
Abstract
Liver failure is a lethal condition with hepatocellular dysfunction, and liver transplantation is presently the only effective treatment. However, due to the limited availability of donors and the potential immune rejection, novel therapeutic strategies are actively sought to restore the normal hepatic architectures and functions, especially for livers with inherited metabolic dysfunctions or chronic diseases. Although the conventional cell therapy has shown promising results, the direct infusion of hepatocytes is hampered by limited hepatocyte sources, poor cell viability, and engraftment. Hence, this review mainly highlights the role of stem cells and progenitors as the alternative cell source and summarizes the potential approaches based on tissue engineering to improve the delivery efficiency of cells. Particularly, the underlying mechanisms for cell therapy using stem cells and progenitors are discussed in two main aspects: paracrine effect and cell differentiation. Moreover, tissue-engineering approaches using cell aggregates and decellularized liver scaffolds for bioengineering of functional hepatic constructs are discussed and compared in terms of the potential to replicate liver physiological structures. In the end, a potentially effective strategy combining the premium advantages of stem cell aggregates and decellularized liver scaffolds is proposed as the future direction of liver tissue engineering and regeneration.
Introduction
As one of the most important organs, liver participates in various physiological activities, including the production of bile and albumins, metabolism of toxins and drugs, and maintenance of glucose and lipid balance. 1 Therefore, a severe hepatic injury can cause serious consequences despite the great regenerative capacity of a liver, particularly for patients who have inherited metabolic disorders or chronic liver diseases such as liver cirrhosis. 2 Basically, transplantation of a living-donor-liver is a gold standard for treatment of severe liver failure. However, the shortage of suitable donors constitutes a major obstacle for prompt treatment. 3 Meanwhile, immune rejection is another potential issue after liver transplantation, which requires long-term immunosuppressive management. 4
Bioartificial liver (BAL) as a temporary liver supporting device can provide essential hepatic functions for the patients who are waiting for the transplantation of living-donor livers. 5 Conventional BALs work by the extracorporeal circulation of patients’ blood through the devices, which have some limitations such as large and complex equipment. 6 Therefore, there are urgent needs to enhance the hepatic functions of BALs while simplifying and minimizing their overall complexity and size, respectively.
Cell therapy is a novel approach for the treatment of liver diseases by direct injection of functional cells. 3 Hepatocyte is the most important component of liver, which consists of ~60% of the total hepatic cell population and is responsible for the majority of liver functions. 1 To retain at least 30% of the minimal liver functions, approximately 8.4 × 1010 hepatocytes are required, which poses a huge challenge for the treatment of liver failure only using autogenous cells, due to their few numbers of normal hepatocytes in vivo and limited proliferation capacity in vitro, not to mention the need for other non-parenchymal functional cells (liver sinusoidal endothelial cells, Kupffer cells, hepatic stellate cells, and cholangiocytes). 7 Furthermore, low cell survival rate and poor cell engraftment are the common issues during infusion of single-cell suspensions in a conventional cell therapy.8–11 Therefore, seeking an alternative cell source and enhancing cell delivery efficiency are two potential solutions for the effective treatment of liver failure. 12
Several excellent reviews describing biomaterials and advanced techniques, such as 3D printing and microfluidics, for liver disease modeling and treatment are already available.13–16 For example, Morais et al. 13 systematically summarized a wide range of natural and synthetic biomaterials in various forms such as hydrogels or solid scaffolds for liver regeneration. Meanwhile, different 3D-printing strategies for liver tissue engineering and regeneration have also been discussed in the review. 13 Herein, in this review, we mainly focus on the stem cell therapy and tissue engineering strategies using cell aggregates and decellularized scaffolds for liver tissue engineering and regeneration (Figure 1). The role of stem cells and progenitors in the rescue of liver failure is revealed. The up-to-date cell aggregate-based (Table 1) and/or decellularized liver scaffold-based approaches (Table 2) to enhance the delivery efficiency of cells and bioengineer functional liver constructs are also discussed. Finally, we propose perspectives on research directions for effective treatment of severe liver failure based on liver tissue engineering and regeneration.

A schematic illustrates the use of stem cell therapy, cell aggregate-based or decellularized liver scaffold-based tissue engineering strategies for the rescue of liver failure. To reconstruct a normal liver, stem cells/differentiated cells/bioengineered cell aggregates can be implanted into the abnormal liver or its decellularized counterpart.
Bioengineering of functional hepatic constructs based on cell aggregates.

Bioengineering of functional hepatic constructs based on decellularized liver scaffolds.

Stem cell therapy
Stem cells and progenitors can not only self-renew through cell proliferation but also differentiate into terminal functional cells.1,35 Theoretically, pluripotent stem cells, embryonic stem cells (ESCs) and induced pluripotent stem cells (iPSCs), are able to replicate indefinitely and differentiate into any cell types derived from the three germ layers, while mesenchymal stem/stromal cells (MSCs) are only capable of multipotent differentiation and progenitor cells are more committed to their tissue of origin such as the bi-lineage differentiation potential of hepatoblasts into mature hepatocytes and cholangiocytes.1,3 Typically, there are two major roles of stem cells and progenitors in liver regeneration, including paracrine effect and hepatic differentiation. The former targets host cells for activation and acceleration of self-healing, while the latter produces new hepatic tissues to replace the injured liver. 36
Paracrine effect
The secreted growth factors involved in paracrine effects affect numerous intra- and extra-cellular signaling pathways, which may trigger a series of reactions promoting liver regeneration.36–40 For example, small hepatocyte-like progenitor cell (SHPC) clusters, one type of cells that have some phenotypes similar to hepatocytes, that appeared in injured rat livers induced by administration of retrorsine (Ret) and 70% partial hepatectomy (PH) increased in number and size after implantation of Thy1+ cells, one type of hepatic progenitor cells. 41 The liver regeneration was enhanced through the IL17RB signaling pathway regulated by Thy1+ cell-released extracellular vesicles (EVs). Yu et al. 42 showed that hypoxia-preconditioning of bone marrow-derived mesenchymal stem/stromal cells (BMSCs) promoted liver regeneration after the infusion of cells into the portal vein of a rat model with 85% hepatectomy due to their enhanced expression of vascular endothelial growth factor (VEGF). Liver regeneration was also enhanced in a partially hepatectomized mouse model following infusion of conditioned medium of liver-derived MSCs through promoting cell proliferation and reducing proinflammation. 43 Such therapeutic efficacy could be further promoted by using conditioned medium from adipose-derived mesenchymal/stromal cells (ASCs) in a hypoxic condition (1% oxygen) via the JAK/STAT3 signaling pathway. 44 In another study by Lee et al., 45 liver regeneration in the partially hepatectomized mice was observed after intravenous administration of the exosomes derived from lipopolysaccharide-preconditioned ASCs. The superior therapeutic efficacy of MSCs and their exosomes for early hepatic ischemia-reperfusion injury (IRI) was reported by Anger et al. 46 In addition, the paracrine effect of endothelial progenitor cells (EPCs), particularly those derived from bone marrow, has also been evidenced with enhanced liver regeneration and suppression of liver disorders after their extensive proliferation and engraftment into the injured liver.22,47–50 The secreted paracrine factors such as VEGF and hepatocyte growth factor (HGF) from the EPCs could promote hepatocyte proliferation and angiogenesis of resident liver sinusoidal endothelial cells (LSECs) in addition to the induction of apoptosis in hepatic stellate cells (HSCs).48,49,51–53 However, it is notable that such paracrine effect can also aggravate the cirrhosis if circulating EPCs are derived from abnormal donors which might be due to the distinct functions of the two cell subpopulations, early EPCs and outgrowth EPCs that are involved in inflammation and angiogenesis, respectively.51,54–56
Growth factors are demonstrated to be the functional components of exosomes, therefore their targeted and sustained delivery may further promote liver regeneration.44,57,58 For example, Yu et al. 59 recently reported that cell proliferation of hepatocytes was markedly promoted after implantation of VEGF-loaded nanofibers into rats with 70% hepatectomy. Sustainable release of HGF, one type of paracrine growth factors, from carboxymethyl-hexanoyl chitosan (CHC) hydrogel was also reported to enhance cell proliferation of iPSC-derived hepatocyte-like cells and maintain their hepatic functions in vitro. 60 Meanwhile, the reduced area of hepatic necrosis and increased survival rate of thioacetamide (TAA)-induced liver injured mice were evidenced after the administration of HGF in vivo. 60 Moreover, multi-dose administration of insulin-like growth factor-1 (IGF-1), another exosome component, was also recently reported to reduce collagen deposition in mice by Fiore et al., 61 indicating the amelioration of liver fibrosis.
Cell differentiation
In addition to the activation and acceleration of self-healing process in the host liver by paracrine factors, stem cells and progenitors are also used for the treatment of liver failure based on their capability of differentiation toward hepatic cell lineages. 62 For instance, human induced pluripotent stem cells (hiPSCs) have been successfully induced to undergo hepatic differentiation in vitro to produce functional hepatocytes with comparable liver functions to those in vivo under flow conditions, suggesting that hiPSCs may be an accessible cell source for extensive liver tissue engineering.63,64 In addition to hepatocytes, pluripotent stem cells can also be differentiated into other nonparenchymal cells such as endothelial cells, 65 cholangiocytes,66,67 Kupffer cells, 68 and HSCs. 69 Although the low differentiation efficiency still requires further optimization of induction protocols, these successes make it possible for bioengineering of a whole liver organ using a wide range of differentiated cells with identical genomes. Of note, although hepatocytes and cholangiocytes are both derived from hepatoblasts during embryogenesis, recent studies have revealed the possibility of transdifferentiation of reactive biliary epithelial cells to hepatocytes in animal models with severe chronic liver injuries whose hepatocyte proliferation is inhibited.70,71 Unlike the dedifferentiation of mature hepatocytes and their robust expansion after liver injury,72,73 mature LSECs seem to do little favor to the liver regeneration. 49 Instead, bone marrow-derived EPCs were evidenced with enhanced proliferation, migration to the injured liver and differentiation into LSECs for the reconstruction of the hepatic vasculature.49,74,75 Similar results were also found for intrahepatic ECPs when they were isolated and reinfused into rats with hepatic injury.49,74
However, due to the high pluripotency of hiPSCs and ESCs, there is a potential risk of aberrant differentiation leading to teratoma formation. 76 One of the potential solutions for this issue is pre-differentiation of these cells toward multipotent stem cells or progenitors, which can be further induced to differentiate into specific cell types only. For instance, hESCs/hiPSCs were induced into MSCs by Spitzhorn et al. 77 before injection into the tip of the spleen of the 70% hepatectomized Gunn rats (a typical animal model for the study of Crigler-Najjar syndrome type 1). It was found that the implanted MSCs survived and engrafted into the host livers for up to 2 months with normal hepatic functions and no tumor formation, which might be because of cell fusion that led to the transfer of Ugt1a1 gene from human donor cells to rat host cells. Therefore, the hESCs/hiPSCs-derived MSCs could be a promising cell source for the repair of injured livers with heritable dysfunction.
Xu et al. 78 implanted ASCs-derived hepatocyte-like cells into the liver lobes of ganciclovir-administrated TK-NOG mice through ultrasound-guided multi-injection, resulting in favorable liver regeneration and no tumor formation for over 2 months. Interestingly, the authors also reported that iPSC-derived hepatocyte-like cells tended to induce visible tumors in 3 weeks after their implantation at the kidney capsules in vivo. However, in another study by Yuan et al., 79 no tumorigenesis was observed in the main host organs (liver, lung, kidney, heart, spleen, and colon) in 8 weeks after splenic implantation of iPSC-derived hepatocyte-like cells. The different results might be due to the distinct differentiation efficiency of iPSCs toward hepatocyte-like cells or the suppression of tumor growth by hiPSCs-derived MSCs, thus indicating the importance of an efficient pre-differentiation route in the clinical application of iPSCs.78,80
Another alternative cell source for liver regeneration, human bone marrow-derived muse cells, were directly injected into the tail vein of the immunodeficient mice for the repair of liver fibrosis by Iseki et al. 81 Interestingly, this type of pluripotent cells are capable of differentiation into cells within three germ layers but with no risk of tumorigenesis because of the low telomerase activity. The majority of the implanted muse cells showed rapidly homing to the damaged livers and underwent mature hepatic differentiation without fusion with host hepatocytes.
Bioengineering of functional hepatic constructs
Tissue engineering is an emerging field that has shown great potential in the fabrication of liver-mimicking structures for studying biological and disease development of liver, performing drug screening, and supporting hepatic functions and liver regeneration.14,82 There are a variety of studies already revealing the importance of nonparenchymal cells such as Kupffer cells, 83 hepatic stellate cells,84,85 and endothelial cells86,87 for liver regeneration. Thus, many co-culture systems have been established in vitro to mimic the specific liver structures. 88 For example, primary rat hepatocytes (PRHs) and liver sinusoidal endothelial cells (LSECs), freshly isolated cells from the rat liver, were cultured simultaneously on the opposite sides of the same collagen-coated Transwell inserts to imitate the layered organization of liver sinusoids, leading to the maintenance of normal cell morphology and viability as well as hepatic functions for around 40 days. 89 The bile duct-like networks were formed through a “sandwich culture” of bile duct epithelial cells in collagen hydrogels, which showed both functional and morphological similarity to the bile ducts in vivo. 90 Detzel et al. 91 constructed a 3D liver mimic comprised of multilayered primary rat hepatocytes and liver sinusoidal endothelial cells as an in-vitro platform for studying the metabolism of bile acid. The 3D liver mimic not only exhibited bile canaliculi but also showed similar physiological hepatic metabolism as that in vivo. Wu et al. 92 established a non-alcoholic fatty liver disease (NAFLD) model through perfusion culture of HepG2 cells in decellularized liver scaffolds, which provided a useful tool for studying the development of NAFLD in vitro and screening relevant drugs. Co-culture of hepatocytes and hepatic stellate cells as cell aggregates has also been recently reported by Coll et al. 69 and Mannaerts et al. 93 for modeling of liver fibrosis in vitro. Although various strategies have been reported for liver tissue engineering and regeneration, the most promising approaches are based on cell aggregates and decellularized liver scaffolds due to their replication of physiological liver structures.3,7,94 The premium benefits of cell aggregates and decellularized liver scaffolds are discussed in details in the following sections.
Cell aggregate-based approach
Cell aggregates are a group of cells binding with each other via cell proliferation and/or cell aggregation. 95 Previously published articles have different naming rules for this form of cells, which may cause confusion, thus this review uses cell aggregates as the representative of cell clusters, cell spheroids, cell pellets, microtissues, embryoids, and organoids regardless of their overall morphology. Cell aggregates are commonly seen in the formation of embryoids during embryogenesis, condensation of cells during skeletal development, and proliferation of hepatocytes during liver regeneration.84,96–98 There have been a variety of studies showing that cellular functions are enhanced in cell aggregates compared to single-cell suspensions, such as the enhanced immunomodulatory potential and multi-lineage differentiation of MSC aggregates and the increased production of albumin and urea in hepatocyte aggregates.22,99–106 Moreover, cell aggregates also show improved cell viability and survival rate after implantation in vivo.8–11 Cell aggregates can be generated by different methods,107–110 such as cell culture on low attachment plates or in hanging drops, as well as substrate-based and technology-assisted methods, amongst which 3D scaffold/hydrogel-based approaches are of high interest since cells can form cell aggregates in situ after seeding inside the 3D scaffolds/hydrogels.103,104,111,112 The approach does not require extra process, which can avoid cell loss or damage during cell harvest. For instance, Tong et al. 103 formed HepG2 cell aggregates inside glycyrrhizin-contained alginate hydrogels with enhanced cell viability, proliferation, and hepatic functions, suggesting the potential application of the injectable hydrogels in liver tissue engineering and regeneration. Spontaneous formation of cell aggregates in situ was also reported by Lau et al., 104 who embedded murine iPSCs within micro-cavitary alginate hydrogels, showing increased hepatic differentiation compared to monolayer culture. In another study by Kundu and Kundu, 112 HepG2 cell aggregates were formed within the porous fibroin-based scaffolds fabricated by lyophilization. The distinct cell proliferation in the different scaffolds might be correlated with their pore size and porosity since spatial confinement of pores could restrict the growth of cell aggregates. As the size of cell aggregates is highly related to cellular functions, the controllable properties of substrates are critical for their applications in liver tissue engineering and regeneration. 17
Bioengineering of conventional hepatocyte aggregates
Due to the advantages of cell aggregates, hepatocyte aggregates have been engineered and widely used in liver tissue engineering (bioartificial livers or implantable hepatic constructs) for the rescue of liver failure (Table 1). 16 For example, Saadi et al. 8 showed the formation of Huh7 hepatocyte aggregates inside fibrinogen-PEGDA microgels, which enhanced cell engraftment after injection of 15,000 cell-loaded microgels (6 × 106 cells in total) via the portal veins of the lateral and median liver lobes into female Lewis rats with 34% partial hepatectomy. In another case, Erro et al. 22 encapsulated HepG2 cells within alginate microgels and cultured them in a fluidized bed bioreactor functioning as the liver-assist device. The authors demonstrated high throughput of functional hepatocytes (~34-fold increase) without compromise of cell viability, which could be even scaled up to 0.7–1.0 × 1011 hepatocytes, accounting for approximately 30%–50% of normal adult liver mass after optimizing the provision of nutrients and oxygen. Furthermore, the expanded cells were able to be transported at room temperature for 48 h based on the storage protocol established by the authors. A liver-assist device containing the alginate-encapsulated HepG2 cell aggregates have been successfully used to support liver functions in pigs with ischemic liver failure.113,114 Chen et al. 64 recently also reported rescue of acute liver failure in pig models using a multilayer BAL device that contains clinically relevant number of functional hiPSCs-derived hepatic aggregates. Chang et al. 18 co-cultured primary rat MSCs and AML12 cell line, mouse liver-derived immortalized hepatocytes, by encapsulating the cells inside collagen/alginate volvox spheres using a high voltage electrostatic field system (Figure 2(a)). The hepatic differentiation of MSCs was higher during dynamic cell culture compared to that using static culture. After direct injection of the cell-loaded volvox spheres into the SD rats with retrorsine-CCl4 induced liver injuries, restoration of liver functions and regeneration of new normal hepatic tissues were both observed (Figure 2(b)). Ng et al. 17 recently fabricated extracellular matrix (ECM) protein-conjugated porous poly(ethylene glycol) scaffolds and showed the formation of human iPSC-derived hepatic progenitor cell aggregates only inside the collagen I-conjugated scaffolds with mature hepatic functions similar to those of primary hepatocytes. The cell aggregates resulted in better tissue integration, vascularization, and albumin production after implantation of the bioengineered hepatic constructs into the capsule of the mouse caudate lobe.

Bioengineering of implantable hepatic aggregates: (a) the process for generation of microspheres and volvox spheres using mesenchymal stem/stromal cells (MSCs) and hepatocytes (AML12). Scale bar: 200 µm and (b) the aspartate aminotransferase (AST) and alanine aminotransferase (ALT) concentration indicating liver injury level in the normal rats (Normal), injured rats (Group A), injured rats with implantation of MSCs/AML12-encapsulated volvox spheres (Group B), injured rats with implantation of volvox spheres only (Group C), injured rats with implantation of MSCs-encapsulated volvox spheres (Group D), or injured rats with implantation of AML12-capsulated volvox spheres (Group E) on week 0, 4 and 6.
Bioengineering of vascularized hepatocyte aggregates
Owing to the importance of vasculatures for normal physiological liver functions, various strategies have been employed to guarantee the supply of oxygen and nutrients to hepatic cells.115,116 One strategy is to implant hepatocyte aggregates near the existing blood vessels in vivo to promote angiogenesis. 117 As an example, Yap et al. 9 implanted murine liver progenitor cells embedded in growth factor-reduced Matrigel into the vascularized chamber fitted around the superficial epigastric vascular pedicle at the groin of SCID mice. After 14 days, glandular organization of cells and vascularization were only seen in the chambers with implantation of cell aggregates but not single-cell suspensions.
The direct co-culture of hepatocytes and endothelial cells is another strategy for bioengineering of vascularized liver constructs.118,119 The increased hypoxia-inducible factors at the center of hepatocyte aggregates can function as the chemoattractant, which results in the invasion of endothelial cells and the formation of capillaries. 37 However, the fusion of these cell aggregates should be minimized to avoid the central necrosis of large aggregates and maintain the high surface-to-volume ratio for the effective exchange of nutrients and gases. To achieve these goals, Pang et al. 23 used fiber fragments as the spacers to separate vascularized hepatocyte aggregates from each other and improve the supply of nutrients and oxygen. In a follow-up study, they further fabricated a Nylon 12-based 3D perfusion system composed of 43 chambers to house hepatocyte/endothelial cell aggregates, which could be theoretically scaled up to a clinically-relevant size (500 cm3 in volume) (Figure 3). Both cell viability and hepatic functions were improved during the perfusion culture. 24 Moreover, they also seeded hepatocyte and endothelial cell hybrids inside the customized micro-scale scaffolds. The pores and intersecting hollow channels of the scaffolds allowed sufficient penetration of the medium throughout the scaffolds when packed in a bioreactor for perfusion culture, leading to high cell viability and enhanced hepatic functions. 120
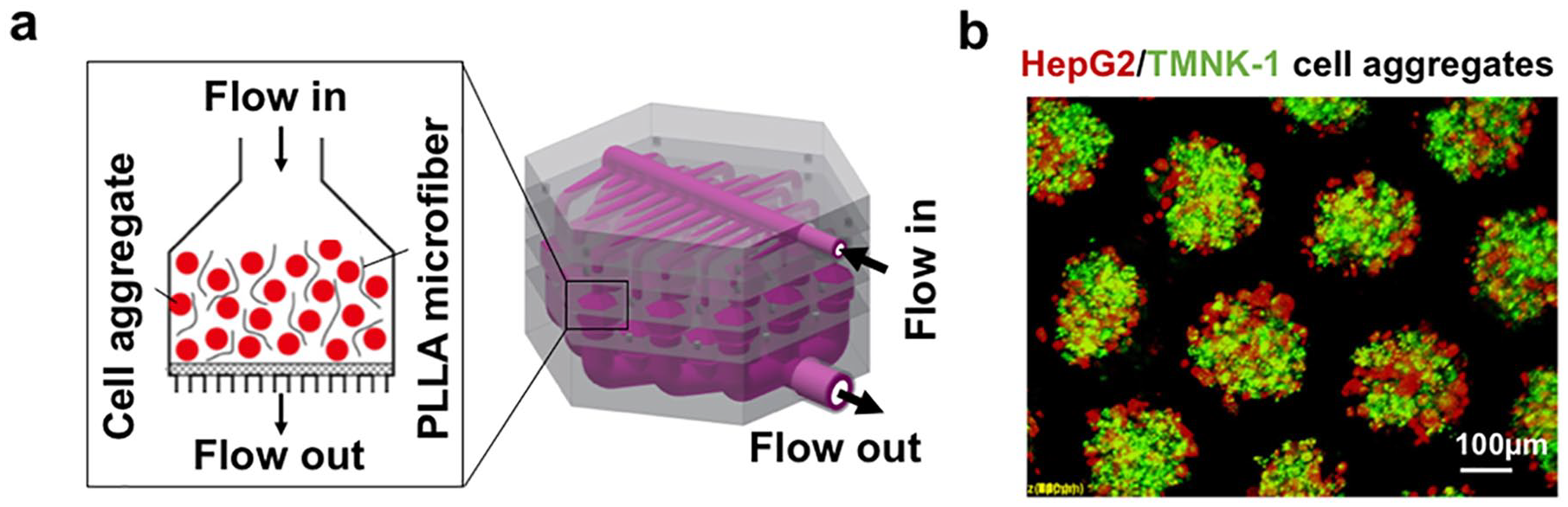
Bioengineering of a cell aggregate-based bioartificial liver: (a) dynamic culture of hepatocyte (HepG2)/endothelial cell (TMNK-1) aggregates in a customized 3D perfusion system and (b) Immunofluorescent staining of HepG2/TMNK-1 cell aggregates in the microwells 24 h after cell incubation.
In addition to fabrication of vascularized hepatocyte aggregates, advanced techniques for bioengineering of in-vivo liver-mimicking architectures have also been developed.19–21,25,121 Yajima et al. 19 recently fabricated a lobule-like hepatic structure through microfluidic device-assisted encapsulation of HepG2 cells within alginate hollow fibers and coculture of endothelial cells on the surface of the fiber bundles (Figure 4(a)). The cell proliferation and hepatic functions were significantly enhanced during perfusion culture. Liu et al. 21 proposed a new approach to construct the lobule-like tissue building blocks through the encapsulation of rat RLC-18 liver cells within the hexagonal poly-L-lysine (PLL)-alginate microcapsules using electrodeposition. Compared to cell spheroids, the same number of cells cultured inside the microcapsules tended to show increased hepatic functions, due to the hollow structure of the microcapsules that facilitated the efficient exchange of nutrients and oxygen. When these building blocks were assembled in a layer-by-layer manner, a 3D lobule-like liver construct with a central channel functioning as the vascular vessel was established (Figure 4(b)). In another study, Grigoryan et al. 25 fabricated a vascularized hepatic construct by seeding endothelial cells and hepatocyte aggregates inside the 3D-printed channels and the external carrying chamber, respectively (Figure 4(c)). Although the 3D bioengineered construct was not directly implanted into the host injured liver, but rather sutured at the perigonadal fat pad of mice, enhanced albumin production and host engraftment were observed in vivo. Du et al. 20 also assembled multiple polyelectrolyte fibers through multi-interfacial polyelectrolyte complexation (MIPC), which consisted of endothelial cells at the center and hepatocytes at the periphery to mimic the liver lobule structure (Figure 5(a)). As the hepatocytes and endothelial cells were both derived from the same iPSC source, they were genetically identical. In addition to the enhanced albumin secretion in vitro (Figure 5(b)), vascularization of the scaffolds and their integration into the host vasculature in vivo were evidenced after implantation of the lobule-like constructs into the livers of hepatectomized SCID mice (Figure 5(c)). Even though the in-vivo vasculatures were far more complex than the vascularized hepatic constructs fabricated in these studies, the positive results did indicate the importance of endothelial cells for a functional bioengineered hepatic construct. Hence, further works with an emphasis on bioengineering of complex vasculatures with physiological liver hemodynamics are required, which can be guided based on the design principles established by Hoganson et al. 122

Bioengineering of vascular hepatic constructs: (a) fabrication of lobule-like structures through encapsulation of hepatocytes within hydrogel fibers and seeding endothelial cells on the surface. Red: endothelial cell (EC); green: HepG2 cell; blue: cell nucleus. Scale bar: 50 µm. Adapted with permission from Yajima et al. 19 (b) fabrication of lobule-like structures through encapsulation of liver cells and multilayer assembly. Red: dead cell; green: live cell. Scale bar: 250 µm. Adapted with permission from Liu et al. 21 and (c) 3D printing of vascularized hepatic constructs by seeding endothelial cells and hepatocyte aggregates into the 3D-printed channels and the carry chamber, respectively. Red: endothelial cell (HUVEC); green: hepatocyte. Scale bar: 1 mm. Adapted with permission from Grigoryan et al. 25

Bioengineering of lobule-like structures: (a) the process for generation of vascularized hepatic constructs through multi-interfacial polyelectrolyte complexation (MIPC) with genetically identical endothelial cells and hepatocytes derived from induced pluripotent stem cells (iPSCs), (b) the albumin production of hepatocytes encapsulated in the fibers with or without endothelial cells in vitro. *p < 0.001, and (c) immunofluorescent staining images of hepatocytes and endothelial cells 4 or 6 weeks after implantation of the bioengineered hepatic constructs in vivo. Red: human albumin-positive hepatocyte; green: CD31 antibody-positive endothelial cell; blue: cell nucleus. Scale bar in i, ii, iii and iv: 100 µm. Scale bar in v and vi: 50 µm. Adapted with permission from Du et al. 20
It is worth noting that restoration of hepatic functions and liver regeneration in vivo are actually two different aspects during the assessment of the therapeutic strategies for specific liver diseases. For example, Sgroi et al. 123 showed that intraperitoneal implantation of alginate/PLLA/alginate-capsulated hepatocytes could support the normal hepatic functions in mice with acute liver failure (administration of acetaminophen and 30% hepatectomy), leading to improved animal survival. However, there was no significant difference in liver regeneration between mice with or without implantation of hepatocytes. Thus, to restore the normal hepatic architecture and functions, tissue engineering strategies for liver regeneration should position bioengineered hepatic constructs into the defect site of an injured liver instead of heterotopic implantation (subcutaneous or intraperitoneal accesses).
Decellularized liver scaffold-based approach
Decellularized liver tissues are ideal scaffolds for liver tissue engineering as the native liver architectures and compositions such as the pre-existing vascular network and scaffold-bound growth factors are retained (Figure 6).124–126 The intact vascular structures allow reconstruction of the complex hepatic vasculature in vivo for sufficient mass and gas supply while the bile tracts permit drainage of the excreted bile from hepatocytes in time. 7 In addition, the retained growth factors, such as VEGF, HGF, and bFGF, are essential biochemical cues for the maintenance of hepatic functions and regulation of stem cell fate.127–129 Therefore, decellularized liver tissues can be either broken into powders as the extracellular matrix (ECM) hydrogel formulation using the functional residues or used as a whole to take the advantage of the retained vasculature.7,130,131 For example, Sellaro et al. 127 revealed the maintenance of primary hepatocyte functions in porcine liver-derived ECM hydrogels comparative to that in the typical Matrigel™. Agarwal et al. 132 showed higher hepatic functions of HepG2 cells embedded within caprine liver-derived ECM 3D hydrogels compared with collagen hydrogels in vitro. Moreover, the ECM 3D hydrogels also supported the formation of microvasculature in vitro, which demonstrated its potential for bioengineering of a vascularized liver tissue construct for tissue engineering. Similar results were also shown in primary rat hepatocytes cultured in the rat liver-derived ECM 3D hydrogels. 133 Meanwhile, ECM can be used as the bio-ink for 3D printing due to its high versatility. 134 For instance, Lee et al. 128 showed that BMSCs and HepG2 cells embedded inside the ECM bio-ink showed enhanced hepatic differentiation and liver functions than that in collagen hydrogels, respectively. In principle, an artificial liver resembling the physiological hepatic architectures can be fabricated by 3D printing. However, technical challenges of 3D printing need to be first addressed, before the rapid engineering of liver organs at a high resolution can be realized.135,136

A schematic illustrates decellularization and recellularization processes for bioengineering of a functional liver construct.
There are a variety of feasible decellularization protocols using either chemical, biological or physical methods to remove cellular and genetic components from the fresh liver.130,131 Generally, chemical and biological decellularization methods can achieve effective decellularization but cause disruption of the ECM ultrastructure and loss of bioactive components, which are reversed by physical methods.130,131 Moreover, different sterilization methods have also been comprehensively studied. Of note, Hussein et al. 137 showed that sterilization of decellularized liver scaffolds using slightly acidic electrolyzed water could retain the most bioactive components such as glycosaminoglycan and collagen compared to ethanol and peracetic acid. To characterize the decellularized liver scaffold, various methods have been previous reported, including SEM imaging and histological analysis through H&E/IF/IHC staining, which are normally used for evaluation of structural maintenance and integrity as well as quantification of the residual DNA and the retained ECM biochemical compositions.130,131 By normalization of remained components to the initial weight of a whole organ unit, various decellularization protocols can be properly compared. 138 As the properties of decellularized liver scaffolds mainly rely on the preparation protocols, it is necessary to establish universal criteria for quality assessment. 131 In the study by Moulisová et al. 139 a three-level scoring system was used to evaluate the morphological structures of a given decellularized liver scaffold based on the measured parameters by the automatic customized software, ScaffAn. The authors claimed that the best discrimination between different decellularized liver scaffolds could be achieved by the multi-scale evaluation system. Despite the availability of diverse decellularization protocols, recellularization of whole liver organs currently still faces some challenges, amongst which poor cell distribution and thrombosis are the two most challenging obstacles in the clinical application of decellularized liver scaffolds. 29
Optimization of recellularization
The most common method utilized for recellularization is infusion of cells into the decellularized liver scaffolds via the existing vasculatures, therefore maintenance of the vascular and hepatic architecture during decellularization is the key for the subsequent cell repopulation. 7 There are a variety of factors that can influence cell distribution during recellularization, including cell density and morphology, infusion time, direction and access of infusion flow, and flow rate. Bao et al. 32 showed that infusion of single hepatocyte suspension through the portal vein of rat liver lobes at a cell density of 3.33 × 107 cells/mL and a flow rate of 2 mL/min resulted in faster cell death than that of cell aggregates (diameter: >30 μm) with the same amount of cells, which indicated the advantage of using cell aggregates in retaining cell viability. Meanwhile, the authors also claimed that infusion of hepatocyte aggregates for a longer time caused their integration into vascular spaces besides parenchymal regions. Therefore, the size of cell aggregates and infusion time still needed to be further optimized for a desirable cell distribution. Poor cell survival was also seen in a study by Uygun et al. 31 who infused primary hepatocytes into a whole decellularized liver organ via the portal vein in a four-step manner. Although cell viability was approximately 80% during the recellularization, the overall value was only 72% if the viability of initial seeding cells (~90%) was also considered.
In addition, antegrade infusion of endothelial cells via portal vein mainly resulted in the distribution of cells near the periportal areas, while retrograde infusion through vena cava led to the selective localization of endothelial cells at the larger and smaller vessels up to the pericentral regions of liver lobes but not yet reaching the periportal areas, which reflected the complex vasculatures in the decellularized liver tissues and how the direction of infusion flow influenced the re-endothelization. 140 Direct infusion of a mixture of human EpCAM+ fetal liver cells and endothelial cells via the vasculatures achieved good parenchymal recellularization, but the underlying mechanism was still not clear.140,141 Homing and differentiation of these highly migrant EpCAM+ cells through the attraction of specific growth factors bound on the decellularized liver scaffolds might be one of the explanations.140,141
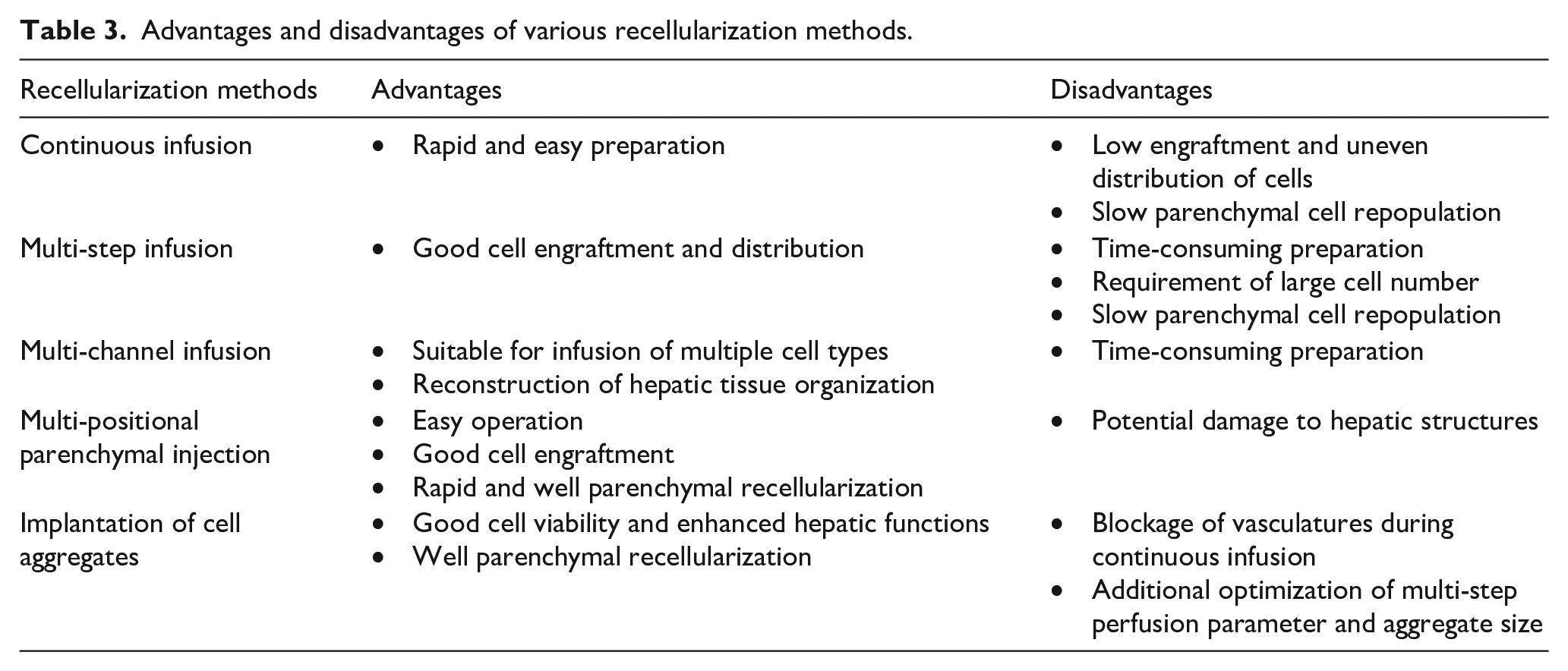
Although good recellularization can be obtained from cell infusion, cell migration-dependent repopulation of hepatocytes via vascular channels into parenchyma is not an efficient strategy for rapid restoration of liver functions, since it usually takes a few days. 31 Therefore, the optimization of cell seeding methods is still being pursued for recellularization. For example, endothelial cells and hepatocytes could be infused via different accesses to achieve a cell distribution resembling that of native liver. Infusion of endothelial cells via vasculatures and hepatocytes through bile ducts resulted in adequate reendothelization and well parenchymal recellularization, respectively.140,142 Alternatively, Zhou et al. 143 revealed that multi-positional parenchymal injection (10 sites) resulted in better cell engraftment and distribution than that of continuous cell fusion. In another similar study, Soto-Gutierrez et al. 144 reported that multistep infusion (86 ± 5%) of hepatocytes into the whole decellularized liver organ of rat via the portal vein exhibited the highest cell engraftment compared to direct multi-positional parenchymal injection (five sites, 12.6 ± 9%) and continuous infusion (69 ± 0.5%). All these results demonstrated that multiple cell infusions outperformed continuous infusion, and the efficiency of cell engraftment depended on the number of injections. 145 A comparison between various recellularization methods is summarized in Table 3.
Advantages and disadvantages of various recellularization methods.
The infusion rate is another factor that needs to be considered. A slow infusion rate always leads to cell precipitation, while a fast one may cause cell aggregation. 31 Hence, a suitable flow rate during recellularization is necessary along with additional contributing effects such as rotating/shaking, multistep infusion, or direct parenchymal multi-injections. 144 Baptista et al. 146 showed that hepatic cell organization and neovascularization in the decellularized liver tissues were in a flow rate-dependent manner. The results suggested the highest cell viability, proliferation, and occupancy within the decellularized liver tissues were achieved at an infusion rate of 9 mL/min, and it was regulated via nitric oxide (NO) pathway by flow-induced shear stress.
Elimination of blood coagulation
Although the vascular network is maintained during the decellularization of a whole liver organ, the exposure of the vascular basement membrane often triggers thrombosis upon blood reperfusion. Administration of heparin in the blood before the implantation of decellularized liver scaffold was attempted to avoid coagulation but failed after reperfusion for 1–2 h. 29 Immobilization of heparin to the vascular structures of the decellularized liver scaffolds was also carried out by Bruinsma et al. 147 through layer-by-layer assembly of positively charged poly(diallyldimethylammonium chloride) (PDADMAC) and negatively charged heparin. Anti-coagulation was demonstrated in a heparinization-dependent manner in vitro with no visible blood clots in the 8-layer heparin-coated scaffolds after 2-h perfusion of fresh diluted whole blood, but congestion occurred in 24 h during the in vivo assessment. Hence, further optimization of endothelialization might reduce the resistance of grafts and facilitate their successful transplantation in vivo. Hussein et al. 33 achieved efficient endothelialization of decellularized liver tissues through surface modification by heparin/gelatin after recellularization with hepatocytes, which showed enhanced hepatic functions and absence of thrombosis both in vitro and in vivo (Figure 7(a-i)). Although the authors monitored thrombosis in a pig model in vivo for 1 h only, the importance of endothelization for the successful liver repair was stressed. Heparin was also employed in a study by Bao et al. 32 to avoid thrombosis of the decellularized rat livers in vivo. Recellularization of hepatocytes in the scaffolds improved the hepatic functions and extended the mean lifespan of rats with 90% hepatectomy from 16 h to 72 h. In another study by Ko et al., 34 the authors showed uniform endothelial attachment throughout the liver blood vessels even for capillaries by conjugation of CD31 antibodies on the vasculature of the decellularized liver scaffolds (Figure 7(a-ii)). In addition, whole blood reperfusion in pigs both in vitro and in vivo displayed normal physiological blood flow up to 24 h (Figure 7(b) and (c)).

Bioengineering of whole liver organs: (a) the re-endothelialization process for decellularized liver scaffolds with vascular surface modification using either (i) heparin-gelatin or (ii) CD31 antibody. IH-IVC: intrahepatic inferior vena cava; PV: portal vein; RV: renal vein; RA: renal artery; HA: hepatic artery; SH-IVC: suprahepatic inferior vena cava, (b) Fluoroscopic angiography and immunofluorescent staining images of the unseeded decellularized liver (EC(-)) and re-endothelialized CD31 antibody-conjugated liver (EC(+)) one day after implantation in vivo. Red: platelet; green: endothelial cell (EC); blue: cell nucleus (Nu). Scale bar: 50 µm. Black arrows point to vasculatures indicating vascular patency. White arrows point to vascular vessel walls, and (c) the quantitative fluorescent intensity of platelets in EC(-) and EC(+) one day after implantation in vivo.
All these studies reveal the importance and challenge of recellularization, therefore a comprehensive study for the influence of all factors on the recellularization is essential for the clinical application of decellularized liver tissues.
Conclusion and perspectives
Living-donor-liver transplantation is the gold standard for the treatment of serious liver failure. However, alternative therapeutic strategies, including stem cell therapy and liver tissue engineering, are still needed because of the severe donor shortage.1,3,6,14 Although conventional cell therapy using direct injection of single-cell suspensions in vivo is a simple strategy for the treatment of liver diseases in a minimally-invasive manner, poor cell engraftment and survival often result in the requirement of multiple injections of high-dose cell suspensions to achieve expected therapeutic efficacy, which is not applicable for primary normal hepatocytes due to their limited number in vivo and challenging proliferation in vitro.8–11 Poor cell engraftment and low cell survival rate are also shown during the repopulation of cells in decellularized liver scaffolds, particularly for the recellularization of multiple cell types. 7 In addition, due to the difference of liver architectures across species, cell repopulation in decellularized xenograft did not show better results than that in homograft either in vitro or in vivo. 148
Stem cells and progenitors have shown promise in the repair of injured livers either through paracrine effects or hepatic differentiation, therefore they can be the alternative to primary hepatocytes in cell therapy.1,3 The paracrine effects modulate the local environment and accelerate the self-healing process of the injured livers, while the hepatic differentiation provides functional building blocks for the reconstruction of a normal liver. Although the synergy of paracrine effects and cell differentiation can maximize the therapeutic efficacy, treatment of inherited hepatic dysfunctions using stem cells or progenitors may mainly depend on cell differentiation since the host cells are characterized by genetic disorder. Before cell transplantation for the treatment of inherited metabolic liver diseases, the host liver is often pre-treated to increase cell engraftment in vivo using a variety of strategies, such as partial hepatectomy, portal ligature/embolization, or administration of chemotherapeutic drugs, which provide necessary stimuli for preparation of donor cell housing.149–152 For instance, Pourcher et al. 152 showed that cell engraftment was increased by around 3-folds after inducing volumetric portal embolization in the C57BL6 mice using polyester microspheres and subsequently injection of hepatocytes into the spleen.
Decellularized liver scaffold-based tissue engineering is a potential strategy for the treatment of liver failure due to their well-preserved hepatic architecture.124–126 In addition to recellularization, other concerns such as the potential risk of xenogeneic/allogeneic livers, long-term stability of liver grafts in vivo, and bioengineering of a physiologically relevant sized liver construct should also be addressed before clinical application of the decellularized liver scaffold-based approach.7,33,34 Wang et al. 153 successfully achieved in vivo decellularization of partial liver lobes in rats with a good survival rate through the perfusion of 1% SDS solution. Although recellularization and anti-coagulation processes still require further optimization, the study represents a promising strategy for liver engineering and regeneration in vivo. Hence, decellularization in vivo that removes injured or metabolism-disordered cells but maintains hepatic ECM might be a potential solution for liver tissue engineering and regeneration in situ. Cell aggregate-based liver tissue engineering has also shown some promising results for the rescue of various liver diseases due to their functional enhancements, however, only simple hepatic organizations have been successfully constructed.20,25 Therefore, recellularization using stem cell aggregates in decellularized whole liver organs may facilitate the recovery of normal hepatic functions and the reconstruction of complex liver architectures. Li et al. 154 previously showed that improved hepatic differentiation, as well as maintenance of hepatocyte-like cell morphology and viability, were achieved by employing human umbilical cord-derived mesenchymal stem/stromal cell (hUC-MSC) aggregates cultured in the decellularized porcine liver scaffolds, suggesting the great benefits of the novel approach that combines stem cell aggregates and decellularized liver scaffolds for liver tissue engineering and regeneration.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Key Research and Development Program of China (2019YFA0111300), the Guangdong Province Science and Technology Innovation Special Fund (International Scientific Cooperation, 2018A050506035), the National Natural Science Foundation of China (51903256, 21907113, 32001012), Research Grants Council (Hong Kong)-Early Career Scheme (24204819), and the Chinese University of Hong Kong Direct Grant (2018.018).