
Abstract
Bacterial infections are a major public health threat, with a substantial global burden of ∼5 million deaths in 2019, of which ∼1.27 million were attributed to antibiotic resistance. The formation of bacterial biofilms has significantly enhanced bacterial resistance to antibiotics. Worse still, it increases overall bacterial pathogenesis, contributing to inflammation and potentially to carcinogenesis in humans. Biofilm is implicated in approximately 65% of all bacterial infections and 78.2% chronic wound infections. Alarmingly, about 100–1000-fold increase in antibiotic concentration is required to eradicate bacteria within biofilms, further compromising the health of already ill-patients. Therefore, it is imperative to explore potential antibiofilm agents, especially ones with novel mechanisms of action, to clinically manage inpatient biofilms. Bacteriophage (phage) use is a promising evolutionary approach but is also challenged with potential resistance. Bacteria have developed several antiphage defense mechanisms, some of which exhibit synergistic antiphage activity. In this review, we provide several lines of evidence supporting the efficacy of phages against antibiotic-resistant clinical biofilm-forming bacteria. Observations reveal that phage enzymes disrupt biofilm structural components (e.g., EPS, pectate, and hyaluronic acid) and pave the way for phage infection of naked bacterial cells. We further provide insights into the recent advancements in phage use against biofilm-associated antibiotic-resistant bacteria in patients. Current knowledge shows that phages are rapidly evolving and counteracting antiphage bacterial mechanisms. Here, future perspectives to enhance phages efficacy against biofilm resistance are provided to establish their clinical antibiofilm application. Enhancing the clinical application of phages against biofilms requires addressing bacterial host biofilm resistance and optimizing strategies accordingly. Beyond phage cocktail and phage genetic engineering, conjugating phages with antimicrobial agents (eg., antimicrobial peptides) offers a compelling strategy to enhance phage antibiofilm efficacy.
Introduction
In previous centuries, sulfonamides, arsenicals, heavy metals, natural products, and drugs made by chemical tinkering with synthetic dyescrude were utilized in the treatment of a variety of bacterial infections, but these had significant side effects, limiting their systemic antibacterial application. 1 The discovery of penicillin by Fleming in 1928 and the subsequent development of various classes of antibiotics enhanced the golden age of the antibiotic era, where innumerable lives were saved from the devastating effects of bacterial infections. However, the widespread excessive dispensing, the irresponsible use of antibiotics, and the multifaceted bacterial capacity for “adversity-induced” evolution have resulted in the development of antibiotic-resistant bacterial strains.2–4 This has impacted the potency of antibiotics, and is rapidly driving bacterial infectious disease management into the “post-antibiotic era,” with many bacterial infections now requiring last-resort antibiotics in their treatment.5,6 Nevertheless, the introduction of lipopeptides in 1987 marked the last addition of a new class of antibiotics to the market, 7 leaving a gap of over three decades in antibiotic innovation. 3 The problem is further exacerbated by the continuous rise in infections caused by multidrug-resistant (MDR), extensively drug-resistant (XDR), and pan-drug-resistant (PDR) bacteria.8–11 The emergence and escalation of these drug resistance traits pose a major threat to public health on a global scale. A recent analysis indicates that antimicrobial resistant bacteria accounted for a substantial global burden in 2019. It was associated with an estimated ∼5 million deaths, of which ∼1.27 million were attributed to their resistance. 12 This menace is more concentrated in developing countries and may be higher than estimated due to the inadequate surveillance structures in these regions.12–16
The formation of bacterial biofilms increases overall bacterial pathogenesis, contributing to inflammation and potentially to cancer formation (carcinogenesis) in human systems. 17 Additionally, it significantly enhances bacterial resistance to antibiotics among patients, thereby facilitating bacterial dissemination to other potential sites for infection. This observation is supported by recent statistics that infections associated with biofilms account for approximately 65% of all bacterial infections and up to 78.2% of chronic wound infections.18–21 Worryingly, eradicating bacteria within biofilms requires a substantial increase in antibiotic concentration, often 100–1000 times higher than the usual dosage.22,23 This extremely high antibiotic concentration is not only toxic (e.g., nephrotoxicity) to consumers, but also erodes the protective physiological microbiota (dysbiosis). 24 Consequently, it is imperative to explore potential clinically applicable antibiofilm agents, especially ones with novel mechanisms of action 25 to overcome inpatient biofilms. The use of bacteriophages, also known as phages, has emerged as a plausible evolutionary approach.
Phages are natural predators of bacteria that target specific strains of bacteria and are the most abundant entity on earth, thereby making it possible to isolate phages specific to numerous clinically relevant bacterial strains. Phages can be grouped into two categories based on their life cycle: the lysogenic/temperate phage, which establishes a coexisting state with the bacteria by integrating its genetic material within the host bacterial genome, and the lytic phage (the type of phage under consideration in this review), which injects its genetic material into the host bacteria and utilizes the host ribosomes to synthesize its own proteins, thereby inducing bactericidal activity (Figure 1).19,26,27 Nonetheless, just like antibiotic resistance, phages are also challenged by bacterial resistance due to the multifaceted antibiotic resistance mechanisms of bacteria, especially biofilm-forming ones. This review, therefore, sought to provide insights into the basic science and recent advances of phage therapy for targeting antibiotic-resistant biofilm-forming bacteria. It further provides future perspectives on enhancing phage efficacy over biofilm-associated resistance to support its clinical application in antibiofilm therapy. Even though lytic phages are often combined with antibiotics in the treatment of biofilm-forming infections,28,29 this review focuses exclusively on the use of lytic phages.

The life cycle of lytic and temperate phages.
Phage interaction with bacterial biofilms
Bacterial biofilms are sessile communities of bacterial cells encased in a self-produced extracellular matrices. 30 They commonly adhere to biotic or abiotic surfaces, including lung tissues, medical implants, air–liquid interfaces, and inanimate surfaces as single-cell biofilms or mixed-species biofilms.31,32 The formation of biofilms involves a complex, yet finely regulated series of processes that commence with quorum sensing, and can be categorized into a few steps: (1) sensing, (2) initial contact with surface (reversible and irreversible attachments), (3) microcolony formation, (4) maturation and architecture, and (5) dispersion of biofilms (Figure 2).18,19,23,33 Biofilms of different bacterial strains may be made up of different matrices, but are generally comprised of core components such as a polysaccharide matrix, lipids (lipoprotein), fibrin, exopolysaccharides, proteins, nucleic acids; RNA, extracellular DNA (eDNA), ions like Ca2+, and other biomolecules to generate a hydrated, gelatinous, three-dimensional architecture biomaterial referred to as “extracellular polymeric substances” (EPSs), which is the core of biofilms.19,23,34–40 These mixed components provide mechanical stability, protect adhering bacteria from environmental assaults, enhance the default mode of growth, and impede the penetration of antibiotics by conferring rigidity and viscosity. The extracellular polymer matrix of the biofilm not only acts as a diffusion barrier but also possesses the capability of directly binding to antibacterial agents, effectively preventing their entry into bacterial cells.23,41–43 These culminate in the promotion of highly persistent bacterial infections, 44 due to the pathogens’ ability to withstand antimicrobials and evade the immune system.42,45 While biofilms prevent the entry of stressors and antibacterials, cells within the biofilm perform both active and passive diffusion for their movement and any associated molecules transfer, respectively. These movements prevent the bacteria from being trapped in a confined zone. 23

The biofilm formation cycle and bacteriophage invasion at each stage.
As with many other antibacterial agents, phages, which are highly specific in their actions toward bacterial species, first penetrate the biofilm structures that surround the bacterial colonies via intricate interactions with the components of the biofilm. Despite this prerequisite to therapeutic application on bacterial biofilms, phages are nonmotile and rely on Brownian motion, which is a random movement resulting from collisions with surrounding molecules, to navigate their environment and reach their targets.46,47 This, therefore, presents the perplexing question of whether phages can depend on Brownian motion to successfully traverse the biofilm and reach the target bacterial cells. This is, however, not entirely the case—the interaction of phages with these biofilm components involves not only the phage as a whole but proteins and enzymes produced by the phages. These enzymes and protein molecules enhance the favorable antibiofilm interaction between phages with biofilms. In this context, depolymerase enzymes, produced by some phages 48 degrade the polysaccharide layer of biofilms, resulting in potential damage to the biofilm structure by targeting the stages of microcolony formation and biofilm maturation. 48 In the case of naïve bacteria or bacteria with biofilm in the early development stages, the bacterial cells face internal lysis by lysine and putative T4-like lysozyme, effectively inhibiting their proliferation and preventing them from reaching the critical density required for mature biofilm generation. 48 Matured biofilms, on the other hand, can also be raptured with the aid of depolymerase enzymes of phages, degrading the bacterial polysaccharides, 23 alongside dispersing the chemical components of the biofilm matrix. A recent study by Park et al. identified a novel depolymerase, Dpo10, which exhibits the ability to specifically bind to and degrade the lipopolysaccharide of Escherichia coli O157: H7. Dpo10 demonstrated convincing environmental stability with high activity in preventing the formation of biofilms on various abiotic surfaces, as lipopolysaccharide is an integral component of biofilm formation. 49
Regardless of whether phages kill the bacterial cell, they penetrate and damage the biofilm during the biofilm control process.48,50 In addition to depolymerase and lysozymes on biofilm during the early and mature stages of biofilms, phage endolysins also digest the extracellular polymeric substance (matrix) of biofilms 51 and also weaken the bacterial cell wall by degrading the peptidoglycan, allowing for lysis and bacterial cell death (Figure 2). 52 The action of this enzyme potentially paves the way for phage entry into the bacterial colonies by destroying the protective layer of the biofilms. Additionally, the enzyme potentially enhances phage proliferation by weakening the bacterial cell walls to release phage progeny to infect other bacterial cells, consequently resulting in bactericidal activity. For instance, Schuch et al., 53 in their assessment of phage-derived lysin CF-301 in clearing bacterial biofilms under different conditions, saw a complete eradication of Staphylococcus aureus biofilms that had developed resistance to high concentrations of antibiotics, such as ciprofloxacin and daptomycin. Furthermore, in an in vivo experiment testing the use of phage CF-301 in the treatment of S. aureus infections, found that low concentrations of the phage were shown to be effective in destroying all biofilms present as well as eliminating free-floating bacterial cells. 53 Even though persister cells make up a significant fraction of biofilms, 54 and bacteria can exhibit a reduced metabolic activity within biofilms, phages retain the activity to lyse bacterial cells, including persister cells.47,51,55
Moreover, phage itself can prevent the formation of mature biofilms from planktonic bacteria or biofilm in its early stages (Figure 2). An illustration of this is phage Bxb1, which when integrated into the genome of Mycobacterium smegmatis, inactivates the groEL1 gene. This integration allows the bacteria to maintain a planktonic state, preventing the formation of mature biofilms. 56 In addition, many other phage-derived enzymes (summarized in Table 1), including hydrolases like glucanase, levosidase, sialidase, rhamnosidase, and xylosidase, along with lyases such as alginate lyase, hyaluronidase, and pectin lyase, are able to selectively identify, attach to, interact with and degrade EPSs, polyanionic carbohydrates, and extracellular DNA produced by the host bacterial cells.19,57–59 This enzymatic activity disrupts the structural integrity of the biofilm, thereby facilitating the infiltration of phages into the deeper layers of the biofilm and subsequent interaction with the enclosed bacterial cells, and further allowing the immune system to degrade bacterial cells (Figure 3).59–61 An illustration of this is the recent identification of lytic IME180 phage depolymerase activity on the degradation of Pseudomonas aeruginosa 1193 biofilm by degrading the exopolysaccharide of the biofilm, enhancing bactericidal activity mediated by serum complement proteins. 62 These hydrolases and lyase enzymes function differently, the hydrolases catalyze hydrolysis reactions, where the EPS molecule is broken down by the addition of water, while the lyases catalyze lytic or elimination reactions, where EPS components are cleaved or rearranged. All this evidence suggests that increasing the scope of the arsenal of antibiofilm activities of phages to make therapeutic application efficacious is highly likely.
Summary of phage encoded enzymes that degrade bacterial biofilms.


The interaction of phage enzymes with biofilm. (a) Enzymatic degradation of biofilm components by various phage enzymes and exposing naked bacterial cells. (b) Phages and lysins attach to and lyse naked bacterial cells.
Additionally, a selective combination of several lytic phages of different host range(s) into a single-phage cocktail formulation also provides a multifaceted interaction of phages with biofilms. While some phages in cocktails produce depolymerases, other phages that are lysis-producing or lysozyme-producing also generate their respective enzymes. This induces a simultaneous attack on biofilm components, and when bacterial cells are exposed, phage tails bind to specific receptors on bacterial cells to undergo the lytic life cycle. For example, Chhibber and his team reportedly reduced the mixed-species biofilm of Klebsiella pneumoniae and P. aeruginosa by utilizing a phage cocktail therapy comprising of two specific phages: Pa29, a non-depolymerase-producing phage which targets P. aeruginosa, and KP01K2, which targets K. pneumoniae and produces depolymerase enzymes. Utilizing a phage with degrading enzymes, the outer structure of K. pneumoniae was hydrolyzed, allowing Pa29 to access P. aeruginosa located beneath the biofilm. As a result, there was a significant reduction in the biofilm biomass for both bacteria, a challenge that may have been difficult to eliminate without phage cocktail therapy. 63 Emphatically, another study revealed that the utilization of a three-phage cocktail, including certain phages with polysaccharide depolymerases, exhibited superior efficacy in inhibiting and eradicating biofilms of 2–3 strains of Proteus mirabilis compared to individual phages, without any observed interference in the respective phage activities in the cocktail. 64
Bacteriophage against biofilms in patients and clinical isolates
Clinical isolates in vivo and animal models
Due to ethical limitations on the use of phage therapy in patients except as compassionate treatment in some countries, 65 its application remains largely limited to in vivo studies, where it yields encouraging results in many cases (Table 2). In the application of phages to control bacterial biofilms, there is an uneven distribution of studies across bacterial species. The strong biofilm formation of P. aeruginosa has made it a primary target for phage antibiofilm therapy, leading to more studies on P. aeruginosa infections compared to other bacterial infections.
Phage-mediated treatment of biofilms in preclinical studies; animal models and ex vivo or in vivo studies.
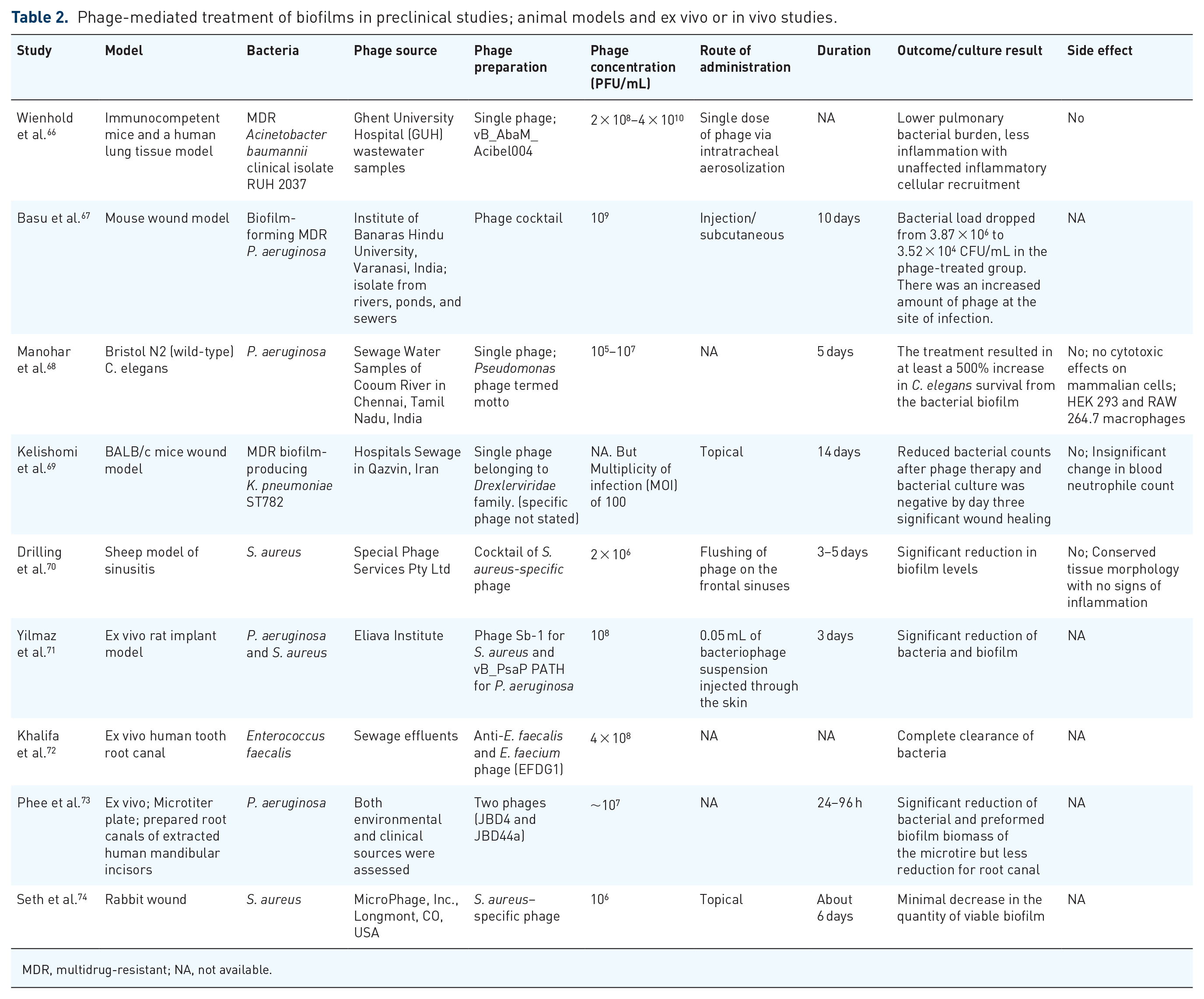
MDR, multidrug-resistant; NA, not available.
P. aeruginosa biofilm
Efficacy of monophage formulation against clinical isolates of P. aeruginosa biofilms
In a case report that attempted to determine the required minimum dosing of phage OMKO1 for the removal of P. aeruginosa biofilm, Chan et al. observed high effectiveness of phage OMKO1 in reducing the density of biofilms even at very low relative frequencies of phage particles. This was observed in an in vitro biofilm reduction assay after OMKO1 of 1011 PFU/mL was applied on biofilms of cell densities at an optical density of 600 nm (OD600) using isolates from the patient’s fistula discharge. These compelling findings led the authors to suggest that a single treatment by use of phage OMKO1 at only 103 PFU could potentially be sufficient to efficiently reduce a biofilm consisting of approximately 108 CFU/mL of the P. aeruginosa patient strain. 75
Jeon et al. conducted a comprehensive in vitro, in silico, and in vivo study, utilizing acute pneumonia mouse models to assess the efficacy of two novel phages (BΦ-R1836 and BΦ-R656) in fighting XDR P. aeruginosa infections. Both phage types demonstrated robust inhibitory activity, effectively lysing XDR P. aeruginosa strains isolated from pneumonia patients. The establishment of two distinct in vivo infection models by the researchers resulted in unequivocal findings, demonstrating the successful elimination of XDR P. aeruginosa strains by phage BΦ-R1836 and phage BΦ-R656 in Galleria mellonella larvae and acute pneumonia mouse models. The findings further demonstrated notable effectiveness in extensively eradicating XDR P. aeruginosa biofilms within the host. As a result, these significant observations led the authors to propose the potential utilization of these phages as biocontrol agents, demonstrating the ability to not just hinder the formation of biofilms on medical equipment and in healthcare settings but also efficiently eradicate infections associated with biofilms within the body. 76 In a separate simulated investigation, the efficacy of the phage PELP20 was assessed in combating chronic lung infections caused by P. aeruginosa in a mouse model, as well as in a simulated cystic fibrosis lung-like environment associated with biofilm formation. The authors noted a significant reduction of 3-log in the number of CFU of P. aeruginosa recovered from the phage-treated biofilm (after 24 h), suggesting that PELP20 possesses the ability to effectively infiltrate and eradicate bacteria within biofilms associated with a cystic fibrosis lung-like environment. 77
Efficiency of phage cocktail targeting clinical isolates of P. aeruginosa biofilms
In one of their studies, Debebe and his team investigated the efficacy of two phages (ΦNH-4 and ΦMR299-2) in combating nine clinical P. aeruginosa biofilms in murine lungs. Their findings showed that the two phages effectively eradicated all their clinical isolates within 6 h. The phage cocktail exhibited potent antimicrobial activity against both mucoid and non-mucoid strains grown in the cystic fibrosis bronchial epithelial cell line CFBE41o-. Furthermore, the phage cocktail exhibited a lethal effect on P. aeruginosa in murine lungs, effectively clearing the bacteria from the lungs within 6 hours. The authors attributed this efficacy to the phages’ ability to penetrate the exopolysaccharide of the biofilms, which seemed to require a longer exposure time (22–24 h) to clear the P. aeruginosa from the infected rat lungs. Additionally, they proposed that employing a variety of phages in a cocktail often targets different bacterial receptors, making it more challenging for bacteria to develop resistance. Consequently, employing a combination of different phages can effectively control bacterial populations by minimizing the emergence of resistance. 78
Moreover, in their 2018 study, Forti et al. designed a six-phage cocktail, including two previously examined phages (PAK_P1 and PAK_P4) and four newly discovered phages (E217, PYO2, E215, and DEV), that effectively cleared MDR strains of P. aeruginosa bacteria (PaPh5 and AA43) isolated from clinical samples. The release of phages resulted in near-complete eradication of biofilm formation within 48 h. In an attempt to determine the ability of phage cocktails to cure acute P. aeruginosa respiratory tract infections in mice, the authors observed that the utilization of phage cocktails with varying MOIs, even at very low MOI (an MOI of 0.05 for each phage), reduced the P. aeruginosa infections in mice in 48 h and resulted in a 100% survival rate. Additionally, the authors found appropriately timed phage cocktail administration as effective in rescuing P. aeruginosa-infected wax moth (G. mellonella). 79 The authors further found that administering the cocktail to the wax moth larvae before bacterial infection offered a prophylactic effect. 79 Analogous to the benefits of phage cocktail, Khalifa et al. demonstrated the effectiveness of the phage cocktail of two phages (EFDG1 and EFLK1) against planktonic and biofilms of vancomycin-resistant E. faecalis V583 (ATCC 700802). The phage cocktail showed an additive activity on resistant E. faecalis V583 strains compared to single phages, even against EFDG1r, which is a phage-resistant mutant strain. 80 Likewise, Abou Zeid et al. formulated a blend of four lytic phages (ΦEcp1, ΦEcp2, ΦEcp3, and ΦEcp4) that effectively suppressed the proliferation of highly biofilm-producing MDR E. coli strains and significantly impeded biofilm formation compared to the use of individual phages. The phage mixture exhibited broad infectivity, a substantial reduction in biofilm biomass, and achieved a 87% inhibition of biofilm formation while maintaining stability under environmentally challenging conditions (pH range: 7–9, temperature range: 55°C–65°C). 81
An in vitro study aimed at testing the activity of environmentally isolated phage against P. aeruginosa isolates from patients with cystic fibrosis identified that five selected lytic phages (Φ4_ZP1, Φ9_ZP2, Φ14_OBG, Φ17_OBG, and Φ19_OBG) showed significant wide host activity. 82 In another in vivo study, Fong and his team examined the efficacy of a four-phage cocktail against biofilms formed by P. aeruginosa isolates obtained from patients with chronic rhinosinusitis (CRS). The authors carried out an in vitro experiment where biofilms were treated with the phage cocktail, followed by a subsequent measurement of the biofilm biomass after 24 and 48 h with the aid of crystal violet assays. The results demonstrated that the cocktail treatment at concentrations of 108 and 109 PFU/mL significantly reduced biofilm biomass at both time points. 83 In a similar vein of compounding the benefits of cocktail application, the isolated and studied phages ΦKpnM-vB1, ΦKpnP-vB2, and ΦKpnM-vB3 demonstrated not only lytic capabilities against K. pneumoniae and E. coli strains but also proved highly effective in reducing Klebsiella biofilms when administered as a combined cocktail formulation. 84 These findings demonstrate that the application of phage cocktail formulation on biofilms holds promise in targeting a variety of bacterial strains and enables efficient biofilm control by preventing the emergence of resistant strains.
Efficacy of phage formulation against E. faecalis, S. aureus, and A. baumannii biofilm
In addition to effectively lysing significant species of bacteria (Table 2), phages have demonstrated immense antimicrobial activity against biofilm-producing A. baumannii strains, highlighting their therapeutic potential in addressing antibiotic-resistant A. baumannii infections, particularly in urinary tract infections. 85 A study by El-Atrees and his team identified four phages (EPA, EPB, EPD, EPF) that have the myovirus morphology and two (EPC, EPE) portraying the siphovirus morphology, all at titers of 107 PFU/mL, were found to have a promising antibiofilm activity against strong biofilm-forming urinary tract E. faecalis isolates (OD range 0.24–2.66), which is commonly found in urinary tract infections. The application of these phages resulted in a significant reduction of both preformed and newly formed biofilms, ranging from 38.02% to 45.7% and 71.0% to 80.0%, respectively, compared to the control group. 86 Another identified lytic staphylophage, Stau2, characterized by its double-stranded DNA, demonstrated the ability to lyse 80% of clinical S. aureus isolates (164 out of 205). 87 It is worth highlighting that complete lysis was observed within 3 h post-phage treatments in most isolates, and experimental mice infected with S. aureus showed 100% protection with Stau2 treatment. 87 Although it was not tested on biofilm-forming clinical isolates, the wide host range of the phage suggests its potential for biofilm applications.
Bacteriophage against biofilms within patients
Biofilms play a crucial role in enabling bacteria to endure in hostile environments, contributing to chronic infection, 88 as well as facilitating the dispersal of bacteria to different sites, leading to the initiation of new infections. 89 Phages are biocompatible and can be cleared via hepatic elimination by phagocytosing phages, making them a favorable applicable therapeutic agent. However, phages are challenged by with shortfalls such as short half-life and instability under varying pH.90–92 Concerning these shortfalls, the mode of phage administration to patients, which greatly influences phage–antibiofilm activity and adsorption, needs to be carefully determined based on the nature and location of the infection. 93 The findings from several preclinical studies suggest that to achieve a significant reduction in biofilms, it is necessary to administer phage directly to the precise location of the biofilm infection, 47 as observed in wound infection, where topical administration is the preferred mode of therapy.94,95 However, this idea becomes difficult to adopt when the site of infection is an internal part of the body and must therefore rely on indirect application or systemic delivery. Intravenous, intraoperative, and intrarectal phage administration are relatively unproblematic.93,95,96 These routes provide direct access to the bloodstream and relatively facilitate effective phage penetration into various tissues, particularly those affected by infection-induced inflammation, enhancing the permeability of the endothelial barriers as demonstrated in humans96,97 and animal models such as rats.93,98 Oral administration of phages is also quite effective, but inconsistent levels of different phages in the bloodstream after oral administration have been reported. Its adoption in clinical trials should therefore be preceded by preclinical experiments. 93 In the case of respiratory (lung and other pulmonary) biofilm-related infections, inhaled formulations (nebulization and other hydrogel formulations) have yielded promising results (Table 3), even though the loss of phage viability may be encountered. 99 Successful phage treatment via any of these routes is warranted by a variety of phage concentrations ranging from 104 to 1010 (Table 3).
Phage-mediated treatment of biofilms in human studies only.

Shown only are studies in which biofilm presence and treatment have been explicitly indicated.
MDR, multidrug-resistant.
Bacterial biofilms have been cleared in many parts of humans (summarized in Table 3), including, but not limited to the skin, femur, skeletal muscle, lungs, genitourinary tract, and biofilm-contaminated invasive devices. One such piece of evidence is a case report by Cano et al. 103 in which in vitro experimentation and subsequent therapeutic application on a 64-year-old patient was carried out at the Mayo Clinic Clinical Research and Trials Unit, a recovery of the knee function was observed. In the preclinical study, the treatment of K. pneumoniae with 200 μL of phage KpJH46Φ2 at a concentration of 1.26 × 109 PFU/mL led to a reduction in biofilm biomass. Following this convincing result, the authors proceeded to administer phage KpJH46Φ2 to a 62-year-old male with a history of diabetes, obesity, and bilateral knee osteoarthritis complicated with numerous bacterial infections. Daily intravenous administration of KpJH46Φ2 at 6.3 × 1010 PFU in 50 mL of normal saline for a total of 40 doses against the biofilm-forming K. pneumoniae complex isolated from the patient was performed. Just after 22 h of phage administration, a trend in biofilm biomass reduction was observed. The therapy resulted in the resolution of local signs and symptoms of infection, increased range of motion, reduced inflammation, and recovery of function without the patient experiencing treatment-related adverse effects. 103
Another convincing outcome of lytic phage therapy was observed in another case study on a 72-year-old male with morbid obesity and hyperlipidemia with chronic MRSA prosthetic joint infections. 107 To successfully sterilize the patient’s chronic MRSA from the prosthetic joint infection, two doses of a selectively purified phage SaGR51Φ1 at 5.4 × 109 PFU were intra-articularly administered in 10 mL of normal saline at the end of surgery, after which antibiotic (IV daptomycin 1000 mg) administration was started and the subsequent daily intravenous phage (5.4 × 107 PFU/mL) administration. After 2 months, intraoperative cultures were again culture-negative.
There are documented instances of successful oral biofilm elimination, which is a key objective in preventing tooth decay. Phage SMHBZ8, obtained from samples of human saliva, demonstrated significant efficacy in preventing the formation of biofilm and reducing existing biofilm of S. mutans in both biofilm cultures and a cariogenic dentin model. 108 In this domain, Khalifa et al, characterized a lytic phage, an anti-E. faecalis and E. faecium phage (termed EFDG) obtained from sewage. These EFDGs were seen to efficiently infect and kill planktonic and biofilm cultures of E. faecalis in vitro and efficiently prevent ex vivo E. faecalis root canal infection of an extracted one-rooted human teeth. 73
Recent advances by which bacterial cells embedded in biofilm can confer additional resistance against phages
Apart from the general inherent resilience of bacterial biofilms as shields against antibacterial agents (including antibiotics and phages), bacteria within biofilms employ a strategy of upregulating specific essential biofilm substances to capture viral particles, hindering phage invasion and safeguarding bacterial cells from potential damage.23,109 This is highly unfavorable to phages due to their movement being Brownian Motion. In addition to the shielding mechanism of biofilms, the bacterial cells within the biofilm also have recently been observed to protect themselves against their viral predator phages in a multifaceted manner: the release of phage-receptor decoys, release of outer membrane vesicles (OMVs), altering or concealing viral receptors, superinfection exclusion (Sie), retron production, expression of DNA-intercalating metabolites, use of toxin–antitoxin system, use of restriction-modification (R-M) systems, BREX system, abortive infection (Abi) systems, CRISPR-Cas and other mechanisms to counter the invasion of phage DNA (Figure 4).19,32,38,110–118 These novel defense systems possess a wide range antiphage defense mechanism, as noted by Ofir and his group in the case of a defense island system associated with restriction-modification (DISARM) in restricting incoming phage DNA. 113 Many of the defense systems exhibit synergistic antiphage activity. In this domain, Wu et al. 119 demonstrated that Zorya II synergizes with Druantia III and ietAS defense systems, while tmn exhibits synergy with co-occurring systems Gabija, Septu I, and PrrC.

Mechanisms of bacterial resistance to phage.
Release of phage-receptor decoys, OMV, and altering or concealing viral receptors are important separate mechanisms adopted by bacteria, enabling them to survive in the presence of phages. The outer membrane of Gram-negative bacteria has phage receptors that can be secreted and serve as decoys, causing phages to irreversibly bind to these decoys. This results in futile infection attempts by the phages. An example of this phenomenon is observed in Vibrio cholerae, where OMV from the bacteria inhibits phage infection.115,120 Studies assessing the evolution of bacterial resistance have identified that bacteria render themselves less susceptible to phage attachment and infection by altering the surface structures or receptors that phages typically recognize and bind to Refs. 116 and 121. This strategy hampers the ability of phages to recognize and infect bacterial cells, providing a means of defense.
Expression of phage DNA-intercalating metabolites has been reported among some bacteria to confer antiphage activity. A study investigating molecules that inhibit phage φScoe2 and φScoe25, as well as E. coli phage and P. aeruginosa phage replications, identified eleven compounds, nine of which were DNA-intercalating agents. Among these were a variety of naturally produced secondary metabolites of Streptomyces spp., including daunorubicin, doxorubicin, epirubicin, and idarubicin. 122 The DNA intercalation is suggested to prevent the circularization of phage linear DNA, 35 thereby inhibiting the integration of phage DNA into the bacterial DNA.
Sie is a bacterial defense phenomenon in which a preexisting viral infection prevents subsequent infections by closely related viral particles. 123 Bacteria that are capable of this mechanism can restrict the entry of additional phages into an already infected cell by producing proteins to hinder the secondary infection. For instance, protein Imm and protein Sp encoded by phage T4 prevent subsequent infection of the same bacteria by other similar T-even phages by ensuring the bacterial peptidoglycan is free from degradation, thereby hindering the phage DNA transfer across the membrane.124,125 When a Sie is bypassed and the bacteria are infected, Abi systems are triggered in some bacterial cells; a self-destruct mechanism that leads to the premature death of the infected cell (“suicide of the infected bacteria”).19,126 The precociously death of the infected cells prevent the concomitant replication and spread of phages, safeguarding the rest of the bacterial community from phage invasion.126,127 Additionally, retrons, which are bacterial genetic elements that produce small RNA molecules that can interfere with viral replication or other viral processes also activate their Abi system. This was evident when Millman et al. examined multiple retron systems and demonstrated their ability to confer defense against a wide array of phages through the mechanism of abortive infection. 112 Furthermore, some bacteria employ toxin–antitoxin systems as a defense mechanism to counter phage invasion 128 but the mechanism of action underlying this is currently elusive.
R-M, BREX, and CRISPR-Cas systems are separate and highly diverse mechanisms adopted by bacteria to enable them to survive phage predations via recognition and cleavage of phage DNA at specific sites while protecting their own genomes.19,129 R-M systems, comprising restriction endonucleases and methyltransferases as their two essential components, are vital defense mechanisms adopted by bacteria in preventing phage DNA invasion. The endonucleases recognize and cleave specific DNA sequences within the phage genome, rendering it nonfunctional, and further providing innate immunity against foreign genetic material. To prevent the bacteria’s DNA from being damaged by the cleaving ability of the endonuclease, methyltransferases serve a protective role in the R-M systems, methylating the host bacterial DNA at the same recognition sites, safeguarding it from the activity of the restriction endonucleases.130,131 Similar to the R-M system, BREX system also prevents phage genome replication, but its underlying mechanism is yet to be elucidated. It comprises methyltransferase (BrxX) and ATPase (BrxC). The BrxX is said to methylate adenine in the bacterial DNA, 118 while the latter may be providing energy to the system. This methylation is proposed to be the distinguishing feature of the bacterial genome and the phage genome. 132 CRISPR-Cas systems are adaptive immune defense systems in some bacteria, providing defense against phage and plasmids. These systems consist of CRISPR arrays, which are composed of short repetitive sequences interspersed with unique spacer sequences derived from previous phage encounters. When a phage invades, the Cas proteins associated with the CRISPR array use the spacers as a guide to recognize and cleave the phage DNA, effectively neutralizing the phage while maintaining the integrity of the bacterial genome.133,134
In addition to these underlying and novel resistance mechanisms, selecting the type of phage for biofilm eradication should be highly regarded, as it may enhance biofilm formation or tolerance. For instance, if selectively lytic phages are not carefully chosen, filamentous phages might be mistakenly used, resulting in the enhancement of bacterial stability and their biofilms. An example includes the interaction of filamentous phage with host and microbial polymers, resulting in the formation of a more complex liquid crystal structure within the biofilm matrix, thereby improving adhesion, survival in dry conditions, antibiotic resistance, tolerance, virulence, and dispersion capabilities of the biofilms.135,136
Mechanisms by which phages overcome bacterial resistance
To counter these resistance mechanisms, some phages rapidly evolve to carry genes encoding anti-R-M and anti-CRISPR proteins like AcrF1-5 (extensively reviewed in Refs. 137 and 138)), while other phages introduce mutations into the specific sequences targeted by CRISPR-Cas systems, including protospacer.137,139 Phages have also evolved to encode antitoxins produced by their host. For instance, ToxN toxin produced by E. coli against phages is inhibited by TifA protein produced by T4 phage. 140 A recent study also identified phage ICP1 protein (AdfB) to nullify DarTG, an antiphage toxin produced by V. cholerae. 141 Recently, the T7 phage protein Ocr has been identified as a novel protein that counteracts the BREX system, 142 which is an antiphage defense mechanism. Ocr achieves this by suppressing the enzymatic activity of BrxX. 118
Moreover, in response to bacteria concealing phage-binding receptors and releasing phage-receptor decoys, some phages have evolved to recognize alternative receptors on bacterial surfaces. 143 In superinfection, where a first-time infection prevents subsequent infection by similar phages, the rapid phage mutation rate has enabled the bypassing of these bacterial defense mechanisms. For instance, the phage P22 encodes the SieA protein, which blocks DNA injections by other P22-like phages. However, mutant strains of phageP22 with amino acid changes at the C-terminal regions of the gene 16 and gene 20-encoded ejection proteins can overcome this block. 144
In cases where bacteria completely resist their predator phage, they often experience a fitness cost. This trade-off can render the bacteria more susceptible to other phages with different mechanisms or other antibacterial treatments due to the metabolic burden of maintaining energy and resources for the resistance. 145 Essentially, the metabolic burden of bacterial resistance mechanisms can slow down the bacterial growth rate, as observed in antibiotic-resistant bacteria. 146 While the bacteria may become resistant to one type of phage, they may become deficient in virulence traits, 147 including biofilm formation. 148 A recent study confirmed impaired biofilm formation of phage-resistant A. baumannii AB900 and A9844 following their defective capsule production to avoid predation from their respective phages, ΦFG02 and ΦCO01. 149 In addition to the impaired biofilm formation, these phage-resistant A. baumannii strains were resensitized to beta-lactam antibiotics and the human complement system, as well as other phages. 149
Future perspectives on optimizing phage therapy against biofilms
The key to effectively treating biofilm-associated bacterial infections lies in preventing the biofilm maturation from the early stages of development, allowing conventional antibacterial agents to easily access the nascent bacterial cells. However, when dealing with preformed or mature biofilms, the successful eradication of biofilms depends on the capacity of antibacterial agents to rupture or penetrate the bacterial biofilm involved (Figures 2 and 3). Unfortunately, only a limited number of antibiotics possess such remarkable capabilities. Therefore, optimizing the antibiofilm activity of phages would be of utmost importance in advancing its therapeutic application in the near future (Figure 5).

Perspectives on enhancing the antibiofilm activity of phages.
Phage cocktail
The utilization of phage cocktails, harnessing the combined effect (sometimes synergistic) of individual phages, potentially enhances phage adsorption and spread within bacterial biofilms to lyse bacterial cells, as elaborated earlier. An example of this is when a phage enzymatically (e.g., depolymerase) breaks down the EPS structure of a multispecies biofilm, including those produced by its host and closely related species. This enzymatic action potentially improves the accessibility of other phages to their respective hosts within the biofilm.50,63,150
Furthermore, considering that, the isolation of phages that can lyse a variety of hosts (broad host range) is increasing, combining broad-host-ranged phages, such as the novel phages (e.g., ΦParuNE1 151 and HZ2201 152 known for their respective potent antibiofilm activity) into a phage cocktail can result in a highly effective treatment against a range of pathogens. This approach reduces the number of phages in the cocktail while maintaining their efficacy, simplifying the purification and safety assurance processes. Utilizing a smaller number of phages with a broad host range can also lead to cost savings in the formulation of the phage cocktail.
Genetic engineering as a beneficial tool for phage therapy
Harnessing the power of genetic engineering to address the limitations of phages holds tremendous potential for the effective eradication of biofilm infections. Incorporating genetic engineering techniques that augment phage shortfalls would empower phage therapy to overcome the inherent shortcomings in combating and eradicating bacterial biofilms. Phages can be engineered to incorporate anti-CRISPR proteins and anti-R-M system proteins to avoid phage DNA cleavage,153,154 as well as antitoxin proteins to neutralize bacterial secreted toxins. However, before these features come into play, phages must disrupt or penetrate the biofilm to reach the individual bacterial cells. In this regard, phages that lack genes encoding degradative enzymes can be genetically modified to express such enzymes. This enhances their ability to adsorb onto, penetrate, and diffuse through the exopolysaccharide matrix, ultimately facilitating biofilm destruction. 155 For instance, a study by Pei & Lamas-Samanamud engineered phage T7 to incorporate a lactonase enzyme with a wide-ranging capability to quench quorum sensing. This cell–cell communication mechanism (quorum sensing), mediated by acyl homoserine lactones (AHLs), is crucial for biofilm formation, particularly in the early stages. In this study, the authors demonstrated that an engineered T7 phage expressing the AiiA lactonase effectively degraded acyl homoserine lactones from various bacterial species. When introduced to mixed-species biofilms containing P. aeruginosa and E. coli, this engineered phage significantly inhibited biofilm formation. 156 More innovatively, Lu and Collins designed and engineered a phage T7, an E. coli-specific phage, to be capable of expressing the DspB enzyme, which is set to be released extracellularly upon cell lysis to degrade the β-1,6-N-acetyl-D-glucosamine (a crucial adhesion component for biofilm formation) during infection. 157 The assessment of the modified phage’s effect on E. coli biofilms yielded impressive findings. Upon phage infection, the efficient expression of the biofilm-dispersing enzyme (i.e., DspB) resulted in a nearly absolute biofilm degradation rate (~99.99%). Furthermore, there was a substantial decrease in the population of viable bacterial cells within the biofilm, surpassing the efficiency of the parent T7 phage. These notable outcomes were observed within 24 h, and no instances of phage resistance were detected. 157 Analogously, Yang et al. also highlighted the potential of an engineered lysin ClyR as a preventive or therapeutic agent for the prevention of dental caries caused by Streptococcus mutans, as it has been demonstrated to have high activity against S. mutans biofilms both in vitro and in vivo. 158 Furthermore, there is compelling evidence that the recombinant depolymerase P510dep (a prospective tail fiber protein derived from Przondovirus phage P510) demonstrates substantial efficacy in degrading the essential polysaccharides involved in carbapenem-resistant K. pneumoniae biofilms. 159 Additionally, an engineered LysAB2 exhibited notable activity against A. baumannii and demonstrated an exceptional ability to disrupt the formation of biofilm. 160
In another study by Wu et al. that isolated and sequenced a phage SH-KP152226 (encoding depolymerase Dep42) that exhibits a specific affinity for infecting and lysing K. pneumoniae capsular type K47, they highlighted the potential therapeutic application of Dep42 (a novel depolymerase) in effectively controlling infections caused by the K47 capsule of K. pneumoniae. This was observed after conducting a recombinant study with the Dep42 and noticed the depolymerization of the K47 capsule of K. pneumoniae with an additional notable disruption of the preformed biofilms and in some cases, inhibition of biofilm formation. 161 In a similar context of engineering phage-derived enzymes, PM-477, an engineered endolysin derived from phages through domain shuffling, has demonstrated the ability to effectively eradicate Gardnerella spp. and physically disrupt its biofilms without affecting lactobacilli and other species of vaginal bacteria flora. 162 The recent identification of the Ocr protein, which nullifies the BREX system, 118 offers new possibilities. Genetically engineering Ocr into other phages enhances their defense against a wide range of bacteria that rely on the BREX system. Quorum sensing has recently been associated with inhibiting phage infection via biofilm regulation. 163 Therefore, engineering phages to encode broad-spectrum quorum quenching molecules would potentially enhance their ability to disrupt bacterial communication systems, leading to more effective treatments against biofilm-associated infections and reducing the likelihood of bacterial resistance development.
Phage-based supramolecule development
A promising avenue for combating biofilms lies in the development and utilization of phage-based supramolecules (PBS). These supramolecules can be crafted by amalgamating phages with enhancing agents and other antibacterial components, like antimicrobial peptides, to cause direct damage to bacterial cells or as a delivery method of the phage. The resulting supramolecules exhibit a diverse range of forms, including broad-spectrum variants, while possessing inherent antibiofilm properties. This unique characteristic empowers these supramolecules to disrupt and ultimately eliminate both single and mixed bacterial species residing within biofilms.164–166 An important feature of developing phage-based supramolecules is the opportunity to alter the overall size of the developed supramolecule, as it offers larger multivalent surfaces that can be manipulated to provide selectivity against bacteria. Controlling the composition, size, and surface properties of these PBS offers a considerable tendency to generate an antibacterial agent with a diverse array of antibacterial activities. In a recent study, Rotello et al. 167 described that cationic nanomaterials are capable of penetrating biofilms. Synthesizing a supramolecule made of these nanomaterials coupled with appropriate phages may yield a promising antibiofilm effect. This is demonstrated using phage T4 with chitosan and polyethylene imine-coated alginate beads to treat E. coli. The T4 phage (108 PFU/mL) was encapsulated in alginate beads in the presence of 2.0% (w/v) sodium alginate and CaCl2 solution. 168 Similarly, Chhibber et al. 169 also used PC: Tween80, cholesterol, and liposomes with lytic phages, MR-5 and MR-10, in the treatment of MRSA-infected rat wounds.
While the clinical implementation of PBS has been limited due to safety concerns, individual components of PBS have been shown to be independently biocompatible with human cell lines through in vivo and in vitro studies. This is demonstrated in a recent study where the combination of phage T7 and AgNPs was highly effective on bacterial biofilms and was safe for Jurkat E6-1 cells and adult human dermal fibroblasts, except that AgNP was observed to be toxic at a very high dose of 0.1 mg/mL. 170
Limitations and challenges of the prospects in developing phage therapies for clinical use
These perspectives on enhancing phage–antibiofilm activity hold promise, but they are not without limitations. Safety concerns surrounding phage therapy remain significant due to our limited understanding of its pharmacodynamics and pharmacokinetics. In addition to the controversies on strict ethical regulation of genetically modified therapies, 171 the genetic modification of phages to express degradative enzymes and the use of phage cocktails may face challenges such as the complexity of ensuring consistent and effective phage delivery to biofilms. Additionally, the clinical implementation of phage-based supramolecules may be hindered by safety concerns, including immunogenicity and potential toxicity, as well as the need for extensive research to validate their efficacy and biocompatibility.
Conclusion
This review has provided insight into the basic science and recent advances of phage therapy against biofilm-forming antibiotic resistance. It also highlighted some challenges with biofilm resistance to phages, as well as potential benefits of optimized phage administration systems. It is evident that phages are efficacious antibiofilm agents. However, the intricate interaction at the molecular level of how phages interact with biofilms is not fully exploited. Also, the biochemical interaction of phage enzymes with biofilms remains to be elucidated. Understanding these key concepts that underlie phage–biofilm interaction is imperative to the development of modified systems of phage-derived proteins and enzymes for combating clinically relevant antibiotic-resistant biofilms. In addition to phage cocktail and phage genetic engineering strategies, it would be promising to channel future studies into conjugating phages with other antimicrobial agents such as peptides to enhance phage antibiofilm efficacy.