
Abstract
Extracellular vesicles comprise a heterogenous population of exosomes and microvesicles that have critical roles in intercellular signalling and tissue development. These complex particles have been implicated as mediators of the therapeutic effects of stem cells via the transfer of an assorted cargo of proteins and nucleic acids, which can modulate inflammation and enhance endogenous regeneration in a range of tissues. In addition, extracellular vesicles have the capacity to be loaded with therapeutic molecules for targeted delivery of pharmaceuticals. The versatility, biostability and biocompatibility of extracellular vesicles make them appealing for regenerative medicine and may endow considerable advantages over single molecule approaches. Furthermore, since production can be optimised and assessed ex vivo, extracellular vesicles present a decreased risk of neoplastic transformation when compared with cell-based methods. To date, the contribution of vesicles to tissue development has perhaps been most comprehensively defined within hard tissues, such as endochondral bone, where they were first identified in 1969 and henceforth referred to as matrix vesicles. Within developing bone, vesicles function as vehicles for the delivery of pro-osteogenic factors and initiate early nucleational events necessary for matrix mineralisation. However, advancement in our understanding of the biogenesis and characterisation of matrix vesicles has occurred largely in parallel to associated developments in wider extracellular vesicle biology. As such, there is a requirement to align current understanding of matrix vesicle–mediated mineralisation within the context of an evolving literature surrounding exosomes and microvesicles. In this review, we present an overview of current progress and opinion surrounding the application of vesicles in regenerative medicine with a primary focus on their potential as an acellular approach for enhancing hard tissue regeneration. This is balanced with an assessment of areas where further development is required to maximise their application for regenerative medicine.
Keywords
Introduction
Stem-cell therapies have shown great promise for the regeneration of a wide range of tissues lost to trauma or disease. Several approved stem-cell therapies are currently undergoing clinical trials, and routine procedures such as blood transfusions and bone marrow transplantation have now been applied in clinical practice for decades. However, despite the significant promise offered by these approaches, there are considerable limitations restricting their wider application. These restrictions primarily relate to the low availability of stem cells within donor tissues, loss of potency following expansion, regulatory issues surrounding translation and even questions surrounding their precise mode of action (MoA). 1 Significantly, there is evidence to suggest that only a small percentage (1%–3%) of stem cells actually engraft to the host tissue and that the majority may not reach their target sites, instead becoming trapped in the lungs, spleen and liver. 2 Following a recent paradigm shift, it has been proposed that the therapeutic effects of stem cells are largely evoked through paracrine activity rather than through engraftment and differentiation. A growing body of evidence suggests that paracrine activity is exerted through the action of trophic factors as well as nano-sized bioactive particles termed extracellular vesicles (EVs).3–6
Over the last decade, EVs have demonstrated considerable promise as naturally derived nanoparticles that can be utilised for a number of therapeutic applications, which include predictive and regenerative medicine (Figure 1). EVs are ubiquitous within biological fluids (e.g. blood, urine, semen, milk and amniotic fluid), contributing to critical physiological and pathological processes such as tissue development, regeneration, inflammation and cancer metastasis. 7 Knowledge of the biological contribution of EVs to health and disease dates back to the 1960s.8–10 However, only in the last decade has the potential of these multifaceted particles become fully appreciated, with an exponential increase in the number of publications featuring ‘extracellular vesicles’ and/or ‘exosomes’ (Figure 1). Interest has aligned with the establishment of the International Society for Extracellular Vesicles (ISEV) and its affiliated journal, the Journal of Extracellular Vesicles. In line with growing research activity, position statements have been published outlining guidelines for the definition and analysis of EVs.11,12 These developments are reminiscent of events in the field of stem-cell biology less than a decade earlier when the Mesenchymal and Tissue Stem Cell Committee of the International Society for Cellular Therapy outlined the minimal criteria for defining human mesenchymal stem cells (MSCs). 13 Mirroring progress previously observed with the expansion and translation of stem-cell therapies, a framework is now beginning to be established through which EVs may have a pronounced influence on the future direction of medicine. 14 Much like stem cells, these complex bioactive particles can be combined with a bioactive scaffold or delivery system to enhance endogenous tissue regeneration and resolve inflammation in a host of tissues, including the skeletal system. 15 In this review, we will outline current knowledge surrounding EVs, summarising their critical roles in skeletal development, and highlight the advantages and challenges of an EV-based approach to regenerative medicine.

Publication trend of extracellular vesicles. Data were exported from Web of Science using the following criteria: (1) all databases, keywords (exosomes OR extracellular vesicle) and year range (1930–2017); (2) all databases, keywords (exosomes OR extracellular vesicle AND therapy) and year range (1930–2017).
EVs
History
Wolf and colleagues first acknowledged the presence of phospholipid-rich particulates within platelets, originally classifying the material as ‘platelet dust’ (Figure 1). It was noted that these particulates could be separated by ultracentrifugation (UC) and displayed coagulant properties. 8 In 1967, Anderson and Bonucci observed electron-dense ‘leaf-like’ particles with ‘needle-like’ projections that were attached to collagen fibrils within ossifying cartilaginous matrix, which they subsequently defined as matrix vesicles (MVs).9,10 In the 1980s, the physiological contribution of EVs was found to be more widespread, with these nano-sized messengers shown to mediate important immunological processes, 16 such as antigen presentation 17 and anti-tumour activity. Today EVs are known to play a wide role in intercellular communication and development within a number of hard and soft tissues, where they deliver a cargo of nucleic acids, peptides and lipids to neighbouring cells and tissues. The provision of these heterogeneous biological cargos has been observed to have a downstream effect on a variety of molecular functions, 18 including the signalling and regulation of gene expression in target cells, such as the upregulation of proteoglycan and type-II collagen by osteoarthritic chondrocytes. 19 More recently, the delivery of both osteoblast- and MSC-derived EVs has been shown to promote osteoblast differentiation and mineralisation both in vitro and in vivo. 20 Thus, suggesting these bioactive particles are capable of promoting de novo mineralisation and may have considerable potential for driving skeletal tissue regeneration.
Current definition
EVs are defined as phospholipid-enclosed nanoparticles (30–2000 nm) that carry a complex and variable cargo of biological contents including proteins and nucleic acids. In the context of bone formation, vesicles will also contain elements required to direct early mineralisation such as
To date, three different subtypes of EVs have been identified that are typically classified based on diameter and biogenesis (Table 1). Exosomes (30–150 nm) are formed within the endosomal network and are released when multivesicular bodies fuse with the plasma membrane. Microvesicles (50–1000 nm) are generated by outward budding (blebbing) of the plasma membrane and will contain only local cytosolic proteins and nucleic acids. Larger vesicles termed apoptotic bodies (500–2000 nm) are released as fragments of dying cells and can often be distinguished by the presence of nuclear particulates as a consequence of karyorrhexis. 21 Notably, there is some discrepancy in the literature regarding the size range of each EV subtype, with considerable overlap noted in their size and content. As a consequence, studies that define exosomes purely by size are likely more representative of a heterogeneous population comprising exosomes, small microvesicles and some additional non-vesicular extracellular material – depending on the efficiency of isolation, this may include small membrane fragments and large extracellular proteins. It is of considerable importance that vesicles derived from different tissue and biofluid sources are defined in accordance with published guidelines presented by the ISEV. 11 To further our understanding of the molecular composition of these EV subtypes, online resources are now available that provide a comprehensive and continually evolving database of protein, lipid and RNA analyses of EVs derived from a wide variety of cell and sample types. These can be found freely available online and include the databases Vesiclepedia (http://www.microvesicles.org), EVpedia (http://student4.postech.ac.kr/evpedia2_xe/xe) and ExoCarta (http://www.exocarta.org). Only by adhering to implemented standards and contributing to the developing body of resources can we continue to generate rigorous and reproducible research as well as develop EV therapeutics with translational potential.
Definition and features of extracellular vesicles.

Biological functions
The precise biological function of EVs is a reflection of the parental cell from which they were derived and the local microenvironment (e.g. inflammatory or hypoxic). Divergence in biological function can be demonstrated by the fact that they have been implicated in critical processes such as tissue formation as well as in pathological conditions, including vascular calcification and many forms of cancer such as osteosarcoma.18,22 There have been studies showing that MSC-derived EVs have an anti-inflammatory and immunomodulatory capacity, promoting macrophage polarisation and proliferation. 23 Furthermore, cardiac-derived EVs have been shown to be powerful stimulators of angiogenesis in endothelial cells during a myocardial infarction so they potentially hold promise for applications such as cardiac vessel regeneration. 24 EVs also play a prominent role within the nervous system, regulating myelin formation, neuronal outgrowth and survival, receptor recycling and the removal of pathological proteins, which makes them promising candidates for neuronal regeneration and the treatment of neuro-degenerative diseases. 25 However, the critical and long-defined contribution of these nanoparticles to early osteogenesis will be the focus of this review.
Role of EVs in hard tissue mineralisation
Matrix vesicles as sites of early mineral nucleation
Bone is the most ubiquitous mineralised tissue within vertebrates and is composed of an inorganic multi-substituted hydroxyapatite. Although the biological and physicochemical properties of bone are well described, many of the processes governing mineral formation, transport and deposition within the extracellular matrix (ECM) remain unclear. The pioneering studies of Anderson and Bonucci in 1967 were the first to identify the presence of what they referred to as matrix vesicles (MVs) within hypertrophic cartilage. These vesicles were described as electron-dense bodies that became anchored within the developing ECM. The studies were the first to suggest that matrix mineralisation was not restricted to collagen fibrils. 26 Since these initial findings, subsequent data have consolidated MVs as sites of initial calcification leading to the formation of early inorganic apatite crystals in epiphyseal cartilage in a range of species, including mice and guinea pigs. 27 Within the evolving body of literature, MVs (Figure 2) refer to vesicles of varied size (50–400 nm) that reside in the pre-mineralised matrix of dentin, cartilage and bone. 28 Although not comprehensively defined, MVs likely represent a mixture of exosomes and microvesicles containing specialised components required to direct ECM mineralisation (Table 1).29–31 Currently, no study has sought to define the independent contribution of distinct vesicle subsets during hard tissue mineralisation and few have begun to align MV theory within the context of the burgeoning field of EV biology. 32 Perhaps, our most advanced understanding of the biogenesis of MVs derives from a study by Boonrungsiman et al., 33 where calcium phosphate–containing vesicles were found to associate with the mitochondria, thereby implicating specific intracellular transport pathways rather than simple membrane blebbing, characteristic of microvesicle biogenesis, as a mechanism for MV-mediated mineralisation. Even though the contribution of MVs to early mineralisation events in tissues such as calcifying cartilage, bone and dentine can no longer be disputed, the underlying mechanisms by which this is achieved remain unclear. Here, we will present an overview of current opinion surrounding the role of MVs in physiological and pathological mineralisation.

Matrix vesicle (MV) cargo. Graphical representation of the matrix vesicle contents and membrane orientation of proteins, lipids and nucleic acids. Some of the listed components may be present in some matrix vesicles but not in others. For instance, in our previous study, we did not detect MHC complexes.
MVs are enriched in membrane proteins with well-defined roles in critical processes required for ECM mineralisation. These include the calcium-dependent phospholipid-binding annexin proteins (annexins II, V and VI) and enzymes including ectonucleotide pyrophosphatase/phosphodiesterase-1 (ENPP1) and alkaline phosphatase (ALP), which are required for maintaining the ratio of pyrophosphate (PPi) and inorganic phosphate (Pi) to regulate hydroxyapatite crystal formation
26
and drive matrix remodelling (Figure 3).
27
Under physiological conditions, mineralisation is inhibited by PPi generated through the cleavage of nucleotide triphosphates by ENPP1.
34
Inorganic Pi, derived through ALP-mediated hydrolysis of PPi, which drives mineralisation in both physiological and pathological conditions including vascular calcification, is loaded into MVs via transmembrane phosphate transporter proteins, such as Pit1, where these ions associate with calcium localised to the phospholipid-rich bilayer. Phospholipids comprising the MV membrane, including phosphatidylserine (PS), act in conjunction with membrane-binding proteins (e.g. annexins) to sequester
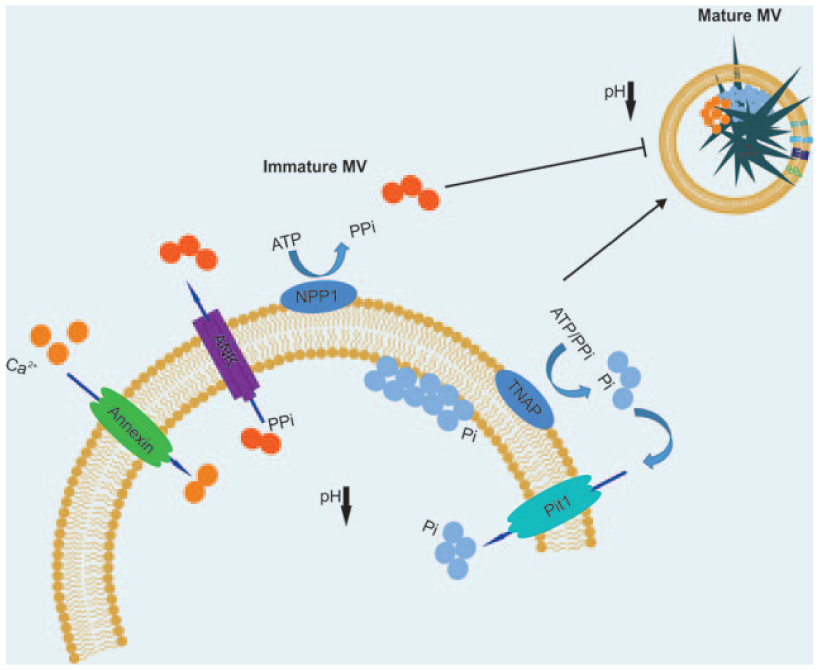
Schematic diagram of the mineralisation process. NPP1 inhibits mineralisation by generating PPi by catalysing extracellular ATP. TNAP promotes mineralisation by hydrolysing PPi into inorganic phosphate ions, which are in turn transported to the matrix vesicle (MV) through phosphate transporters such as Pit1. Conversely, ANK transports PPi from the MV into the developing ECM. Annexins function as calcium channels, transporting
As stated, MV calcification is tightly linked with the presence of acidic membrane phospholipids. 35 The presence of acidic phospholipids is significant in conferring membrane fluidity to promote fusion 34 and uptake of vesicles for intercellular communication, and in the transfer of signalling molecules between cells that comprise the skeletal niche. 11 It also has significant implications when one considers current opinions surrounding the formation of mature and physiologically stable hydroxyapatite from an amorphous precursor. 38 This transition is hypothesised to initiate from an amorphous precursor and proceed via a series of transient intermediate phases that include dicalcium phosphate dihydrate (DCPD) and octacalcium phosphate (OCP) that are only stable at a sub-neutral pH. Based on our published findings, we hypothesise the presence of acidic phospholipids acts to generate a localised reduction in pH, thereby facilitating the stable formation of intermediate phases such as OCP that can be detected analytically. 39 Intriguingly, the presence of DCPD and OCP has also been associated with pathological calcification in which an acidic pH is frequently encountered. Furthermore, chemical analysis of the mineral deposited during pathological calcification, particularly in medial and intimal sites of coronary arteries, has led researchers to suggest an active vesicle-mediated mechanism similar to that underpinning physiological early bone formation. 22 Vesicles localised to sites of vascular calcification derive from atrophying smooth muscle cells and recruited macrophages. The process is inflammation-dependent and associated with the rupture of vulnerable plaques. 40 Significantly, there is evidence to suggest that not all plaque-associated vesicles are harmful, with non-calcifying fetuin-A+ and matrix Gla protein+ vesicles also identified. The presence of vesicles containing these calcification inhibitors is likely to be important for maintaining vascular health, and methods for enhancing their presence in patients at risk of atherosclerosis could have significant clinical implications.
Clinical perspective
A growing body of evidence has accumulated implicating EVs as complex biological mediators of tissue development and regeneration. As such, there exists the potential to harness the regenerative capacity of these natural particles and exploit them to develop acellular yet complex biological approaches for driving tissue regeneration. At present, research into the regenerative application of EVs is in its infancy and the majority of documented research remains pre-clinical, with few clinical trials registered (Table 2). However, interest is rapidly growing with a number of recent publications outlining the potential therapeutic utility of EVs for the regeneration of a wide range of tissues, including bone and cartilage. In this section, we shall discuss current research into EV-mediated tissue regeneration and highlight the promise and current limitations of this approach.
Registered clinical trials in clinicaltrials.gov.

Keyword: exosomes. The inclusion criteria were the following: recruitment status (completed), age (all), sex (all) and study type (interventional).
There are several significant advantages in applying EVs for regenerative medicine when compared to conventional biological approaches that typically incorporate a cell or growth factor to encourage endogenous tissue formation. Firstly, vesicles may be superior to single approaches since they allow the delivery of multiple physiologically relevant factors and, as they are naturally derived, confer enhanced biocompatibility and biostability. 41 The fact that EVs deliver multiple biomolecules to recipient tissues may also enable them to target several therapeutic pathways simultaneously, thus enhancing their therapeutic efficacy. Another advantage is that EVs do not contain replicative machinery, which is significant in that it may allow for genetic modification of the parent cell line without the transmission of manipulated DNA into the vesicle. This can be applied to confer advantages over primary cell-based approaches, such as decreased population doubling times and phenotypic stability, which have significant implications when one considers the limited therapeutic window of primary cell therapies. The third benefit derives from the fact that EVs are not dynamic and cannot change phenotype, which has significant advantages when predicting therapeutic outcomes, ensuring reproducibility and reducing the risk of neoplastic transformation. In addition, the relatively small diameter of EVs means that they are less likely to become trapped inside the lungs, liver and spleen if administered intravenously and their innate homing ability increases the likelihood of these drugs reaching the target site. This makes EVs appealing as vehicles for loading pharmaceuticals and/or other therapeutic compounds to reduce drug clearance rates, with the aim of enhancing or extending their efficacy. In addition, by tailoring vesicle properties such as lipid composition, there may be the opportunity to further improve the stability of the nanoparticle while in circulation and increase their half-life when delivered locally or intravenously. Overall, EVs manufactured ex vivo may offer many of the benefits of a cell-based approach while avoiding the inherent pitfalls. The benefits of EVs as naturally derived particles for enhancing tissue regeneration have been documented in a wide range of tissues. We shall next explore current literature surrounding the application of EVs in soft and hard tissue regeneration.
The majority of studies applying EVs for regenerative applications have utilised vesicles derived from stem cells, including MSCs, embryonic stem cells (ESCs) and other tissue-specific stem cells due to their anticipated immuno-compatibility and documented roles as paracrine mediators of tissue repair. 42 To date, MSC-derived EVs have been shown to enhance proliferation, inhibit apoptosis, decrease inflammation and promote processes such as angiogenesis and cellular reprogramming. 43 In animal studies of myocardial ischaemia, the delivery of MSC-derived EVs has cardioprotective effects, promoting angiogenesis that improved recovery and reduced infarct site in laboratory animals with experimental myocardial infarction. 44 Similarly, delivery of ESC vesicles enriched in miRNA-290-295 was found to promote neovascularisation and enhance cadiomyocyte survival and function following myocardial infarction. 45 In a separate study, cutaneous burns treated with MSC-derived exosomes demonstrated accelerated cell proliferation and re-epithelialisation through the delivery of Wnt4 and downstream β-catenin nuclear translocation. 46 These promising particles have also been shown to promote myogenesis and muscle regeneration in an in vivo model of skeletal muscle injury. Interestingly, in this study, MSC-derived vesicles were found to contain a relatively low concentration of muscle-repair proteins, such as vascular endothelial growth factor (VEGF) and interleukin (IL)-6, with the regenerative effects primarily attributed to the presence of small RNAs, including miRNA-494. 5 Finally, in an in vitro model of inflammatory osteoarthritis, bone marrow–derived and adipose-derived MSC EVs were found to inhibit the action of pro-inflammatory mediators including ILs and tumor necrosis factor-alpha (TNFα)-induced collagenase activity.19,47 This list presents several significant outcomes to date but is by no means comprehensive with additional therapeutic effects observed in a variety of other tissues including the kidneys, liver and nervous system. For a comprehensive overview of the potential of EVs for soft tissue regeneration, we recommend the following articles by Chen et al. 48 and Lamichhane et al. 49
To date, the considerable potential of an EV-based approach to regenerative medicine has perhaps been most comprehensively demonstrated in the skeletal system with prospective applications for enhancing fracture repair and in treatment of degenerative conditions, including osteoporosis. Osteoblast-derived vesicles isolated from actively differentiating cultures are enriched in metalloproteinases conducive to matrix remodelling and calcification. 50 These vesicles have defined roles in regulating osteoclastogenesis through the delivery of the pro-osteoclastic cytokine, receptor activator of nuclear factor κ-B ligand (RANKL). Stimulation of osteoblasts with parathyroid hormone has been shown to further enrich EVs with RANKL. 51 As such, there exists the potential to manipulate culture conditions ex vivo to generate biocompatible and therapeutically enhanced vesicles that are biologically programmed to target osteoclasts for the treatment of osteoporosis. Alternatively, EVs can be modified to modulate osteoclast activity post-isolation through the active loading of anti-osteoclast drugs, such as zoledronate or dasatinib. In addition to osteoclast modulation, we have previously shown that when administered within MSC cultures, osteoblast-derived EVs act as sites for extracellular nucleation of calcium phosphate, enhancing mineralisation when compared to a current gold standard, BMP-2. 33 MSC-derived EVs have been shown to stimulate osteoblastic activity and differentiation through miR-196a, miR-27a, miR206 expression and bone regeneration in vivo. 20 In addition, the injection of MSC-derived exosomes has been shown to promote healing in a murine CD9−/− femur fracture model – a model known to produce reduced levels of exosomes. 52 Evolving the concept of EV-based regenerative medicine even further, a recent study combined MSC-derived vesicles with a scaffold composed of decalcified bone matrix. The regenerative effects of EVs were monitored following subcutaneous implantation in mice and represented the first time that EVs had been delivered in combination with a biological scaffold. Outcomes were positive with enhanced angiogenesis and mineralisation observed in vivo. 53 A subsequent study combined EVs with a poly(lactic-co-glycolic acid) (PLGA) scaffold by immobilising them in polydopamine coating. The scaffold was subsequently implanted in a critical size calvarial defect with sustained release of EVs observed and immunohistochemical evidence of bone regeneration. 54 The aforementioned studies highlight the considerable advances made in EV-focused regenerative medicine. However, these approaches are the first to be published in this novel therapeutic area. As such, there is considerable scope for development and refinement, as we shall now discuss.
Despite the considerable benefits offered by an EV-based approach to regenerative medicine, there are several limitations that need to be overcome before this method can become a clinical reality. For a comprehensive overview of current barriers to translation, the authors recommend a previous publication by Davies and Rafiq. 55 At present, the most limiting factors restricting the advancement of EV therapies can be grouped into the following categories:
Devising efficient methods for the scaling and harvest of EVs;
Identifying markers of therapeutic potency;
Generating a consistent and homogeneous EV product;
Storage, biosafety, biodistribution and pharma-cokinetics;
Positioning regenerative EV therapies within the current regulatory framework.
Manufacturing large numbers of therapeutically viable EVs at scale is essential if we are to meet clinical and commercial demand. This represents a challenging task, particularly when using a primary cell source with a limited window of passage. The task becomes more demanding when we consider the complex media formulations that are typically employed in EV research and the commercial requirement for chemically defined and serum-free systems that maintain EV yield and function. To date, hollow fibre bioreactors have been most applied for this purpose. These reactors apply a fibre-based cartridge with a molecular weight cut off to retain EVs while allowing the free diffusion of nutrients and waste. It has been postulated that the application of hollow fibre reactors could also address issues surrounding the need for serum-free media, since any vesicles contained within the serum fraction will exceed the molecular cut off and, as such, be excluded from the condition medium. 56 We anticipate that other reactor technologies, including stirred tank bioreactors, will also demonstrate utility in EV manufacture given their legacy in the production of numerous biologics, as well as the potential to combine these systems with microcarriers. 57 Furthermore, these reactors can be operated with spin filters to allow for the retention of cells and small particles including EVs. For a comprehensive overview of the application of bioreactors for EV manufacture, we recommend the recent publication by Colao et al. 58 as well as the aforementioned publication by Davies and Rafiq. 55 At present, there is limited knowledge of how the cell culture microenvironment impacts EV production and content. As such, there will be a need to comprehensively evaluate the effects of bioreactor systems designed for cell biomanufacturing on EV production. 59
In addition to manufacturing large concentrations of EVs, it is essential that methods are in place for their efficient isolation. At present, several approaches have been applied to isolate EVs from cell culture media as well as a variety of biofluids. These include differential UC, ultrafiltration, sucrose density gradient sedimentation, size exclusion chromatography, tangential flow filtration, kit-based precipitation and affinity-based selection. A critical overview of several of these techniques for the isolation of EVs from serum can be found in the publication of Buschmann et al. 60 All these methods have advantages and disadvantages and are highly dependent on the availability of specialised laboratory equipment and relevant in-house technical expertise. Perhaps, the most ubiquitously applied, and certainly the one most frequently utilised for the isolation of EVs from hard tissues such as bone, is differential UC. Unlike the other methods listed, UC can be applied to isolate EVs from large volumes of conditioned culture medium. As such, it currently represents the most viable option for the direct isolation of high concentrations required for regenerative applications. However, UC is a specialised, time consuming, laborious and inefficient process. Furthermore, variation can be introduced depending on the type of centrifuge and rotor applied. For example, a shorter sedimentation path length in the fixed angle compared to the swinging bucket rotor will result in a faster sedimentation rate for peripheral vesicles. As a consequence, it is necessary to adjust the duration and force of centrifugation depending on the model and type of centrifuge used. 61 Consequently, different isolation methods will yield EVs of variable heterogeneity, making it difficult to compare results between studies. 55 As interest in therapeutic EVs continues to increase, it is critical that we implement consistent and readily available methods for generating large concentrations of EVs that retain maximal therapeutic efficacy. Recently, a promising method has been developed at the National University of Singapore, which applies centrifugal microfluidics for the label-free isolation of microvesicles. 62 This approach is appealing in that can be applied to efficiently isolate microvesicles over short time periods (typically minutes) using only a standard bench-top centrifuge. However, at present, the technology can only been applied for the isolation of EVs from small sample volumes (µL). As such, further development will be required if this approach is to be used for high-volume samples required for regenerative applications.
Other factors that need to be clarified before EV therapies can begin to be successfully translated related to nomenclature and the comprehensiveness of methods applied for EV characterisation and quantification of batch homogeneity. Current standards have been published in a position statement outlining the minimal experimental criteria required for defining EVs. 11 However, such a definition does not serve to provide a quantitative measurement of therapeutic efficacy, which will no doubt depend on the cell from which the EVs are derived and the purpose of their application. Such a measure can perhaps only be achieved through the identification of specific surface or intra-vesicular markers that can be measured during or post-isolation. In a previous study, we identified a correlation between certain calcium-binding annexin proteins (annexins I, II and VI) and EV-mediated mineralisation in human stem-cell cultures. 39 However, further work will be necessary to determine whether selecting for annexin-enriched EV populations can enhance hard tissue regeneration in vivo. In addition, the application of antibody-based selection methods will pose further challenges when applied for the isolation of large concentrations of EV product at scale. Reflecting on this last comment, it may be advantageous to employ label-free methods that provide non-specific biochemical fingerprints that align with known measures of therapeutic potency. Such methods could include Raman spectroscopy, which has shown successful application for the biochemical profiling of tissue-specific differences in MSC-derived EVs. 63 Once the MoA has been identified and accurate markers of potency defined, we can begin to evaluate critical pre-clinical parameters such as dosage, toxicity and biodistribution. It will also be pertinent to determine how EVs can be stored to maximise biological potency and minimise operational burden in the clinic. There is evidence to indicate sustained viability and functionality of human urinary exosomes when stored at −80°C in the presence of a cocktail of protease inhibitors. 64 This evidence is encouraging given that it may reduce costs associated with more complex cryo-preservation procedures that are a requirement for cell-based therapies. However, studies will be required to determine the effects of long-term storage on EVs isolated from other biofluids and cell sources, including osteoblasts.
Finally, regulation surrounding the application of EV therapeutics is at present unclear. It has been proposed that EV-based therapies will likely fall under the category of biological medicines unless delivering trans-gene products derived from genetically manipulated cells in which case they will fall under the category of Advanced Therapy Medicinal Products (ATMPs). 65 However, regulatory classification will remain challenging until the MoA by which EVs exert their therapeutic effects is established. It is likely that this will vary between applications and may require assessment on a case-by-case basis. For a comprehensive review of current limitations surrounding the advancement of vesicle-based therapeutics in regenerative medicine and beyond, we recommend the article by Gimona et al. 65
Summary and conclusion
EVs offer considerable value as a biological yet acellular approach for driving endogenous tissue regeneration. These complex particles offer significant advantages over growth factor and cell-based approaches and, to date, positive outcomes have been demonstrated in a variety of soft and hard tissues. Within the last 3 years, several groups have shown that these EVs can be combined with a variety of biocompatible scaffolds and implanted in vivo to drive de novo ossification and tissue vascularisation. However, before the considerable potential of EVs can become a clinical reality, inherent variability relating to methods of EV isolation and characterisation need to be standardised.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship and/or publication of this article Owen G Davies was supported by an EPSRC E-TERM Landscape fellowship. Ioannis Azoidis was funded by a School of Chemical Engineering PhD Studentship.