seen in similar other neoplasms, is unknown. Whether the variation is idiopathic or associated with the stage of the neoplasm development or other mechanisms which underlie the absence of MCs in rare cases of some neoplasms needs further investigation.
It is also worth noting that MCs are highly heterogeneous in terms of their morphology, function, and metabolic products. 26 Despite a well-documented association of MCs with tumor angiogenesis and development, variable roles of MCs are reported in different types of tumors in humans. Reports indicate that MC accumulation can either be beneficial or detrimental for tumor growth,27,28 depending on the local stromal conditions 28 and the stage of tumorigenesis. For example, MCs exhibited protumorigenic capacity and promoted angiogenesis in late stage malignancy of intestinal tumorigenesis in humans, and played a protective role by promoting apoptosis of tumor cells in early stage. 29 MCs may play both angiogenic and antiangiogenic roles in different stages of hemangioma. 30 For example, a highest number of MCs was recorded during the involuting phase, less in the involuted phase, and least in the proliferative phase of hemangiomas in humans. 26 Furthermore, angiogenesis is regulated by both activator and inhibitor molecules. More than a dozen different proteins have been identified as angiogenic activators and inhibitors. 24 Inconsistent results between MC density and tumor development in different tumors may be due to differences in effect of MC chemotactic factors released from cancer cells on stromal MC reactions. 5
The fact that tumor growth of human cutaneous lymphoma model was significantly decreased in MC-deficient mice 8 and that MCs are associated with invasiveness, severity, and malignancy of various neoplasms4–7,13,17,27,31 suggests that controlling proliferation of MCs may play an important role in minimizing tumor growth and invasiveness.
Conclusion
Significantly increased MCC in canine nodal lymphomas and its correlation with MVD in this study suggests that MCs may play an important role in development and invasiveness of canine nodal lymphomas. We recommend further detailed investigation on the role of MC as important part of tumor microenvironment in canine nodal lymphomas.
Footnotes
Declaration of conflicting interests
The authors declare that there are no conflicts of interest regarding the publication of this article.
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.