
Abstract
Engineered three-dimensional (3D) in vitro and ex vivo neural tissues, also known as “mini brains and spinal cords in a dish,” can be derived from different types of human stem cells via several differentiation protocols. In general, human mini brains are micro-scale physiological systems consisting of mixed populations of neural progenitor cells, glial cells, and neurons that may represent key features of human brain anatomy and function. To date, these specialized 3D tissue structures can be characterized into spheroids, organoids, assembloids, organ-on-a-chip and their various combinations based on generation procedures and cellular components. These 3D CNS models incorporate complex cell-cell interactions and play an essential role in bridging the gap between two-dimensional human neuroglial cultures and animal models. Indeed, they provide an innovative platform for disease modeling and therapeutic cell replacement, especially shedding light on the potential to realize personalized medicine for neurological disorders when combined with the revolutionary human induced pluripotent stem cell technology. In this review, we highlight human 3D CNS models developed from a variety of experimental strategies, emphasize their advances and remaining challenges, evaluate their state-of-the-art applications in recapitulating crucial phenotypic aspects of many CNS diseases, and discuss the role of contemporary technologies in the prospective improvement of their composition, consistency, complexity, and maturation.
Keywords
Introduction
The field of neuroscience has proven to be one of the more mystical fields in the realm of science and with that has attracted some of the most brilliant minds. Research spanning subdisciplines of cellular and molecular neuroscience, systems neuroscience, cognitive and behavioral neuroscience, and computational neuroscience has gifted the world profound knowledge of the anatomy and physiology of the human brain. Furthermore, researchers immersing themselves in translational and clinical studies have been continuously pursuing innovative model systems to achieve valiant leaps into addressing some of the most complex central nervous system (CNS) diseases through study and trial for effective therapies, yet many of these disorders remain currently untreatable. While the field is rooted in the knowledge and insight that researchers bring with them, progress can only go forward as far as its tools and technologies will take it. There has been a continuous demand to minimize the gap between animal and human models in both basic and translational neuroscience for many years. 1 While animal models provide an in vivo system that is more complete for studying neural development, disease progression, and treatment efficacy, there are several key differences between animal models and humans that begin to manifest themselves on the most important scale: the patient. Therapies for a range of CNS disorders which were validated in animal models have often failed in human trials. 2 Therefore, more relevant human models, especially those generated from patient tissue, will significantly empower researchers to address human-specific disease mechanisms in a cost-effective and scalable manner. In addition, these human models will help shape novel cell transplantation strategies aimed at replenishing lost or damaged cells in CNS injuries and disorders. 3
Limited access to functional human brain tissues impedes a comprehensive understanding of brain development, disease mechanisms, and advancement of therapeutics.4,5 Thus, for years, neurobiologists have turned to a variety of in vitro and ex vivo approaches to study the complexity of the human brain and its molecular processes.4,5 However, the application of model organisms in the study of the human brain has not been fully informative due to remarkable differences between human brain development and that of other species. To date, most attempts at applying human cells or tissues to model CNS disorders have been based on two-dimensional (2D) cell cultures that do not include many aspects of the complex physiological environment seen in the CNS. These cultures do not always accurately represent how cells grow in a human brain and how they may be affected by CNS disease and injury. Thus, the desire to develop a system that can closely recapitulate the crosstalk between various cell types in the human brain has led to the employment of three-dimensional (3D) cultures based on human material. 3D tissue cultures have been in development since the early 20th century, as researchers for decades have realized their potential in disease modeling and drug development. 6 3D human CNS tissue cultures, also known as human mini brains and spinal cords, are a subset of 3D models that usually retain a higher degree of structural complexity and maintain physiological homeostasis for prolonged durations. Some of them also contain various cell types that influence intricate cell-cell interactions, an element essential for the regulation of cell differentiation and metabolization. Ultimately, decades of experimental effort along with a continuously growing understanding of human CNS development have enabled progressive advancements in the generation of human mini brains resembling human brain structures. The field has greatly benefited from the use of these 3D brain technologies in studying cell-cell interactions and the assembly of neuronal circuits. These new types of in vitro human brain models serve as alternatives to existing animal models and close the gap in recreating human specific physiology and pathology of CNS development and disorders.
One of the biggest challenges for obtaining robust human 3D CNS models has been the relative scarcity of pertinent foundational cellular components used in their production. Human pluripotent stem cells (hPSCs), especially human induced pluripotent stem cells (hiPSCs), have proved to be a milestone in the biomedical community. Through various reprograming and gene modification methods, researchers can use hiPSCs with specific genetic profiles as the building blocks to grow any somatic tissue or organ in vitro. 7 From a neuroscience perspective, a great deal of work to generate human mini brains and spinal cords has been done on the front end in developing tuned differentiation protocols that mimic developmental cues to pattern different cell types and regions of the CNS. However, to date, there is still a great deal to be discovered and improved in engineering in vitro human mini brain models. In this review, we characterize and summarize the three main types of 3D CNS models: neurospheres, CNS organoids, and CNS assembloids, based on their production strategies and structural complexity (Figure 1). We also describe the emergence and role of revolutionary organ-on-a-chip systems in modeling specific elements of the CNS. We explain how these diverse but related structures differ from each other, what specific purpose each of them aims to achieve, and where each can be improved. In addition, we discuss some of the supporting technology that may be used in concert with these 3D CNS models to help introduce critical physiological cues and support appropriate culturing environments. Ultimately, this review hopes to create some consistency in the field and inspire researchers to continue to bring creativity to the design and development of in vitro human mini brain and spinal cord modeling platforms.

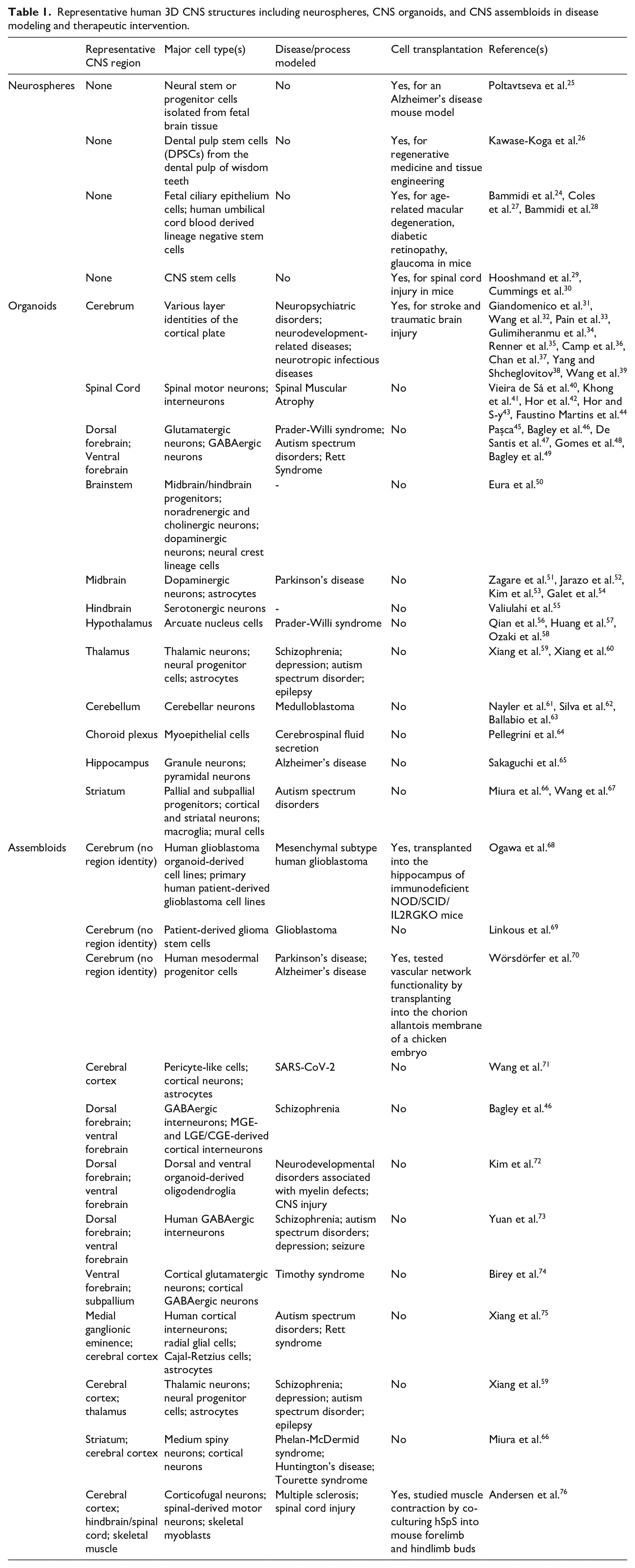
Human stem cells can be used to generate mini brain and spinal cord models. Human pluripotent stem cells (hPSCs), as well as a variety of adult stem cells derived from multiple sources in the body can be differentiated into neural progenitor cells (NPCs). NPCs can then be differentiated into several types of neural lineage cells and aggregated to form neurospheres. HiPSCs can also be aggregated into embryoid bodies, which can be patterned into region-specific CNS organoids, which can then be assembled to create CNS assembloids. Circuitoids are a subset of assembloids which recapitulate neural circuit systems such as the motor pathway and visual system.
Neurospheres: Simplified human CNS spheroids
Spheroids typically serve as an umbrella term for cultured 3D sphere-like tissue structures. The multifunctionality of spheroid cultures with relatively homogenous cellular components allows them to be used as rudimentary 3D cellular models. A variety of cells, including hepatocytes, stem cells, tumor cells, and neural cells have been applied to generate spheroids with restricted cell types. 8 3D human neural spheroids, commonly known as neurospheres, were initially developed as 3D cellular structures that were comprised of neural precursors or post-mitotic neurons derived from 2D cultures.9,10 The early 1990s saw the first culture of neurospheres derived from the periventricular area encompassing the subventricular zone in adult mice. 9 In turn, adult neural stem cells emerged from the niche study of adult neurogenesis and the in vitro study of multipotent precursors from the adult brain. 10 Since then, human neurospheres have been differentiated and developed from many types of human neural progenitors and other types of stem cells including ciliary epithelium cells, dental pulp stem cells (DPSCs), human umbilical cord blood (hUCB), and bone marrow derived mesenchymal stem cells (Figure 2). In general, these 3D multicellular spheroids, although mechanically simplified, manifest better structural representation and functional design than do 2D cultures, including dynamic cell-cell and cell-matrix interactions that represent some features of the CNS microenvironment.11,12 Due to their substantial advantages over 2D neuronal cultures, 3D neurospheres pioneered the way for the use of in vitro human mini brains and spinal cords for CNS disease modeling.13 –15

Neurospheres pave the way for studying the CNS in 3D. Neurospheres are generated by culturing neural progenitor cells (NPC) derived from various stem cell sources in low adhesion plates allowing the cells to self-aggregate. This 3D model is currently used for disease modeling via co-culturing with other 2D cells, drug and toxicity screening, and may one day be used for transplantation into humans.
Unlike in long-established 2D cultures, the absence of an attachment surface or scaffold allows mono-dispersed cells to undergo self-assembly and aggregate to form cell specific spheroids. 12 This self-assembly process somewhat mimics developmental cellular processes like embryogenesis, morphogenesis, and organogenesis.12,16 Spheroid cultures usually form a gradient of oxygen metabolites, soluble signals, and other essential nutrients necessary for an optimal in vivo like microenvironment. 8 Neurospheres can be generated from neural progenitor cells isolated from either adult or fetal CNS.17,18 They are generally created by culturing originating cells in low adhesion plates. 8 Low adhesion plates have an ultra-low attachment surface coating and a well-defined geometry (U-shaped bottom or V-shaped bottom) in each well to minimize cell adhesion and promote self-aggregation.19 –21
One of the many advantages of using neurospheres as an in vitro human CNS model is the ease with which internal and external cues can be manipulated, proving a better means for modeling different elements of the CNS. Notably, neurospheres may be co-cultured with 2D cells or some acellular matrices, which essentially provide contact-mediated directed differentiation cues. Such co-cultures are able to not only instruct neurospheres to reach specific phenotypes, but promote their reparative properties for therapeutic purposes. 22 This dynamic interaction was exemplified in a model in which neurospheres were developed from adipose stem cell derived neural progenitor cells (ASC-NPCs) followed by co-culture with acellular dermal matrix. Such co-culture was demonstrated as a better therapeutic strategy to repair peripheral nerve injuries when compared to using neurospheres alone. 23 Furthermore, neurospheres have been tested in several rodent transplantation studies targeting tissue regeneration in disease including Alzheimer’s disease, macular degeneration, and spinal cord injury (Table 1). Interestingly, transplanted neurospheres were better at restoring native cell populations compared to the transplantation of undifferentiated cells and helped improve certain behavioral phenotypes in cognitively impaired animals.24,25 Indeed, neurosphere cultures allow us to examine fetal or adult CNS cells outside of their normal environment while maintaining its integrity as a model system for developmental processes and therapeutic intervention. To date, several neurosphere phenotypes have been developed by different laboratories (Table 1). Despite its usefulness, there are significant drawbacks associated with neurosphere technology that limit it from being a frontrunner in modeling complex CNS environments. These limitations include variations in development, maintenance, and size, lack of cellular complexity in specific cell types, uncontrolled proliferation and differentiation of cells, disordered cellular aggregation, and imprecise and unstandardized high-throughput compatible assays for drug screenings. 8 While simple and technically friendly, neurospheres have paved the way for the development of more complex 3D mini brain models, and continue to find utility in the study of the CNS and associated diseases.
Representative human 3D CNS structures including neurospheres, CNS organoids, and CNS assembloids in disease modeling and therapeutic intervention.
CNS organoids: Self-organizing multicellular hierarchies mimicking distinct CNS regions
Human CNS organoids, including brain and spinal organoids, have drawn substantial attention in the field of neuroscience by becoming more relevant CNS models in relation to anatomical and physiological features of human brains and spinal cords. Unlike neurospheres, CNS organoids are mostly developed from hPSCs and usually represent multicellular structures that are hierarchically organized with sophisticated interstitial compartments. Various cellular phenotypes with orderly arrangement can be identified in these CNS organoids, including neural precursors, different types of neurons, astrocytes, and oligodendrocytes. To date, brain and spinal organoids have become popular modalities for studying region-specific structures, 3D-microenvironments, and entire organs for both the brain and spinal cord, and own an insuperable advantage over animal models by way of their human origin.31,40,41,77 The first contemporary human cerebral organoid model was established from human embryonic stem cells (hESCs) in 2013 and used for analyzing human brain development, specifically aimed at exploring microcephaly, a condition that cannot be precisely recapitulated in mouse models. 78 Inevitably, this model provided researchers a foundational platform for studying neurological diseases with human specific genetic roots, as well as allowed for the closer study of the unique development of the human CNS. Brain organoids are also a promising tool in healthcare’s shift toward personalized medicine. Through the reprogramming of patient peripheral blood mononuclear cells or skin fibroblasts into hiPSCs, clinicians can create patient-specific organoid models, in turn allowing them to better tailor treatment regimens to patient needs. 79
The generation of CNS organoids involves either direct or indirect neural induction, which starts with the aggregation of hPSCs into embryoid bodies (EBs). These EBs are then grown in suspension culture and patterned and matured in the presence of neuronal induction factors and small molecules that drive spatial organization and specific cell fate differentiation (Figure 3).41,50,64,80–83 As these organoids differentiate and mature, region-specific cell types and tissues begin to emerge including forebrain, midbrain, hippocampus, hypothalamus, spinal cord, etc. (Table 1).56,84 For instance, dorsal forebrain organoids contain significant amounts of excitatory glutamatergic neurons while ventral forebrain organoids contain a high proportion of inhibitory GABAergic neurons. 84 The potential interaction and connectivity between cell types within brain organoids makes them more advanced models than neurospheres, especially when studying the communication between different cell types in a specific brain region. Thus, brain organoids with specific region identities allow for significant external control with limited variation and reinforce the integrated development of a variety of CNS cell types in a way that cannot be achieved with more basic spheroid models, such as neurospheres.

CNS Organoids can be patterned to recapitulate various regions of the brain and spinal cord. hiPSCs and hESCs of different origins can be aggregated to form embryoid bodies, which through the addition of various factors and inhibitors, can be pattern into neuroectodermal-committed organoids. Using unique patterning factors, organoids can be further differentiated toward different regions of the brain or spinal cord. These organoids are useful for modeling neurological and neurodegenerative diseases and can be applied in drug screening studies. They also show potential for use in human transplantation after brain or spinal cord injury.
Spinal cord organoids, a subset of CNS organoids, are usually induced by morphogens and growth factors that caudalize the CNS development. Retinoic acid is introduced to drive differentiation toward spinal motor neurons, while sonic hedgehog (SHH) is used to mimic ventral signals and drive spatial organization of the spinal cord. Growth differentiation factor 11 (GDF-11) is used to increase rostro-caudal patterning of the spinal cord as well as drive differentiation towards motor neurons and interneurons, which may provide either excitatory or inhibitory signals to sensory neurons and other glia.40,41,77 Indeed, the extreme complexity of the human spinal cord structure may intimidate the development of human spinal organoids, as the spinal cord not only demonstrates an intricate separation of gray and white matter, but also contains a wide variety of interneurons and motor neurons heterogeneously distributed along its length. Despite some advances in their development, the generation of 3D spinal cord tissue from hPSCs may ultimately require assembling region-specific brain organoids, as well as peripheral neural ganglia in more complex constructs.
Human CNS organoids have been applied to: (1) disease modeling and drug screening for familial and sporadic neurodevelopmental and neurodegenerative disorders, as well as CNS injuries, (2) elucidating human CNS development for neural differentiation and migration, (3) evaluating neurotoxicity induced by a wide variety of chemicals and pathogens, and (4) testing tissue replacement therapies.78,85
–87 Engineered brain organoid
Despite advances in CNS organoid modeling strategies, there are still several limitations and technical challenges that hinder CNS organoids from accurately modeling the entire complexity of the CNS. A lack of essential cell types, including microglia and sufficient myelinating oligodendrocytes, limit the study of many CNS diseases such as autoimmune encephalitis, which is implicated by neuroinflammation-trigged neuronal death in the brain. 103 Additionally, a lack of vasculature components not only diminishes the structural complexity of CNS organoids, but restricts their culture duration, expansion, and maturation, partially due to the insidious development of cellular necrotic cores within the organoids. Compensatory strategies that use bioreactor devices and orbital shakers have shown some relieving benefits but have not done enough to completely overcome these hurdles. To date, even under the most optimized culture conditions, the size and scale of CNS organoids remains only a fraction of that seen in the human brain. One of the biggest concerns in applying human CNS organoids to model neurodegenerative diseases is their limited maturity. While strategies such as prolonged cultivation (>8 months) to accelerate the maturation and aging process in human cortical organoids have been validated through the analysis of key developmental milestones, including modifications in histone deacetylase complex and NMDA receptor signaling, such approaches are not without other significant challenges and complications, and are particularly sensitive to the aforementioned issues of necrosis. 104 Vascularizing organoids is an intriguing method to both accelerate the maturation of neuroglial components and improve long-term cell survival. One study has shown that the overexpression of ETS variant 2 (ETV2) in hESCs resulted in their differentiation to the endothelial phenotype and formed vascular-like structures in cultures. Coupling ETV2-hESCs with untransduced hESCs in a 20:80 ratio led to the formation of vascularized organoids, as cells organized into a physiologically relevant manner and created the beginnings of the blood brain barrier (BBB). 105 While still subject to optimization, these approaches to improving organoid maturation hold extensive value in allowing organoids to be used for modeling aging and neurodegenerative diseases.
Excitingly, recent advances in human CNS organoid technology have led to breakthrough findings in the field. For the first time, brain organoids with choroid plexus-like structures were developed with active human cerebrospinal fluid production. 64 Neuromuscular organoids developed in 2020 have successfully generated self-organizing populations of neurons and skeletal muscle and have demonstrated muscle contraction and neural activity, allowing for the capture of key features of myasthenia gravis and potentially providing the foundation for the study of other neuromuscular diseases. 106 Human brain organoids may potentially be utilized for the study of consciousness, as pioneering investigations have shown that network oscillation recordings done in dorsal forebrain organoids resemble electroencephalography patterns seen in preterm babies.107,108 Ultimately, CNS organoids have demonstrated their potential as a highly flexible system to be used in a wide array of study designs and applications and will continue to prove their tremendous value for disease modeling and cell therapies.
CNS Assembloids: Assembled and integrated organoids with improved cellular complexity and network connections
Although the field has made great strides with the use of neurosphere and CNS organoid technologies, a more complex model which captures intricate cell-cell interactions and circuitry connections is needed for in-depth study of disease. The rise of assembloid technology has made such requests possible. Assembloids can be described as assembled organoids or other 3D cellular structures with distinct regional identities and multiple cell types, which can be derived from different lineage origins. CNS assembloids usually involve the integration of various hPSC-derived cellular components that add a level of complexity to human mini brain models. They merge different CNS-relevant cell types, fuse region-specific CNS organoids, or create models that blend both CNS cells and organoids, most notably through the addition of non-neuroectodermal cells, such as microglia or endothelial cells. 109 In turn, these models provide insight into the overall spatial communication and network connectivity that occurs within the human brain and the rest of the nervous system. 45 In addition, analysis of the anatomical and functional components of assembloids in both normal and disease models can give a comprehensive understanding of scenarios in which CNS development goes awry as well as inspire novel treatment approaches.76,110
Assembloid technology has been used to create models of bodily systems that involve close interaction with the CNS. Such strategies were initially applied to generate the human visual system in vitro. Human visual system development involves the coordination of a series of spatial and temporal organizational events, including the elongation of axons from retinal ganglion cells (RGCs) to their postsynaptic targets in the brain. 111 In the human body, most RGCs directly project their axons into the thalamus in the brain. Thus, to study this interface within the visual system, hPSC-derived retinal organoids were engineered and organized into an assembloid model with cortical and thalamic organoids derived from the same hPSCs. 112 In a manner similar to behavior of RGC axons observed during visual system development, RGCs within retinal organoids responded to environmental cues by growing longer and extending deeper into neighboring thalamic organoids. More interestingly, astrocytes originating from the thalamic organoids were identified to retrogradely migrate into tdTomato-labeled retinal organoids in a manner that reflected the in vivo cell migration during the development of the human visual system. 112 Additionally, GFP positive axons from the thalamic organoids were seen to project into the cortical organoids to complete the retinal-thalamic-cortical assembloid structure. 112 This early proof-of-concept study showed enormous promise in applying human CNS assembloids to systematic studies of cell-cell and region-region interactions during human brain development. In addition to their applications in studying human brain development, human CNS assembloids have also been used to investigate neurological diseases caused by defective neural circuits. A complex assembloid model was developed to probe Phelan-McDermid syndrome, a disorder related to cortico-striatal deficits and characterized by global developmental delay and severe intellectual impairment.66,113 To accomplish this, lateral ganglionic eminence (LGE) and cortical organoids were generated from hPSCs. Subsequently, LGE organoids were derived into striatal organoids by introduction of activin A, IWP-2, and SR11237 molecules. These striatal organoids were then combined with cortical organoids to form cortico-striatal assembloids. Neurons arising in the cortical organoids formed synapses with those in striatal organoids, a finding consistent with the behavior of cortico-striatal circuits studied in vivo.66,114 However, cortico-striatal assembloids derived from patients with Phelan-McDermid syndrome showed deficits in neural circuit formation, as well as improper calcium signaling.66,114 To date, several assembloid models have been developed for investigating CNS diseases (Table 1). These models give researchers the unique ability to explore interneuron migration and axon projection, two main phenotypic defects seen in both neurodevelopmental and neuropsychiatric disorders.59,66,76,110,111,114
Recent efforts have been made to study long-range axonal connectivity by “upgrading” assembloids into more advanced structures, often named circuitoids, which are generated through the integration of modular assembloids. One breakthrough in circuitoid technology was the generation of cortico-spinal-muscle circuitry for analysis of the motor pathway. 76 Hindbrain/cervical spinal cord organoids (hSpOs) were initially fused with human forebrain cortical organoids (hFCOs) to create cortico-spinal assembloids. Voltage clamp recordings of hFCO-hSpO assembloid slices showed the establishment of synaptic connections between the two regions. Using an extracellular matrix-coated silicone well, hFCO-hSpO assembloids were coupled with human skeletal myoblasts (hSkMs) to create hFCO-hSpO-hSkM circuitoids. These circuitoids in turn established interconnectivity resulting in the modulation of muscle activity (Figure 4). 76 Previously, such studies could only be performed on motor circuit components that were generated separately in a 3D fashion or through co-culturing with 2D cells.41,44,115 –118 This approach depicted a significant advance beyond prior models by allowing 3D assembly of the motor circuit. Looking forward, this system and others like it might be used to develop an in vitro model of spinal cord injury and to evaluate approaches to promote regeneration. Furthermore, assembloids have been used in transplantation based studies to address glioblastoma, motor deficits, Alzheimer’s disease, and Parkinson’s disease (Table 1).68,70,119 The results of these studies highlighted that assembloids could integrate with host physiology to modulate muscle contractions, accept native vasculature, and recapitulate tumor-like structures.68,70,119
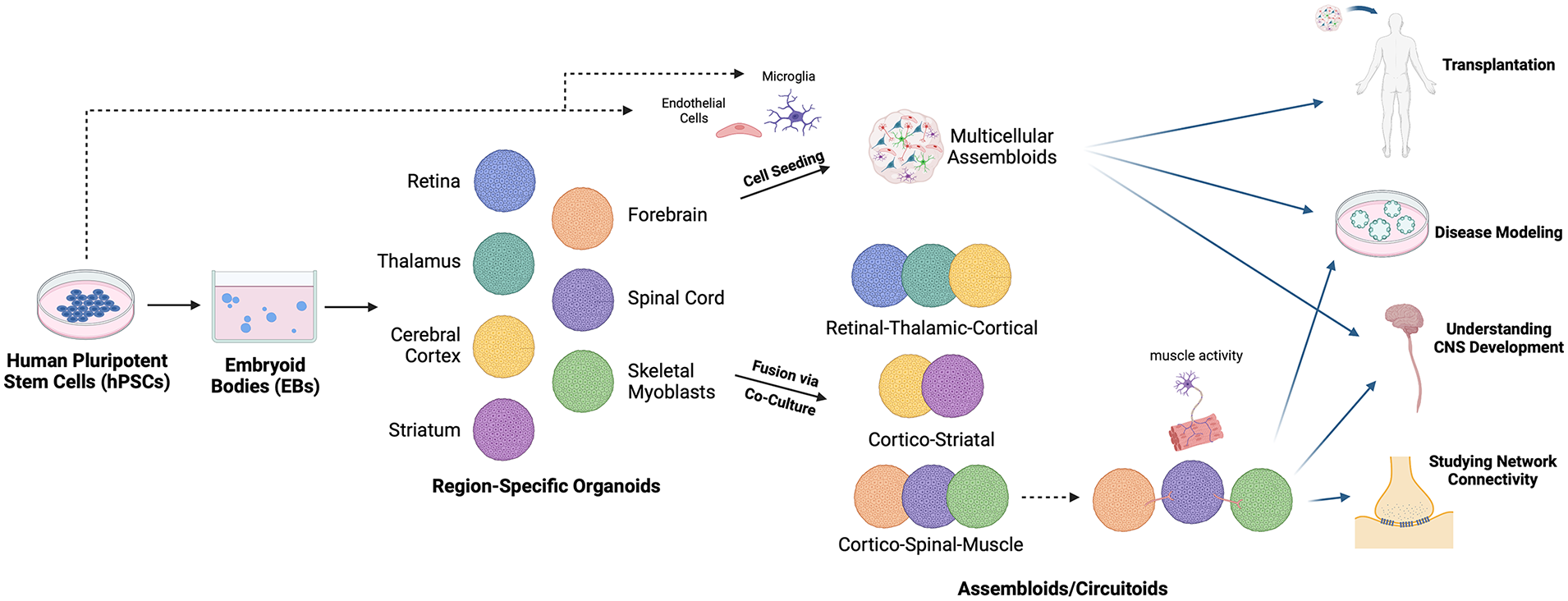
CNS Assembloids allow for the study of region-region circuitry. hiPSCs can be aggregated into embryoid bodies, which can then be differentiated into CNS organoids representative of different brain and spinal cord regions. Organoids can then be co-cultured with other hiPSC-derived cell types to form multicellular assembloids. Organoids with various region identities can also be fused through co-culture to produce assembloids modeling region-region connections. Assembloids can be utilized to study region-region circuitry when modeled as circuitoids. These structures can be used for transplantation studies, disease modeling, understanding CNS development, and studying network connectivity.
Although currently developed assembloids come close to imitating the human CNS, most models still do not accurately represent its complexity. Presently, most engineered assembloids combine only a few region-specific organoids with very restricted cell types.84,120 Ideally, a complete assembloid would be a single coherent and interconnected structure that requires the differentiation and integration of organoids with more complete region identities. The incorporation of all relevant cell types, including myelin-forming oligodendrocytes, microglia, ependymal cells, etc., could allow researchers to better understand the cell-cell interactions that are implicated in CNS development and disease.5,121,122 In models where multiple cell lineages are involved, methods to promote visualization of cell movement and functionality are essential. Thus, the development of cell type-specific reporters for the live-imaging of these cells has become increasingly important.5,122 These limitations leave room for improvement in expanding the assembloid model to capture the entire scope of the human brain. Fortunately, as seen by recent advances in the field, scientists are making every effort to optimize assembloid technology. In order to mimic the precise organization of the CNS, some have succeeded in incorporating distinct signaling centers within their assembloid models.123 –125 These signaling centers polarize the assembloids by engineering the cells to secrete (SHH) protein for establishing an SHH gradient. 123 Such methods introduce a sense of spatial organization to the overall system by taking the assembloid components that are initially combined at random and coordinating them by providing a positional identity. In order to keep each of these components alive and growing, some groups have also begun improving the scalability of the assembloid model. Although several researchers have successfully maintained assembloid cultures for several months after assembly, limitations in long-term cell viability make it difficult to conduct studies at more mature stages. 5 The automatization of cell culture maintenance is starting to overcome this problem by allowing for the exploration of CNS diseases that occur later in life or those that involve long-term degeneration. 126 Ultimately, assembloid technology has provided an in vitro or ex vivo alternative for the use of animal models in human CNS disease modeling and has already showed significant improvement from initial 3D cell culture methods.
Organ-on-a-chip: Utilizing microfluidics for CNS modeling
Key engineering-based modalities, namely the use of microfluidic chips and organ-on-a-chip (OoC) systems, have the potential to bring 3D mini brain cellular structures to life. In the context of building the “next generation human mini brain and spinal cord platform,” microfluidic chips are typically polydimethylsiloxane-based (PDMS) systems designed for the purpose of tuning critical physiological parameters including fluid flow, cell-cell mechanical interaction, and cell to cell biochemical signaling. 127 PDMS chips are low in material cost and highly accessible, enabling a wide range of experiments to be performed and the creation of more physiologically-relevant models. 128
One of the main drawbacks with static culturing methods is their inability to provide sufficient air and nutrient exchange for the robust metabolism of growing 3D cellular structures. While orbital shakers and spinning bioreactors are common solutions for introducing fluid flow in cultured 3D CNS structures, they do not provide a controlled method for regulating the equilibrium between fresh and spent medium, a factor that may influence the real time health of the mini brain. Microfluidic chambers, specifically the use of micropumps, provide the ability to control flow, allowing fresh medium to be pulsed into the 3D microenvironment in a manner much more resemblant to the human vascular system. While this micropump technology has demonstrated its ability to influence and couple with cardiac microtissue, OoC systems on the other hand have been utilized to introduce vasculature into CNS models. 129 One study utilizing OoC systems looked at the effects of brain microvascular endothelial cells (BMEC) on neurogenesis, specifically by leveraging small medium volumes and dual-channel constructs to create vascularized CNS constructs. 130 Another study used chips housing porous polycarbonate membranes to separate two channels filled with endothelial cells and astrocytes, and effectively recreated the permeability and sheer stress properties seen at the BBB. 131 Integrating micropump technology with vascular network forming endothelial cells may one day enable nutrients to be pulsed through vascularized 3D mini brains and spinal cords, an approach that may finally address the issue of progressive necrosis in long-term cultured 3D CNS structures (Figure 5). Other studies have taken different OoC approaches to improve the nutrient accessibility and reproducibility of in vitro 3D culture. One proof of concept study used tubular OoC devices to generate organoids on hollow mesh scaffolds and found improved nutrient diffusion, decreased sample heterogeneity and increased neural progenitor populations. 132 Another OoC study used an open-chamber design to optimize the placement and seeding of a cerebral organoid in the center of a chamber, enabling easy access for downstream analyses and more controllability through single organoid studies. 133 The high customizability of OoC systems offers solutions to improve organoid health and long-term survivability, two of the major limitations implicated in 3D CNS culture.
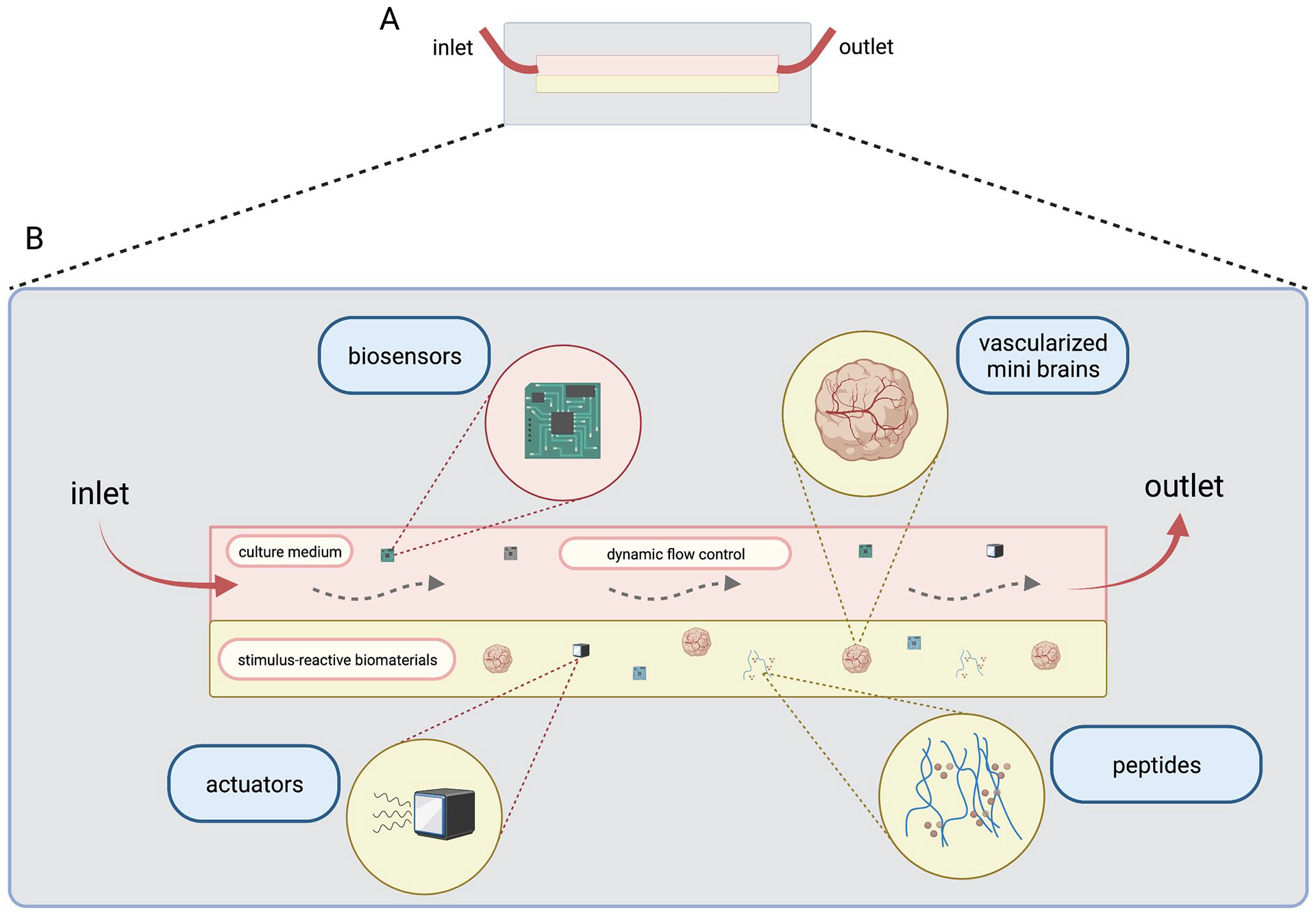
Customizable microfluidic chips provide mini brain models with a variety of physiologically critical stimuli. (A) Side view of microfluidic chip system with inlet and outlet points. (B) Enlarged side view of microfluidic 3D environment with detailed manipulations. The ability to control the flow of culture medium as well as continuously remove spent medium and toxins allows for better sample survivability and maturation. Engineered stimulus-reactive biomaterials enhanced with peptides may be designed for specific biochemical signaling purposes and may be coupled with other microfluidic components to create more physiologically relevant extracellular environments. Under flow conditions, vascularized mini brains may be able to recreate the intricate circulatory system responsible for the nourishment of the brain. A variety of biosensors may be integrated into microfluidic systems to provide real time feedback of culture conditions and can be coupled with actuators that can make real-time changes to critical parameters.
Microfluidic chips also provide unique opportunities for understanding how mechanical forces influence human brain development and activity when combined with mini brain technologies. By controlling the geometry and confines of the culture environment, one can design experiments analyzing the mechanical forces influencing cortical folding during neurodevelopment, which have been implicated in several neurological disorders. 134 One study used an OoC system coupled with a hydrogel filling to culture human brain organoids and study compressive forces on nuclei and how they influence cortical wrinkling. 135 Furthermore, the adaptability of microfluidic technology allows for microsensors to be embedded into OoC systems, providing revolutionary access to the culture environment. Several studies have demonstrated the integration of micro-electrode arrays into OoC systems as a means for measuring the real-time electrical activity of neuronal samples.136,137 Systems integrating sensors measuring mechanical forces, nucleotide variations, and small molecule release have also been reported.138 –140 In turn, one can begin to realize the utility for a microchip with dynamic force control, particularly in the context of studying traumatic brain injury. Approaches to modeling traumatic brain injury in human mini brain models often include needle-stick injury, sheer injury via a moving plate, or even high intensity focused ultrasound (HIFU) induced mechanical injury, all of which have their own caveats.86,141 Microfluidic chip technology may be the solution that laboratories need to expand such study of traumatic brain injury, especially given the multidimensionality of the injury and how it relates to other neurodegenerative disorders such as Alzheimer’s disease and frontotemporal dementia. Ultimately, OoC systems and microfluidic technology open the door to new approaches in uncovering the environmental factors implicated in both normal and neuropathological processes. Despite their fixed constructs and currently limited transplantation potential, their full potential in the field of 3D CNS modeling is yet to be realized.
Biomaterials: Key supporting elements to in vitro 3D CNS environment
The advancement of biomaterial technology has proven to be monumental in improving the feasibility of human mini brain and spinal cord models to be used as reliable in vitro and ex vivo models. Spearheaded by Matrigel, a commercially available mouse-derived mixture rich in ECM proteins, many types of hydrogels provide the ability to tune specific mechanical and biochemical properties of the extracellular matrix, which is important for modulating behaviors like neural process development, synapse formation, and cell migration. 142 While culturing neurospheres, organoids, and assembloids embedded in Matrigel has been and continues to be a popular approach, there are several drawbacks to Matrigel, particularly its animal origin and relative heterogeneity, that limit it from being deeply translational in the study of 3D CNS structures. Synthetic hydrogels on the other hand, can be consistently reproduced and provide a base for manipulating parameters relevant to the application they are involved in. Dynamic covalent chemistry (DCC), which relies on the reversible formation and breakage of covalent bonds, has emerged as a powerful tool for controlling the crosslinking and subsequent viscoelastic properties of a hydrogel. The potential application of DCC in in vitro modeling technology includes precise coordination of cell differentiation, migration, and maturation within the 3D environment.143,144 One study utilizing DCC to link DNA strands to polyacrylamide hydrogels showed that hydrogel stiffness contributes to neural sprouting. 119 Given that the mechanical properties of the brain change throughout development, hydrogels may be utilized to promote more complex organoid and assembloid spatial organization, as the stiffness and stress-relaxation characteristics of hydrogels housing 3D mini brains can be tuned in real-time to match developmental parameters seen in the developing brain. 145 For example, one study used photo-responsive crosslinkers to temporally and spatially tune the stiffness of a hydrogel, in turn demonstrating the ability to optically control cell migration. 146 Precision control of local hydrogel properties may be useful in directing the migration of microglia or endothelial cells, which can be seeded into assembloid-containing hydrogel scaffolds to replicate the infiltration of those non-neuronal cells into the CNS during embryonic development. 147 Moreover, hyaluronic acid (HA) incorporated hydrogels have been shown to improve the stability of hydrogels in 3D cultures. It should be noted that HA hydrogels are particularly important in neural based applications given the high HA composition in the human brain ECM, as it presents a great promise in their application for advancing mini brain and spinal cord technology.75,148 One study investigating how migration is affected in certain neurodevelopmental disorders, such as Rett Syndrome, showed that methacrylate modified HA hydrogels improved neural migration in neurosphere cultures. 149 Another study identified the utility of HA in promoting axon regeneration and angiogenesis, as severed axons at the site of spinal cord injury in mice were found to extend into transplanted HA-composed hydrogels. 150 HA hydrogels containing stem-cell derived axon bundles are already being tested for clinical applications to restore damaged pathways in patients with Parkinson’s disease. 151 Expanding this type of approach to include semi-mature organoid structures may be able to address a wider range of conditions with larger implicated injury sites. In addition, the covalent integration of peptides and other signaling molecules into these “designer hydrogels” opens a new avenue for manipulating culture conditions and further exploring brain properties (Figure 5). The peptide Arg-Gly-Asp (RGD), for example, is commonly used to increase cell adhesion, a property important for cell migration and injury recovery. 152 One study utilized polyethylene glycol (PEG) hydrogels supplemented with RGD-derivative peptides to produce 3D neural constructs of neural, mesenchymal, and microglial cells, and showed this approach was able to consistently reproduce CNS constructs of similar RNA-seq profiles. 153 Similarly, amphipathic β-sheet forming peptides such as EAK16, RADA16, and IKVAV have been developed to improve mammalian cell adhesion while also remaining incredibly stable over a variety of temperatures, pHs, or enzymatic ranges. 154 One study showed that the laminin-mimetic IKVAV sequence was able to promote bone marrow-derived mesenchymal stem cell differentiation into the neuroectodermal lineage after just one week, a finding particularly important in improving the generation and throughput of neurosphere cultures. 155 These biodegradable scaffolds go beyond improving the in vitro culture systems and have already shown potential in treating Alzheimer’s disease and Parkinson’s disease from a neuro-regenerative approach. Given their synthetic origin, peptide-modified scaffolds are great candidates for organoid-transplantation solutions and future work hopes to explore their utility in these approaches. 156 Ultimately, the continued integration of innovative peptides with hydrogels into 3D CNS cultures will enhance both in vitro modeling capabilities as well as inspire novel therapeutic solutions for neurodegeneration and CNS injury.
The real potential of biomaterials begins to reveal itself in “smart hydrogels,” which can perhaps integrate nanotechnology to increase their real time influence on 3D culture. The integration of biochemical sensors and actuators into these hydrogels may allow them to change their properties in response to culture conditions in real time (Figure 5). Stimulus-reactive hydrogels have been implemented in a wide variety of biomedical applications and have so far been designed to use temperature, pH, compressive, light, electric field, magnetic, ultrasound, antigenic, ionic, and enzymatic stimuli to induce desired behaviors in certain systems. 157 While their utility in 3D CNS culture has yet to be fully realized, they hold significant promise in improving culture conditions by providing researchers with a wider range of levers to pull in the customization of their models. For example, several studies have captured the influence of electric fields on nervous tissue development, particularly in terms of cell differentiation, proliferation, and axon growth.158,159 In turn, studies utilizing conductive nanomaterials, such as polypyrrole, polyaniline, and polythiophene, have demonstrated their ability to respond to electrical stimulation in the form of direct, biphasic, alternating, and pulse currents to modulate local cell migration, gene expression, maturation, and process extension of neural progenitors and mature neurons.158,160 –162 Precise electrical stimulation for enhancing the development and maturation of a 3D CNS model may decrease the need to add large cocktails of bioactive compounds and growth factors, which often have off-target effects that confound study results. The design and implementation of smart hydrogels may in turn provide us with a level of control previously unimaginable in engineering complex but orderly 3D mini brains and spinal cords. This level of precision is exactly what the human mini brain and spinal cord field needs to prove translational in the study of human neurodevelopment and many kinds of neurodegenerative disorders.
Conclusion and perspective
In vitro and ex vivo human 3D CNS models have great potential in meeting Neuroscience’s growing need for more advanced CNS modeling platforms. Neurospheres, CNS Organoids, CNS assembloids, OoC systems, and their combinations are several modeling strategies that allow for great flexibility and ingenuity in mechanistic study for CNS development and disorders, as well as fostering translational solutions to treat neurological disorders. Despite the great promise of contemporary engineered human 3D CNS technology, there are still issues in its variability, maturity, and reproducibility that need to be overcome before its large-scale application in translational biomedical research (see their pros and cons in Table 2). Efforts to reduce the variability of generating these 3D structures, especially for organoids and assembloids, involve defining the type and number of the originating cells, standardizing the reagents and procedures of the development process, and adjusting the timing for the patterning and maturation strategy. In particular, reduced variability has been achieved through using more committed cell sources and optimized time-windows for patterning.163,164 Ideally, automated systems with liquid and plate handling should also relieve some of the concerns in culture variability by standardizing medium changes, embryoid body formation, and nutrient distribution in culture. Well-defined and standardized biocompatible materials combined with 3D printing technology have shown to improve the reproducibility of using organoids and assembloids as disease models.165–167 Strikingly, integration of endothelium in these engineered 3D CNS tissues not only escalated the complexity of these structures, but advanced their maturity (our unpublished data).168,169 Innovative prolonged culture protocols, as well as those to accelerate the cell growth and differentiation also showed promise in promoting maturation.62,170 Slice cultures derived from organoids provide an air-liquid interface allowing improved oxygen penetration into the core structure, which also facilitates neuronal differentiation, axonal growth and synaptic formation.171,172 Tubular organoids and other OoC systems have also been able to address some of the shortcomings of 3D in vitro culture. 132 Additionally, supporting materials, including bioreactors, microfluidic chips, biomaterials, etc. have proven to be critical in optimizing these methods for laboratory and clinical use. Current research highlights using human mini brains and spinal cords to underline the rapid development of 3D human CNS models in the bioengineering and neuroscience fields and aspires to stimulate further innovations and advances in overcoming some remaining hurdles that limit their applications.
Summary of strengths and weaknesses of various 3D CNS modeling strategies including neurospheres, CNS organoids, CNS assembloids, and Organ-on-a-chip.

Footnotes
Author contributions
LQ and SK initiated the work and designed the idea. All authors contributed and revised the final version of the manuscript.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The work on using hiPSC derived models in the authors’ laboratory was supported by NIH grant R01NS115977 awarded to LQ; the Lisa Dean Moseley Foundation grant awarded to LQ; Spastic Paraplegia Foundation grant awarded to LQ, and the grant from the CURE program via Drexel University College of Medicine to LQ (CURE Grant SAP Number: 4100083087).