
Abstract
Brain diseases including neurodegenerative disorders and tumours are among the most serious health problems, degrading the quality of life and causing massive economic cost. Nanoparticles that load and deliver drugs and genes have been intensively studied for the treatment of brain diseases, and have demonstrated some biological effects in various animal models. Among other efforts taken in the nanoparticle development, targeting of blood brain barrier, specific cell type or local intra-/extra-cellular space is an important strategy to enhance the therapeutic efficacy of the nanoparticle delivery systems. This review underlies the targeting issue in the nanoparticle development for the treatment of brain diseases, taking key exemplar studies carried out in various in vivo models.
Introduction
Brain diseases including Alzheimer’s, Parkinson’s, Huntington’s and brain tumours are globally challenging issues. While therapeutics has been significantly developed for the treatment, recent nanotechnology emerges as a potent tool to deliver therapeutics to brain. Among others, nanoparticle (NP)-mediated drug delivery has been considered promising because the NP can be tailored to target the receptors expressed on cells while incorporating and releasing the therapeutic molecules in a controlled manner.1–4
Among other obstacles for the NP to overcome, blood brain barrier (BBB) is the first that segregates the inner central nervous system (CNS) from blood. 5 For the BBB penetration, the surface of NPs has been designed with various kinds of molecules such as antibodies and peptides to recognize the receptors and transporters expressed on BBB. Furthermore, the BBB-crossed NPs need to reach the diseased site. Therefore, NPs are further tailored to target specific type of cells or the surrounding environment. For example, in brain tumours, NPs are modified to recognize the cancer cells to enable targeted delivery of anticancer drugs with improved therapeutic efficacy. Also, NPs can be designed to target the intracellular component mitochondria to ameliorate oxidative stress induced neuronal cell death and to recover neural functions. 6 Furthermore, the extracellular molecule amyloid beta (Aβ), considered as the major cause of Alzheimer’s disease, is often targeted by engineered NPs, 7 which leads to degradation of Aβ more effectively.
The schematic Figure 1 summarizes the NPs developed for targeting, either specific cell type or intra-/extra-cellular space. NPs that circulate in a blood stream need to cross BBB and to target specific cell type such as neuron, astrocyte, oligodendrocyte, microglia and tumour cell, and sometimes to localize to intracellular space (e.g. mitochondria, nucleus, endoplasmic reticulum), or to scavenge extracellular molecules, such as amyloid beta plaques in Alzheimer’s disease. In this mini-review, among other issues in NP development for brain diseases, we focus on the targeting effort that is to enhance the penetration of BBB, and to localize diseased site such as cells and extracellular molecules. For this, the studies carried out mainly in the in vivo models are surveyed and the results and significance are communicated.

Nanoparticles with targeting ability used for the brain diseases. Nanoparticles circulating in a blood stream (a) need to cross BBB (b), and then localize to target cells (e.g. neuron, astrocyte, oligodendrocyte, microglia, tumour cells) (c), and sometimes target cellular organelles (e.g. mitochondria, synaptic cleft) (d) or extracellular molecules (e.g. amyloid beta plaques in Alzheimer’s disease) (e).
Targeting BBB
BBB consists of brain capillary endothelial cells (EC) tightly connected to each other and has low permeability (fluid phase endocytosis), allowing very limited entry of some specific molecules. 8 Most of molecules or particles, except water, glucose or amino acids, are almost impermeable to BBB, restricting the effective treatment of brain tumours and CNS disorders with drugs and NPs. 9
For the BBB penetration, the surface of NPs has been modified. Some of the highly positive NPs were found to cross BBB through adsorptive and receptor/transporter-mediated transcytosis. Cell penetrating peptide TAT or positively charged albumin proteins were often used to coat NPs to enable electrostatic attachment to BBB and penetration.10,11 Another family of cell penetrating peptide (SynB peptides RGGRLSYSRRRFSTSTGR, Penetratin, Mastoparan) was also reported to increase the drug permeability in a co-cultured BBB model, and to double the drug transport in vivo. 12 However, the stability of cell penetrating peptides in the in vivo conditions is reduced significantly with time that eventually reduces the activity and practical applications.
Receptor-mediated transcytosis is another way that NPs can cross BBB. Brain can uptake transferrin, insulin, apolipoproteins and 2-myoglobulins through receptor-mediated pathways. In many cases, BBB penetrating vectors (proteins, antibody, peptides, etc.) are attached to NP surface to help receptor or transporter-mediated transcytosis. Transcytosis through EC barrier starts with either clathrin coated pits, caveolae or caveolae-like membrane domains similar to endocytosis in normal cells. Clathrin-mediated transcytosis leads the NP to late endosome or lysosome. Caveolae-mediated internalization is a slow process but the main difference is that it does not result in a pH decrease and not lead to a degradative pathway of their cargo, dissimilar to the endosomal/lysosomal pathway.13,14 Interestingly, the transcytotic potential was found to be higher for the receptor-mediated and caveolar-mediated pathways than for the adsorptive-mediated transcytosis.
Transferrin is a transmembrane glycoprotein and a widely studied receptor for brain targeting. Transferrin receptor consists of two linked 90 kDa subunits and each of them can bind transferrin molecule. Transferrin receptors have a specific function for brain, that is, regulation of uptake of iron via transferrin that is present as a plasma protein and transports iron in the circulation. As transferrin has high affinity to transferrin receptors, it is often functionalized on NP surface for brain targeting. Au NP with 80 nm in size and having nearly zwitterionic surface charge was conjugated with transferrin via acid cleavable linker for targeting brain. At endosomes, the transferrin molecules get detached from NP surface facilitating the entry of NP to brain. This methodology was found to enhance the brain uptake of NP compared to the previously reported method. 15
However, a real problem is that transferrin receptors are saturated with free endogenous transferrin in normal conditions and the transferrin modified NPs have to compete with the free transferrin. To avoid this, different antibodies are used against transferrin receptors. 16 These antibodies recognize different epitopes of the transferrin receptors, abolishing the binding competition with the endogenous transferrin. So far, many antibodies including OX26, 8D3 and R17217 have been reported. 17 They are all monoclonal antibodies and specifically bind to transferrin receptors and undergo receptor-mediated transcytosis. While these are used for brain delivery, there still exist some drawbacks. As the antibodies specifically recognize transferrin receptors, they can accumulate in other peripheral organs such as liver, spleen, bone marrow and kidney due to a widespread expression of transferrin receptors. 16 It was observed that 0.44% of injected dose of OX26 was accumulated in brain in 24 h whereas for 8D3 and R17217, the accumulation was 1.5% and 0.8%, respectively, after 1 h.
Apolipoprotein E (Apo E) is a 34 kDa protein that transports cholesterol and other lipids in the plasma and in CNS. The lipoproteins can be taken up by brains through recognition of Apo E via specific receptors on the BBB including low-density lipoprotein receptors (LDLRs) and LDLR-related proteins. Therefore, NPs functionalized with lipoproteins can be recognized by these receptors and undergo receptor-mediated transcytosis. This phenomenon was observed when polybutylcyanoacrylate (PBCA) NPs were coated with PS80 surfactant. PS80 surfactant was found to adsorb Apo E from blood serum and cross BBB in an in vivo mouse model.18,19 Apo E was also conjugated to albumin-NPs or liposomes, enhancing BBB passage in in vivo rodent models. 20
Like the transferrin receptors, insulin receptors are also found in luminal membrane of brain ECs and plasma membrane of brain cells. 21 They are responsible for the delivery of insulin to brain via receptor-mediated transcytosis. Insulin receptor is a 330 kDa membrane glycoprotein consisting of α and β subunits. 21 These two subunits are linked via disulphide bond, forming a cylindrical structure, and when insulin binds, it undergoes a conformational change, activating tyrosine kinase and subsequently enabling receptor-mediated internalization. However, unlike transferrin, insulin has very short half-life in blood serum which restricts its use as a brain targeting agent. Therefore, similar to transferrin receptors, insulin monoclonal antibodies (83-14 antibody) have been used as brain targeting vectors. A fully humanized version of 83-14 antibody is called HIRA mAb that is used for lysosomal storage disorders. Mucopolysaccharidosis Type I or MPS-I disease is caused due to a deficiency of L-iduronidase, an enzyme responsible for the degradation of mucopolysaccharides in lysosomes. Here L- L-iduronidase is delivered by fusing with HIRA mAb for the treatment of MPS-I which causes 70% reduction of glycosaminoglycans in MPS-I patients. On the contrary, it was observed that 2% of L-iduronidase was able to reach the rhesus brain after 2 h of intravenous administration (Figure 2). 20
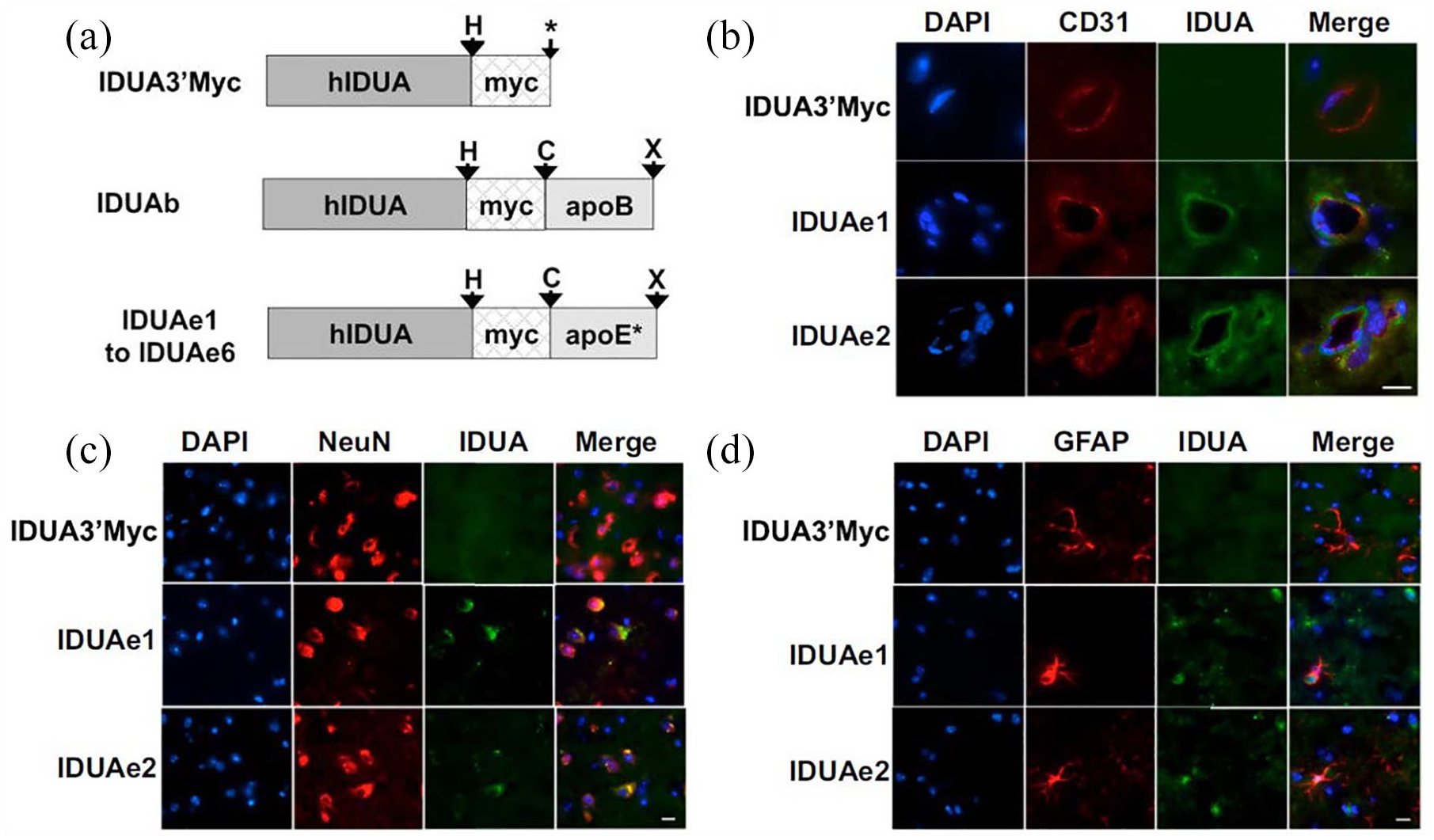
Targeting BBB: (a) Engineered IDUA proteins introduce LRP1-mediated endocytosis in genetically modified cell lines. Diagram of modified human IDUA proteins (hIDUA) fused in-frame with the myc-tag and the receptor-binding domain of apoB or various peptides from apoE (apoE*). (b–d) Liver-generated IDUA fusion proteins in the circulation can be transported into the CNS and normalize the accumulation of GAG in the brain of MPS I mice. Whole brains were collected from well perfused animals 2 days after hydrodynamic injection. Samples were stained with antibodies against IDUA protein (green) and endothelial marker (CD31; red) (b), terminally differentiated neuron marker (NeuN; red) (c), or astrocyte marker (GFAP; red) (d). Reproduced with permission from Wang et al. 20
Rabies viruses are neurotropic that have to cross BBB to infect the brain. These viruses have developed five proteins: nucleoprotein, phosphoproteins, glycoproteins, matrix protein and RNA polymerase. Among them rabies virus glycoproteins enable virus to enter brain. It is found that rabies virus binds specifically with acetylcholine receptors to cross BBB. For example, silica-coated Au nanorod was conjugated with RVG29 peptide to deliver into a mouse brain for the treatment of brain glioma. Here the Au nanorod was utilized for photothermal therapy and the rod shape could enhance the interaction with receptors. In another study, RVG29 was used to design dendrigraft poly-L-lysine NPs loaded with caspase-3 shRNA-coding plasmid (RVG29-DGL-shRNA) for the treatment of Parkinson’s disease (PD) in rat model. Exosomes were also designed with RVG29 to deliver siRNA to brain. For example, BACE-1 siRNA was delivered to brain using RVG29 designed exosomes for the treatment of Alzheimer’s disease (AD). 22
Brain cannot synthesize glucose by itself so glucose uptake into brain is essential for sustaining cerebral functions. Therefore, different glucose transporters are expressed on ECs in brain which are responsible for transporting glucose in or out of brain. A number of GLUT transporters (GLUT1-7) are found to express on BBB that transport glucose D enantiomers to the brain. There are many reports where glucose is conjugated with NPs or liposomes to cross BBB and deliver drug molecules inside CNS. For example, glucose-modified liposomes with different chain lengths of polyethylene glycol (PEG) were reported. Long PEG chains were shown to prolong blood circulation time, increasing the possibility to target brain, but too long PEG chains were highly flexible to fold together, which reduces targeting efficiency. Therefore, GLU1000-LIP was shown to be more efficient to target brain when compared with GLU2000-LIP. 23 Glucose-coated Au NPs with a size of 4 nm were also capable of penetrating BBB to reach astrocytes. Although the NPs did not follow any receptor- or transporter-mediated endocytosis, the glucose coating was necessary for efficient penetration of BBB. Similarly, glutathione-coated Au NPs were developed with the same size, which however were not able to penetrate BBB more efficiently than the glucose-coated one. Also, similar surface functionalization of 30 nm Au NP could not penetrate as efficiently as the 4 nm Au NP, underscoring the importance of NP size. 24
Targeting cells in brain
In brain, there are different types of cells, including neurons, astrocytes, oligodendrocytes and microglia. The rationale on targeting cells with NPs comes from their different functions which need specific therapeutic molecules at proper doses.
Neurons transmit signalling information through synapsis. Neuronal cell death often leads to various neurodegenerative diseases. There are different reasons for the neuronal cell death, such as necrosis/apoptosis, excitotoxicity and autophagic cell death. 25 Among them, excitotoxicity causes excessive synaptic release of glutamate which activates postsynaptic receptors such as N-methyl-d-aspartate (NMDA), 2-amino-3-(3-hydroxy-5-methylisoxazol-4-yl) proprionate (AMPA) and kinase receptors. Therefore, targeting neurons to reduce the excitotoxicity and neuronal cell death helps preventing various diseases. Adenosine plays a neuroprotective role. It activates adenosine A1 receptors which inhibit Ca2+ influx and activate phospholipase C, consequently inhibiting the secretion of various neurotransmitters, such as glutamate, dopamine, serotonin and acetylcholine.25,26 However, the delivery of adenosine to brain is very difficult. An exemplar study by Gaudin et al. demonstrated using lipid nanocarrier Squalenoyl adenosine. The nanoassembly did not induce any change, either in daily food intake behaviour or in weight variations of the animals, compared to the controls. Moreover, in vitro and in vivo studies demonstrated that the nanoassembly could extend adenosine circulation and its interaction with the neurovascular unit, leading to significant improvement of neurologic deficit score in cerebral ischaemia and early motor recovery of hindlimbs in spinal cord injury by the targeting effect with nanoassembly (Figure 3). 27 Mishra et al. 28 also delivered valproic acid and N-acetyl cysteine (NAC) conjugated with dendrimers which could effectively localize to the injured neuronal sites in brain while significantly reducing side effects.

Targeting neurons with lipid nanocarrier Squalenoyl adenosine: (A) Squalenoyl adenosine (SQAd) was prepared in a three-step synthesis, and the SQAd nanoassemblies were obtained by nanoprecipitation. (B) Ischaemic volumes in control and treated mice subjected to transient (2 h MCAo and 22 h reperfusion) and permanent (24 h MCAo) focal cerebral ischaemia were identified by reduced Nissl staining under a light microscope. A significant therapeutic effect was also observed when SQAd nanoassemblies were administered 2 h post-ischaemia in the permanent MCAo model. (C) The pharmacological efficiency of the SQAd nanoassemblies was assessed in a T9 contusion SCI model. After 72 h, the SQAd nanoassemblies-injected animals showed a complete recovery of their hindlimbs, in accordance with the absence of visible traumatic area on the cord (C, c) compared with the trauma group (C, a) and the adenosine-treated group (C, b). Reproduced with permission from Gaudin et al. 27
On the contrary, astrocytes are the glial cells that perform various functions in brain such as the maintenance of extracellular microenvironment, metabolisms, communication with neurons and BBB endothelium, and the modulation of excitatory synaptic transmission.29–31 Targeting astrocytes can control the neurotransporters (e.g. glutamate, γ-aminobutyric acid, glycine and histamine) that participate in neurotransmitter release and thus are crucial for the termination of synaptic transmission and maintenance of neuronal excitability. Astrocytes can protect CNS against various insults and diseases.32,33 Thus, targeting astrocytes can minimize the toxic effects of reactive oxygen species (ROS) and reactive nitrogen species (RNS) that are substantially generated upon brain trauma and injuries34,35; thus, future studies on targeting astrocytes with NPs may help recovering those functions in brain, which, though, are yet to be realized.
Oligodendrocytes are specialized neural cells of which the role is to myelinate the axons in CNS. Myelin sheaths are large extensions of the oligodendrocyte cell membrane and have a highly complex structure. 36 The myelin sheaths consist of 80% lipid providing insulating properties and 20% proteins which stabilize the structure. Demyelination of axons results in prevention of electric signal and removes the metabolic support which leads to neurodegeneration. To target the myelination of cells, Rittchen et al. 37 used poly(lactic-co-glycolic acid)(PLGA) NPs loaded with leukaemia inhibitory factor (LIF). The PLGA-LIF NPs could target NG-2 chondroitin sulphate proteoglycan, which is expressed on oligodendrocyte precursor cells (OPCs). The NG2-targeted LIF-NP induced OPC differentiation into mature oligodendrocytes in the in vitro culture condition and increased myelin repair in the in vivo focal CNS demyelination model. This study highlights that the targeting of OPCs with NP-based delivery system was highly effective in promoting myelin repair by directly altering the fate of progenitor cells, implying a promising strategy for restoring electrical signal and metabolic support in degenerative nerves (as shown in Figure 4).
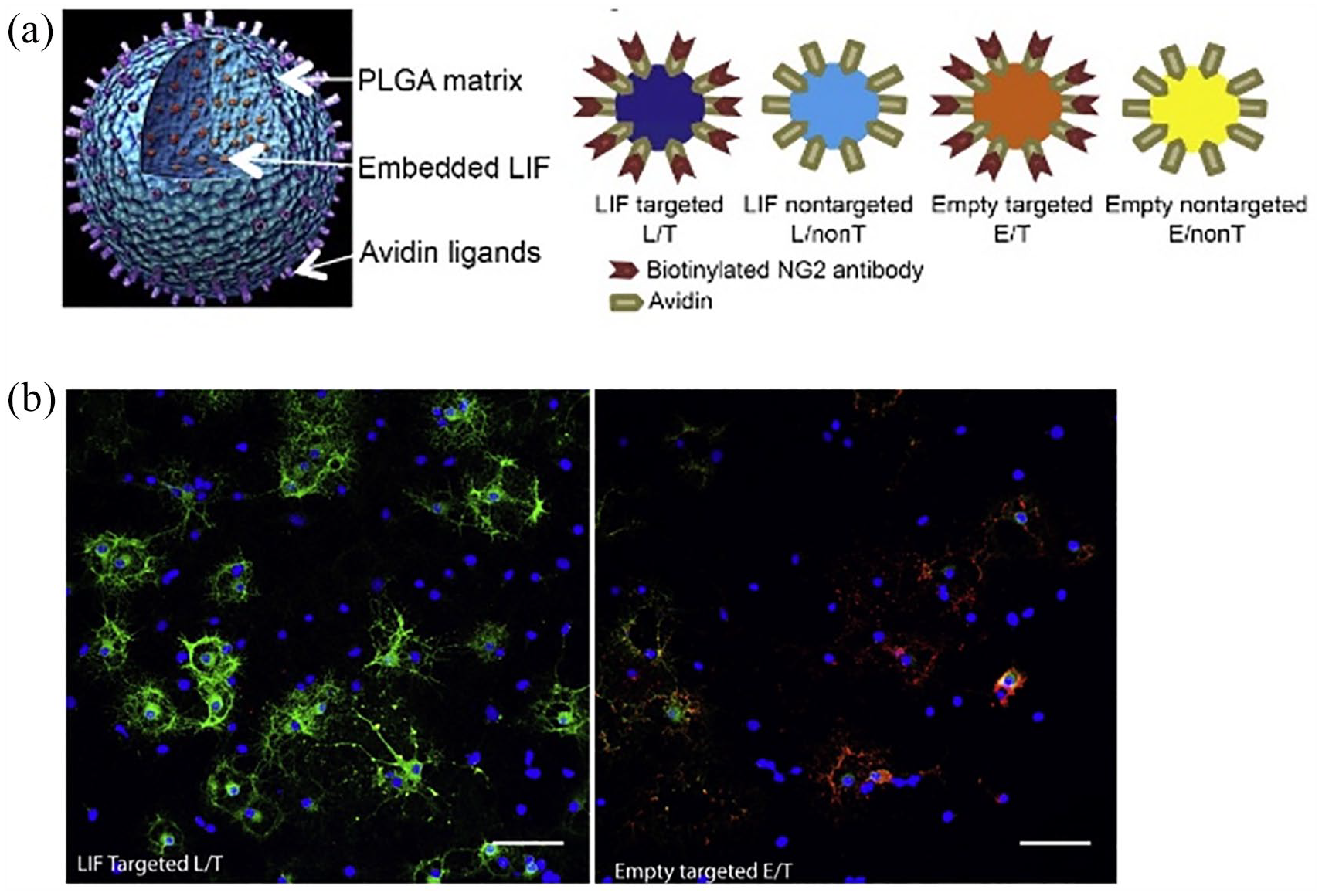
Targeting of glia cells with LIF-NP through STAT-3 signalling in OPC: (a) Composition of PLGA-based NP with embedded LIF and surface avidin for attachment of biotinylated targeting antibody. (b) OPC mature into OD in response to NG2-targeted LIF-NP. Reproduced with permission from Rittchen et al. 37
In order to mediate neuro-inflammatory diseases, targeting microglia is a proper strategy. Microglia are the immune cells that play an important role in brain inflammation and infection. They act as macrophages and clear infectious agents, dead neurons and plaques in CNS. As other macrophages, microglia become activated upon surrounding changes in immune response, presenting different phenotypes (e.g. M1 and M2 phase). While the M1 phenotype is considered pro-inflammatory that produces cytokines such as tumour necrosis factor-α (TNF-α), IL-1β and IL-6, the M2 phenotype generates cytokines TGF-β and IL-10 to relieve the inflammation.38,39 The activation of microglia (mostly M1 phenotype) aggravates inflammatory responses by producing excessive ROS, cytotoxic metabolites and pro-inflammatory cytokines resulting in severe brain damage. 39 NAC is used as anti-inflammatory and antioxidant agent and thus has a potential for neuroprotection. To enable targeted delivery of NAC to activated microglia, Kannan et al used hydroxyl-terminated generation-4-PAMAM dendrimers (Figure 5).40,41 When treated to cerebral palsy (CP), the PAMAM conjugated with NAC (D-NAC) could localize to activated microglia and astrocytes and demonstrated significant reduction of the white matter injury. The suppression of inflammation was assessed by quantifying nuclear factor κB (NF-κB), which induces transcription of pro-inflammatory genes and TNF-α, in the periventricular region (PVR) of brains removed from 5-day-old rabbits treated on day 1. A single dose of D-NAC10 led to a 3.5-fold decrease in NF-κB expression when compared to equivalent dose of the free drug (NAC10). A significant decrease in mRNA expression of TNF-α was also noted when treated with D-NAC (1 and 10 mg/kg) compared to the free drug alone.

Targeting microglial activation state as an anti-inflammatory strategy in neuro-inflammatory diseases: (a) Synthesis and characterization of the D-NAC conjugate. Reaction schematic for the synthesis of the dendrimer-NAC (D-NAC) conjugate 1, starting from the free dendrimer and free NAC. (b) Inflammation on day 5 after treatment on day 1. NF-κB and TNF-α mRNA levels in the PVR of the brain. Western blot of NF-κB p65 expression was quantified and normalized to b-actin. TNF-α mRNA was quantified and normalized to GAPDH expression. (c) The periventricular white matter regions of rabbit kits on day 5 of life were stained for all microglia with tomato lectin (red) and for pro-inflammatory microglia with anti-CD11b antibody (green). Reproduced with permission from Kannan et al. 41
Targeting cells in brain tumours has been intensively studied. The most common form of brain tumour is glioblastoma that originates from astrocytes. The general strategy to treat glioblastoma is the targeting of tumour mass by active or passive targeting processes. The passive targeting delivery can reach the tumour sites when no targeting ligands are attached on NP surface. An enhanced permeation and retention effect is considered a major mechanism of the passive tumour-targeted systems and highly depends on the tumour biological parameters such as perivascular tumour growth and intratumor pressure. On the contrary, active targeting can significantly increase the quantity of drug delivered to the targeted tumour compared to passive targeting. Different active-targeting strategies have been developed and the CD44 receptors overexpressed on glioblastoma cells are one of the major targets.42,43 One of the most common molecules is CD44 that is a receptor of hyaluronic acid where the CD44 targeting motif exists. The RGD peptide has also been used to target integrins. For example, Liu et al. 44 used RGD peptide conjugated with octa arginine (R8) where the RGD peptide recognizes the integrin receptors on brain capillary ECs while the conjugated R8 enhances the penetrating capability. The liposome carrier conjugated, R8-RGD (R8-RGD-lipo), was found to deliver paclitaxel drug with significantly enhanced capacities (2- and 30-fold, respectively, by R8 and RGD). When systemically administered to C6 glioma bearing mice, the survival rate of mice was significantly enhanced. Quader et al. 45 also designed cRGD-conjugated nanomicelle loaded with epirubicin through a pH responsive link (Figure 6). The presence of cRGD could enhance the penetration of nanomicelles through three-dimensional spheroid of U87MG cells. The intravenous injection of cRGD-conjugated epirubicin-loaded nanomicelles significantly suppressed the growth of orthotopic glioblastoma tumours. Boron neutron capture therapy (BNCT) has also recently emerged as a promising tool, but the most significant challenge in BNCT is specific targeting of brain tumour cells. Kuthala et al. aimed to tackle this by the RGD approach. They synthesized 10 B-enriched (96% 10B enrichment) boron NPs (10BSGRF NPs) and modified with a fluorescein isothiocyanate–labelled RGD-K peptide. 46 The NPs could effectively inhibit the brain tumour while allowing the contrast of magnetic resonance imaging to detect the location, size and progress of brain tumour.

Targeting brain tumours with NP: (a) Preparation of cRGD-peptide installed epirubicin-loaded polymeric micelles from a mixture (1:3) of cRGD-poly(ethylene glycol)-b-poly(hidrazinyl-aspartamide) and MeO-poly(ethylene glycol)-b-poly (hidrazinyl-aspartamide) polymer. (b) Penetration of epirubicin-loaded micelles (Epi/m) and cRGD-installed Epi/m (cRGD-Epi/m) in U87MG spheroids. Red: Fluorescence from epirubicin. (c) Antitumor activity against bioluminescent orthotopic U87-MG-luc tumours in mice injected with PBS, Epi, Epi/m and cRGD-Epi/m. Reproduced with permission from Quader et al. 45
Targeting intracellular components
Not only the type of cells, but the intracellular organelles are the sites to target for effective therapeutics. Mitochondria play a central role in ageing-related neurodegenerative diseases.47–49 Mitochondria produce ATP that is required for different structural and metabolic activities. For example, axons demand high energy to maintain the ionic gradient and to keep structural integrity to support the neurotransmission. Furthermore, myelination process requires cholesterol and other complex lipids that are produced from CoA synthesized during mitochondrial beta-oxidation.50–52 Therefore, the mitochondrial dysfunction is often involved with the oxidative stress that leads to severe damage in CNS, such as disruption in oligodendrocytes differentiation, viability to form myelin sheaths.
For these reasons, targeting mitochondria has been sought to gain therapeutic efficacy in various neurodegenerative diseases. For example, ceria NPs (CeO2) conjugated with triphenyl phosphonium (TPP) were used to target mitochondria of neuronal cells and to supress the neuronal death of transgenic Alzheimer’s mouse (Figure 7). 6 Ceria NPs have strong and recyclable ROS scavenging activity which reduces the ROS production and mitochondrial dysfunction. Polyanhydride NPs loaded with antioxidant apocynin were also used to target oxidative stress induced neuronal cell death. 53 Another exemplar study used poly(N-vinylpyrrolidone)-b-poly(ε-caprolactone) polymer to load antioxidant resveratrol (RES) and deliver to neuron cells though endocytosis. The RES-loaded polymer carrier showed neuroprotection effects at significantly lower doses when compared to the free RES. 54 Furthermore, the NPs made of acidic PLGA were developed to restore impaired lysosomal function in PD. The acidic PLGA NPs were shown to transport to the lysosome within 24 h, lowering lysosomal pH and rescuing chloroquine-induced toxicity. Also, after intracerebral injection in neurons, the NPs could attenuate PD-related neurodegeneration in vivo by mechanisms involving a rescue of compromised lysosomes, 55 underscoring the effective strategy of targeting lysosomes to restore neurodegenerative functions by means of the NP development.

Targeting mitochondria with NP: (a) Design, synthesis and ROS recyclable scavenging activity of TPP-ceria nanoparticles as a therapeutic mitochondrial antioxidant for Alzheimer’s disease. (b) TPP-ceria NPs significantly inhibit Aβ-induced mitochondrial ROS in SH-SY5Y cells. (c) TPP-ceria NPs reduce reactive glial activation. Reproduced with permission from Kwon et al. 6
Targeting extracellular space
The synaptic sites of brain cells are the right target to regulate the receptor activities. A recent study by Savchenko et al showed the effective role of NPs by means of controlling the NP size, in the modulation of synaptic transmission signalling when properly targeted (Figure 8). The memantine conjugated Au NP hybrid nanodrug (AuM) was developed to preserve the physiological role of N-methyl D-aspartate receptors (NMDARs) in synaptic transmission while blocking only excessive pathological activation of NMDARs. 56 Because of its larger size (35 nm) than synaptic cleft, the engineered AuM could not access the synaptic NMDARs but could only access the external NMDARs which are the target for inhibiting pathological activation of cells. The results could demonstrate that the AuM selectively acted as an external NMDAR antagonist, and the study offered a new arena for studying neurotransmitter receptors through the selective targeting of extracellular space (not intracellular) with size-controlled NPs.

Targeting neurotransmitter receptors with NP: (A) Design of hybrid nanostructured antagonist of AuM for exclusive inhibition of eNMDARs. (B) AuM is an efficient antagonist of NMDARs as demonstrated by representative calcium imaging traces from cerebrocortical neurons activated by 200 μM NMDA and inhibited by 10 nM AuM (left) or 10 μM memantine (right) (B, a), whereas AuM is not effective in inhibiting glutamatergic synaptic activity as demonstrated by representative calcium imaging traces in spontaneously active neurons in the presence of 10 nM AuM (left) and 10 μM free memantine (right) (B, b). Reproduced with permission from Savchenko et al. 56
Targeting amyloid beta (Aβ) plaques is a key strategy to treating AD since its accumulation in brain is considered a major cause of AD. NPs developed to interact with the peptides and clear them out of brain or inhibit their formation can thus offer a solution to this. As the peptide can leave the brain by crossing BBB with the help of various apolipoproteins including apolipoprotein E2/3 (Apo-E2/3) and Apo-E4, many NPs have been designed to bind the peptides in blood, which ultimately induces the efflux of Aβ and reduces the level of peptides in brain. The PEGylated polymeric NP that has a long circulation time in blood and strong binding affinity with Aβ is a representative example. 57
Huang et al. 58 developed monosialotetrahexosylganglioside (GM1)-modified reconstituted high-density lipoprotein (GM1-rHDL) that shows high binding affinity to Aβ monomers and oligomers and facilitates Aβ degradation by microglia, and Aβ efflux across the BBB (Figure 9). After intranasal administration, the uptake efficiency of GM1-rHDL was found to be higher than rHDL in cortex + hippocampus. Due to higher binding affinity, GM1-rHDL can perform proteolytic digestion of Aβ and efflux across BBB. To determine the degradation of Aβ1-42 by microglia, the microglial cells were co-incubated with GM1-rHDL and rHDL. The undegraded Aβ1-42 was found to be significantly reduced in case of Aβ1-42 compared to rHDL. Similarly, Song et al. 59 have shown that apolipoprotein E3 modified HDL (ApoE3-rHDL) exhibited high binding affinity to Aβ1-42 monomers and oligomers and also facilitated microglial cells, liver cells and astroglial cells degradation of Aβ by accelerating lysosomal transport. ApoE3-rHDL was highly efficient to cross BBB and to facilitate intracerebral Aβ clearance by glia-mediated degradation. Here the co-incubation of Aβ with ApoE3-rHDL could facilitate the lysosomal transport of Aβ for degradation. After 3 h, the undegraded Aβ was significantly lowered in case of Aβ + ApoE3-rHDL treated cells compared to only Aβ treated cells. This suggests that ApoE3-rHDL leads to intracellular degradation of Aβ in both microglia and astrocytes.

Targeting amyloid beta (Aβ): (A) The evaluation of the binding affinity of rHDL and GM1-rHDL to Aβ1-42 monomers and oligomers by surface plasmon resonance analysis. (A, a and A, c) Binding of rHDL and GM1-rHDL to Aβ1-42 monomers, respectively. (A, b and A, d) Binding of rHDL and GM1-rHDL to Aβ1-42 oligomers, respectively. (B) Effects of rHDL and GM1-rHDL on the intracellular distribution, and degradation of Aβ1-42 in microglia. Colocalization of FAM-Aβ1-42 with DiI-rHDL or DiI-GM1-rHDL after 4 h of co-incubation. Arrow: FAM-Aβ1-42 colocalized with GM1-rHDL. (C) αNAP-GM1-rHDL reduced Aβ deposition (brown signals as indicated by arrowheads) in the hippocampus of AD model mice. AD model mice were daily intranasally administrated with rHDL, GM1-rHDL and αNAP-GM1-rHDL at the lipid dose of 5 mg/kg for 2 weeks. The brain sections were immunostained with anti-Aβ antibody 6E10. Reproduced with permission from Huang et al. 58
Concluding remarks
Although therapeutic molecules have been extensively developed, their delivery to brain remains a challenging issue. NPs hold great promise for delivering therapeutic molecules to brain with proper tailoring of size, shape and surface chemistry that allows the delivery into brain tissues. Among other strategies in NP development, targeting is an important approach to enhance the therapeutic efficacy. In brain, there exist different types of cells including neurons, astrocytes, oligodendrocytes and microglia, so the targeting of these cells with NPs can regulate specific cellular functions, such as synaptic signalling, neuroprotection, myelination and immune modulation, by delivering relevant therapeutic molecules at proper doses while reducing significant side effects. Not just cell type, but the intracellular organelles such as mitochondria is also a promising target to improve the therapeutic efficacy, such as the ameliorated oxidative stress in severe neuro-inflammatory conditions. Targeting synaptic site and extracellular molecules (such as amyloid beta plaques) has also been demonstrated to be effective in regulating neurotransmitter receptors and in the treatment of AD, respectively. As discussed, the use of engineered NPs helps crossing BBB and localizing therapeutic molecules to specific cells and sites in brain. Thus far however, the NP-based drug delivery system has not been translated for the treatment of brain diseases, although there are several Food and Drug Administration–approved nanodrugs for other purposes. The potential toxicity and safety concern of nanomaterials may be one key issue to be solved for future clinical applications. Also, the advances in our understanding of the NP interactions with BBB and brain tissues and the preclinical studies on more relevant and large animal models may accelerate the translational research of CNS-targeting nanocarriers. Furthermore, together with the delivery of therapeutic molecules, the rapidly progressing imaging techniques will further reinforce the potential of NPs for the treatment of intractable brain diseases in the near future.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship and/or publication of this article: This research was supported by the National Research Foundation of Korea (NRF) from Republic of Korea (GRDC Programme 2018K1A4A3A01064257, 2018R1A2B3003446 and 2018R1D1A1B07048459).