
Abstract
Background and objective:
The term ‘fibroblast’ covers a heterogeneous cell population in idiopathic pulmonary fibrosis (IPF). The fibroblasts are considered as main effector cells, because they promote disease progression by releasing exaggerated amounts of extracellular matrix proteins and modifying cell microenvironment. As IPF-derived human lung fibroblasts (IPF-HLFs) were shown to express higher levels of integrin alpha-5 (ITGA5) than normal derived HLFs (N-HLFs), we explored the importance of ITGA5 to IPF progression.
Methods:
IPF-HLF and N-HLF primary cultures were established. ITGA5 was silenced by specific small interfering RNA (siRNA)s and its effects on cell phenotype (e.g. cell number, size, cell death, migration) and gene expression (e.g. RNA sequencing, quantitative polymerase chain reaction [qPCR], western blot and immunofluorescence) were tested. Specific integrin expression was evaluated in IPF patient formalin-fixed paraffin embedded sections by immunohistochemistry (IHC).
Results:
ITGA5-silencing resulted in reduced IPF-HLF proliferation rates and cell migration (p < 0.05), as well as elevated cell death. transforming growth factor beta (TGF-β) targets (e.g. Fibronectin (FN1), Matrix metalloproteinase 2 (MMP2), TGFB1) were surprisingly elevated following ITGA5 silencing (p < 0.05). N-HLFs, however, were only slightly affected. Interestingly, ITGA5-silenced cells differentiated into myofibroblasts (e.g. elevated alpha-smooth muscle actin [αSMA], collagen1a, large cell size). RNA-sequencing revealed that following differentiation on 3D-Matrigel for 24 h, ITGA5 levels are reduced while integrin alpha-8 (ITGA8) are elevated in IPF-HLFs. This was confirmed in IPF patients, in which ITGA5 was mainly found in fibroblastic foci, while ITGA8 was mostly observed in old fibrous tissue in the same patient.
Conclusions:
ITGA5 expression facilitates a more aggressive proliferative phenotype. Downregulation of this integrin results in myofibroblastic differentiation, which is accompanied by elevated ITGA8. Specific targeting could present a therapeutic benefit.
Introduction
Idiopathic pulmonary fibrosis (IPF) is a specific form of chronic, progressive fibrosing interstitial pneumonia of unknown cause. It is characterized by progressive worsening of lung function, and is associated with poor prognosis.1,2 The pulmonary interstitial fibroblast is presumably the main cell responsible for exaggerated amounts of collagen and other extracellular matrix (ECM) proteins that are being accumulated in the interstitial spaces of lungs during pulmonary fibrosis.3–5 It becomes more and more evident that the term ‘fibroblast’ covers a heterogeneous cell population.6,7 In IPF, pro-fibrotic signals may act on the existing heterogenous fibroblast population to mediate the rise of cell sub-populations resulting in the predominance of the fibrotic phenotype (i.e. myofibroblasts), 8 a phenotype that remains in vitro. 9
Integrins are transmembrane receptors composed of a single alpha and a single beta subunit, 10 which mediate various cell–matrix and cell–cell interactions. In mammals, integrins act primarily as signaling proteins, involved in cell growth, division, survival, differentiation and apoptosis. Integrins facilitate communication between ECM, non-parenchymal cells including inflammatory cells, fibroblasts, and parenchymal cells, and due to these interactions, they are directly involved in the initiation and progression of tissue fibrosis. Therefore, integrins represent highly interesting therapeutic targets. 11
Transforming growth factor beta (TGF-β), which is a major profibrotic cytokine inducing myofibroblast differentiation 12 is known to interact with multiple integrins. TGF-β1 also induces alpha-smooth muscle actin (αSMA) expression and other components of the myofibroblast contractile cytoskeleton, 13 as well as an increased collagen production. 14
In our previous study, we showed that the IPF-derived human lung fibroblasts (HLFs) express significantly higher levels of integrin alpha-5 (ITGA5). 15 Therefore, in the current study we further explored the importance of ITGA5 for IPF progression by silencing its expression in primary HLFs derived from patients with IPF.
Materials and methods
Fibroblast culture
Primary HLFs were isolated from IPF (histologically confirmed) and from control tissue samples (histologically normal lung distant from a resected tumor), obtained at the time of biopsy, as previously described. 15 Fibroblasts were cultured in Dulbecco’s Modified Eagle’s medium (DMEM) supplemented with 20% fetal calf serum (FCS), L-glutamine (2 mM) and antibiotics (Biological Industries, Israel). Cells were passaged once or twice a week without reaching full confluence in order to avoid contact inhibition and maintain an active proliferative state.
RNA silencing
Lipofectamine RNAiMAX (Invitrogen, USA) and four sequences of ITGA5 siRNAs (GeneSolution, Qiagen, Germany) were added to OPTIMEM-I (GIBCO, USA). AllStars Neg. siRNA AF488 served as control. Transfection efficiency was evaluated using flow cytometry (Navious, Beckman Coulter).
Cell count
Cell number and viability were evaluated by a manual count following a Trypan blue staining. Counts were verified by flow cytometry. Cell size measurement was done using Fiji (http://fiji.sc/).
Fibronectin adhesion assay
A total of 5 × 104 cells from each treatment were seeded to 96-well plates coated with fibronectin (FN) (R&D, 10 μg/ml) and allowed to adhere for 60 min. Following washes with phosphate-buffered saline (PBS), the bound cells were counted.
Cell migration
HLFs (5 × 104) were placed in 96-well plates and allowed to adhere for 24 h. Wound closure was monitored immediately after scratching and at 24 h. Areas were measured using the ImageJ (http://rsbweb.nih.gov/ij/).
Western blotting
Western blotting (WB) was performed as previously described. 16 The following antibodies were used: caspase-3, phospho/total Smad3 (CST, USA); proliferating cell nuclear antigen (PCNA) and ITGA8 from Santa Cruz Biotechnology, USA; phospho-focal adhesion kinase (pFAK), total FAK, αSMA, ITGA5, FN and collagen1a from Abcam, USA, Alpha-Tubulin (Sigma USA), ITGAV (Millipore).
Immunohistochemistry
Paraffin sections were deparaffinized and immersed in ethylenediaminetetraacetic acid (EDTA) buffer (pH = 8). Samples were blocked and incubated with primary antibodies overnight. Following washing, slides were incubated with horseradish peroxidase labeled polymer (Zytomed systems, GmbH), and developed with AEC chromogen (ScyTek Laboratories, USA). Isotype-matched control excluded non-specific staining. Images were analyzed using QuPath. 17
Immunofluorescence
Glass 8-well Millicel EZslides (Merck Millipore, Ireland) were covered in fibronectin (10 μg/ml, R&D). Fixated cells were blocked and incubated with primary antibodies [αSMA and active ITGA5 (SNAKA51, Millipore, USA)] overnight. Slides were incubated with secondary fluorescent antibodies (Bethyl Laboratories, USA). 4',6-diamidino-2-phenylindole (DAPI) (1000 ng/ml, Abbott, USA) was used for nuclei staining. Actin fibers were stained using Phalloidin-Fluorescein isothiocyanate (FITC) (Tocris Biosciences). Confocal images were taken with a Nikon Ti2E microscope equipped with a Yokogawa W1 spinning disk system and a Plan Apo 60x oil NA1.4 objective using a 405 nm and 561 nm laser. Images were analyzed using Fiji.
Cell death
Tested using propidium iodide (Sigma Aldrich, USA). Assessment of apoptosis/necrosis was done with AnnexinV-FITC supplemented with Propidium iodide (PI) (MEBCYTO, MBL) by flow cytometry according to manufacturer’s instructions.
RNA extraction and Reverse Transcription (RT) cDNA synthesis
RNA was extracted using the RNeasy kit (Qiagen, Germany). Extracted RNA was converted to cDNA using GeneAmp (Applied Biosystems, USA).
RNA sequencing
RNA libraries were generated using CEL-Seq preparations protocol, and sequenced on Illumina HiSeq2500, 15/50 paired-end run. The reads were mapped to the human genome (ftp://ftp.ensembl.org/pub/release89/fasta/homo_sapiens/dna/Homo_sapiens.GRCh38.dna.primary_assembly.fa.gz) using Tophat2 version 2.1.0 (uses Bowtie2 version 2.2.6, http://www.ncbi.nlm.nih.gov/pubmed/19289445?dopt=Abstract). Only uniquely mapped reads were counted to genes, using ‘HTSeq-count’ package version 0.6.1 with ‘union’ mode (http://www.ncbi.nlm.nih.gov/pubmed/25260700?dopt=Abstract). Normalization and differential expression analyses were conducted using DESeq2 R package version 1.18.1.
Real-time qPCR
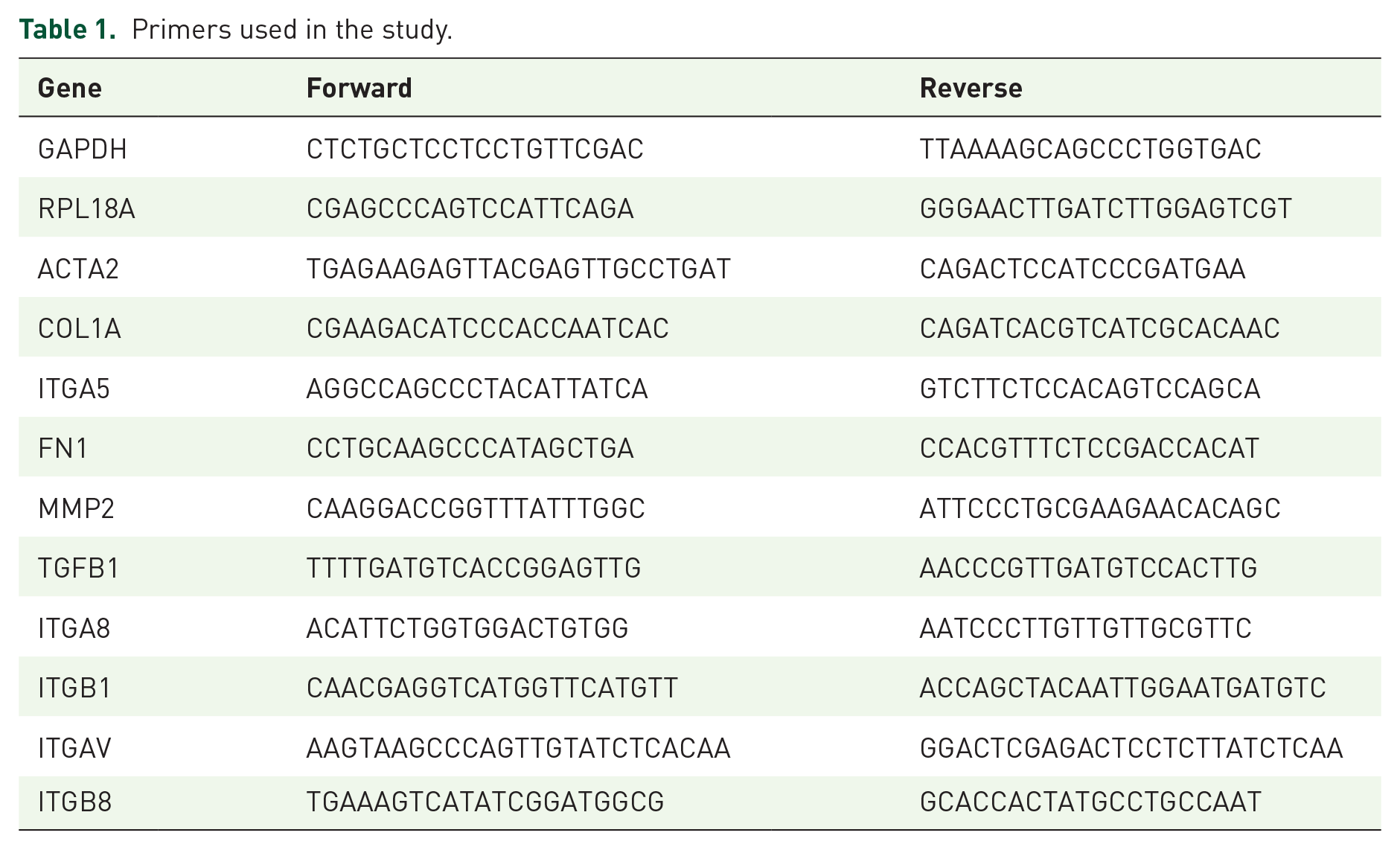
Real-time qPCR was done using Power SYBR Green (Applied Biosystems, USA). The primers (Hylabs, Israel) are listed in Table 1. Glyceraldehyde-3-Phosphate Dehydrogenase (GAPDH) and Ribosomal Protein L18a (RPL18A) served as housekeeping controls.
Primers used in the study.
Statistical analysis
Differences between two cohorts were analyzed with Student’s paired t-tests. Multiple cohorts were analyzed using analysis of variance (ANOVA). p < 0.05 was considered significant.
Ethical approval
The study was approved by the institutional review board (IRB) of Meir Medical Center (MMC) (approval number: 016-16-MMC). Signed informed consent was obtained from all patients.
Results
ITGA5 silencing affects IPF, and not normal tissue derived HLFs
ITGA5 was silenced in IPF and control (N) HLFs. Silencing efficiency of over 80% was observed at the total protein level (Figure 1A and B), as well as at the active form, which was estimated by binding of SNAKA51 antibody to the active conformation of α5β1-integrin (Figure 1C). ITGA5 mRNA levels were also significantly reduced by 66% at 72 h (p < 0.001, n = 9). Moreover, ITGA5 silencing led to reduced attachment to FN (Figure 1D), accompanied by lower levels of phospho-focal adhesion kinase (pFAK) (Figure 1E and F), thus validating functional inhibition. Already at 48 h, a significant reduction in cell migration was observed (Figure 1G), with no change in cell counts or cell death. However, following 72 h, there was a small reduction in IPF-HLF counts (Figure 1H), with an elevation in fragmented caspase 3 (120% increase in siITGA5 versus control, p < 0.05, Figure 1I). These changes were not supported by flow cytometry (30% elevation in early apoptosis in ITGA5 silenced cells versus control, p = 0.6). At 72 h, a significant 30% reduction in PCNA levels and a 55% reduction in cyclin D1 level were also observed in the ITGA5 silenced IPF-HLFs (p < 0.05, Figure 1I). Nevertheless, when silencing was performed on the N-HLFs, no changes in cell counts, cell death, migration or attachment were observed at any time point (48–96 h). No change in PCNA, cyclin D1 or in fragmented caspase-3 levels was found in ITGA5 silenced N-HLFs (Figure 1I). Therefore, the baseline elevated ITGA5 level in the IPF-HLFs may raise their sensitivity to ITGA5 knock-down.
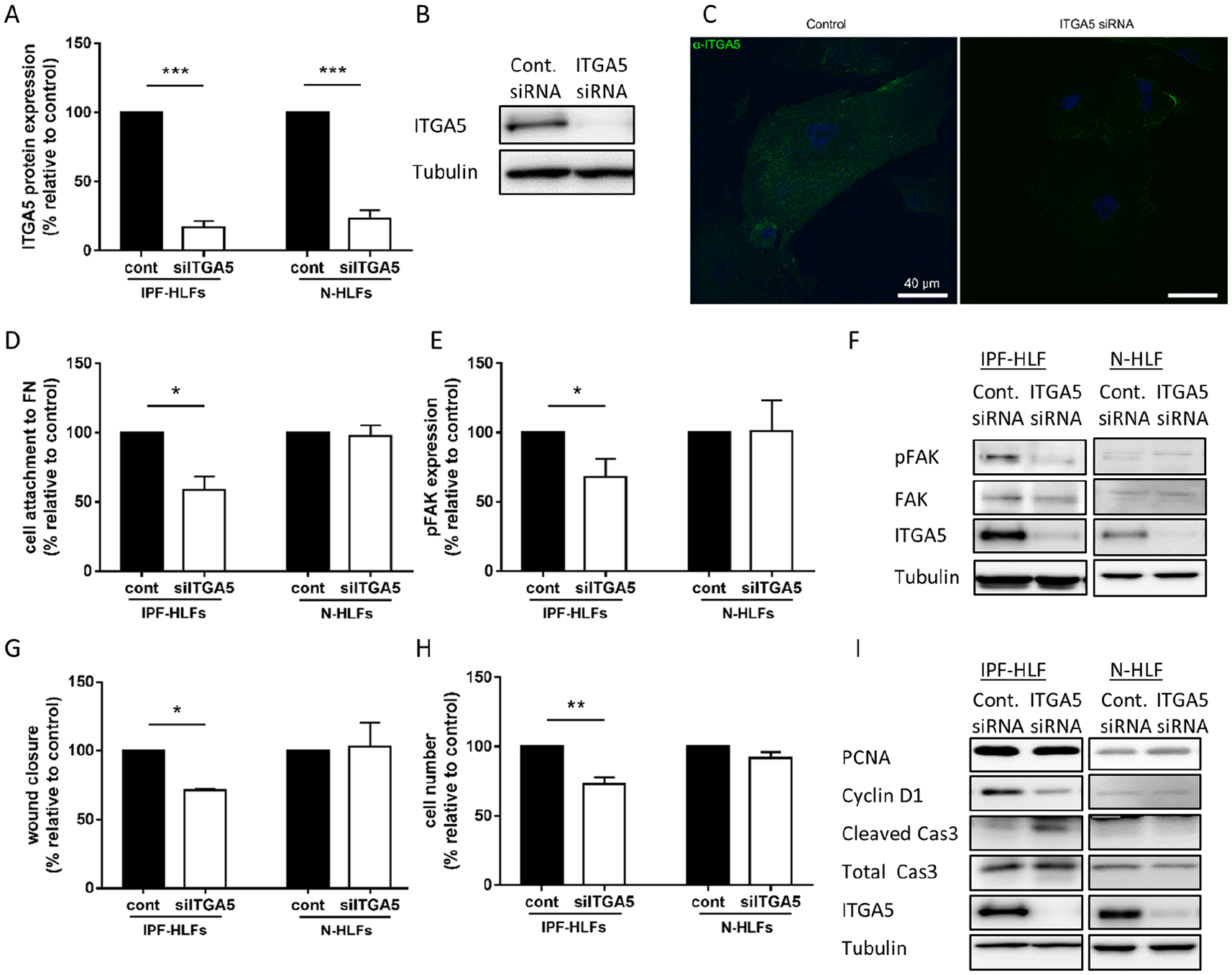
Integrin alpha-5 (ITGA5) silencing affects the idiopathic pulmonary fibrosis (IPF) and not normal tissue derived cells.
ITGA5 silencing leads to fibroblast to myofibroblast transition (FMT)
Collagen1a and αSMA (ACTA2) that are induced by TGF-β and considered as markers of myofibroblast differentiation 18 were elevated following ITGA5 silencing in the IPF-HLFs (p < 0.05, Figure 2A). As the baseline expression of αSMA (ACTA2) is significantly lower in the N-HLFs, an even greater increase of ACTA2 was observed in N-HLFs following ITGA5 silencing (p < 0.05, Figure 2B). Collagen1a (COL1A), however, was only slightly elevated (NS, Figure 2B). These findings were then validated at the protein level (Figure 2C), and using IF for αSMA (Figure 2D). In addition, ITGA5 silencing also induced a larger cell size in both cell types (Figure 2E and F), again implicating cell differentiation towards the myofibroblastic phenotype.

Integrin alpha-5 (ITGA5) silencing leads to fibroblast to myofibroblast differentiation.
Since ITGA5 activation is often associated with increased TGF-β signaling, 19 we explored several known TGF-β targets following the ITGA5 siRNA treatment. Interestingly, all tested targets (e.g. MMP2, SMAD3, FN1 and TGFB1) were significantly elevated in the IPF-HLF cells (p < 0.05, Figure 2G), while N-HLFs were again less affected, as only MMP2 was significantly elevated (Figure 2H). These findings support the differential change in collagen1a elevation between normal and IPF-HLFs, as collagen 1a is also a target of the TGF-β pathway.
An integrin switch may possibly be linked to fibroblast to myofibroblast differentiation
Following the previously mentioned results, we suspected an elevation of another integrin as a compensatory mechanism linked to the fibroblast to myofibroblast transition
Here, we cultured N-HLFs and IPF-HLFs on Matrigel and plastic for 24 h to compare ITGA5 expression. Interestingly, a significant reduction in ITGA5 was only observed in the IPF-HLFs cultured on Matrigel versus plastic (Figure 3A). Moreover, the relative integrin expression of Matrigel cultured N-HLFs and IPF-HLFs was evaluated by RNA-seq (Table 2). The significantly upregulated integrins were ITGA8, ITGB8 and ITGAV in comparison with N-HLFs cultured on the same matrix. Interestingly, the ITGA5 was in fact downregulated in the differentiated IPF-HLFs, with no change in its major coupling unit, the ITGB1. These genes were then validated by qPCR (p < 0.05, Figure 3B). As a next step, we tested the previously mentioned targets in the ITGA5 silenced IPF-HLFs, and found correlating results (p < 0.05, Figure 3C). In fact, when testing these targets in N-HLFs no upregulation in ITGA8 was observed (Figure 3D). At the protein level, the ITGA8 was elevated only in the ITGA5-silenced IPF-HLF cells in comparison with controls (Figure 3E). Thus, ITGA8 presented as a potential candidate, as it also binds the β1 subunit and was linked to IPF. 20 In conclusion, our results suggest a possible switch between the A5 and the A8/AV subunits in the process of FMT in IPF-HLFs (illustrated in Figure 3F).
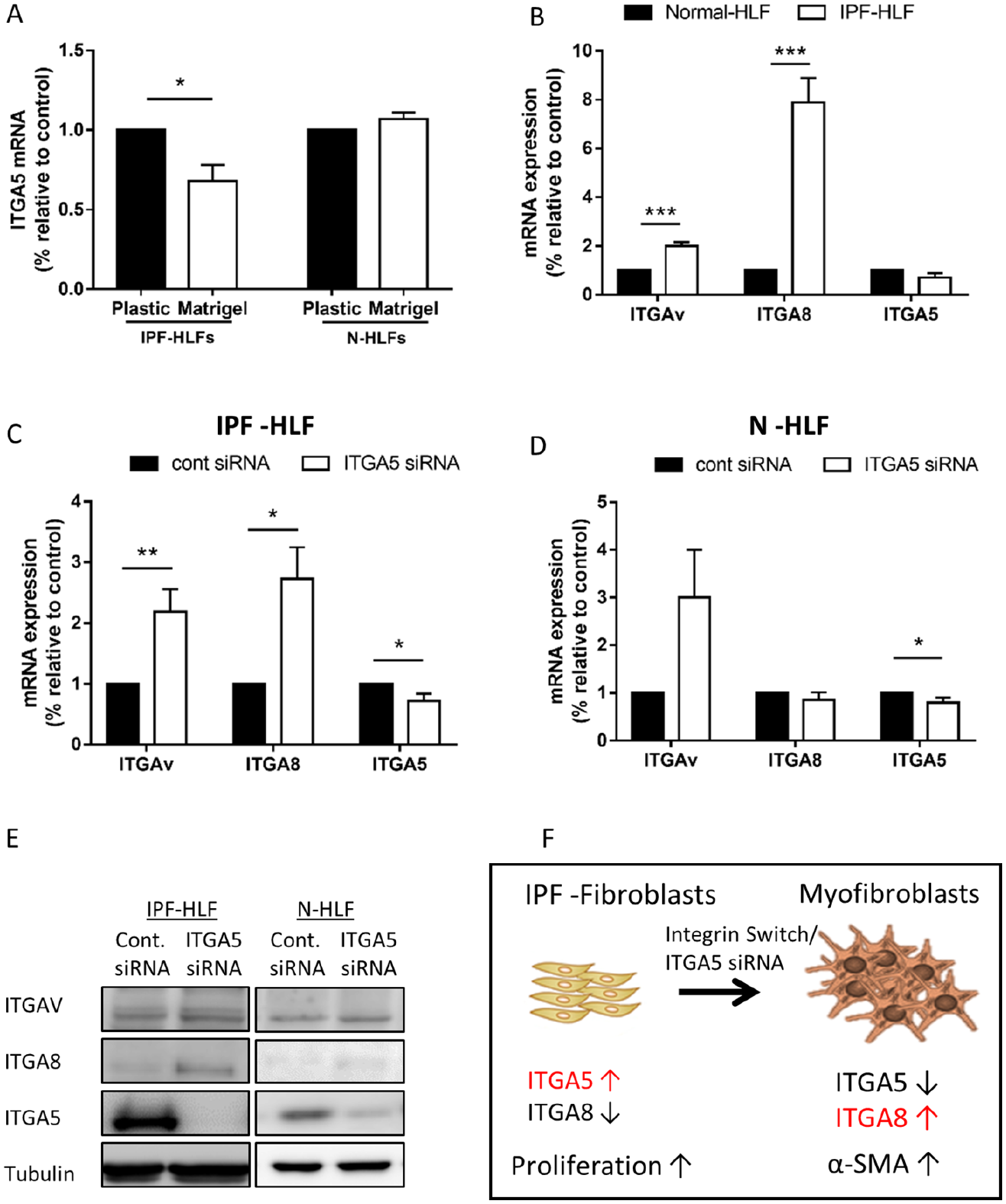
Integrin switch may be linked to cell differentiation.
Integrin expression in IPF versus normal tissue-derived HLFs.

HLFs, human lung fibroblasts; IPF, idiopathic pulmonary fibrosis.
ITGA8 is expressed in old collagenous tissue
In order to confirm our hypothesis regarding a possible ‘integrin switch’, serial sections of IPF patient lung tissues were stained with ITGA5 and ITGA8. Areas of loose fibrotic tissue containing fibroblastic foci (FF) and old fibrous tissue with significant collagen changes were examined in the same patients. As previously shown, 15 ITGA5 was primarily expressed in the FF, while barely detected in the old fibrous tissue (Figure 4B and E). Interestingly, the ITGA8 expression in the FF was relatively weak (Figure 4F), whereas there was a significant staining for ITGA8 in old fibrous collagen areas (Figure 4C). These areas were quantified for the percentage of ITGA5/ITGA8 positive cells to show differential expression (Figure 4G).

Differential integrin expression in idiopathic pulmonary fibrosis (IPF) lung tissue.
Discussion
Integrin–ECM protein interaction results in cytoskeletal changes and signaling that affect various cellular processes, such as cell growth, differentiation, motility and gene expression,21,22 suggesting them as targets for anti-fibrotic therapy. In this work, we silenced the ITGA5 in IPF-derived primary HLFs. We found that specific inhibition of ITGA5 resulted in increased cell death and reduced migration specifically in IPF-HLFs, but not in N-HLFs, suggesting this integrin to be important for IPF progression. In fact, a study by Sen et al. showed that stromal progenitor cells in culture develop into myofibroblasts through proliferative growth during which they express high levels of ITGA5, which is later decreased with cell maturation. 23
The main defining feature of myofibroblasts is the overexpression of αSMA, 24 yet it is expressed at low levels in fibroblasts as well. Specific cell-matrix receptors, such as integrin α5β1, have been identified as regulators of myofibroblastic αSMA expression.25,26 In fact, Franco-Barraza et al. recently showed myofibroblastic conversion, re-localization and upregulation of αSMA 27 using α5β1-integrin-inhibiting mAb16 antibody. 28 We also found that silencing of ITGA5 leads to an elevation in the αSMA expression, as well as cell size, suggesting a possible differentiation process towards a more myofibroblastic phenotype.
The integrin–TGF-β interplay is highlighted in fibrosis, cancer and wound repair. 29 Intriguingly, the induction of integrin expression by TGF-β can be driven by cooperative signaling between the integrin and TGF-β, thereby creating a feed forward loop. 30 The latency-associated protein (LAP)s of TGF-β1 and TGF-β3 contain the Arg-Gly-Asp (RGD) motif that can be potentially bound by five αv-containing integrins, as well as by α5β1 and α8β1. 29 Similar to ITGA5, ITGA8 requires formation of a heterodimer with the β1 unit.20,31 Since all TGF-β targets were unexpectedly upregulated following ITGA5 silencing, we assumed that there might be a compensatory mechanism by another integrin upregulation. Notably, the silencing experiments were performed on plastic, in which the HLFs maintain a relatively undifferentiated phenotype when cultured for short periods of time, which is characterized by elevated ITGA5 and increased proliferation of the IPF-HLFs. 32 Thus, we first cultured normal versus IPF-HLFs on a 3D matrix for 24 h for them to differentiate and performed the RNA-seq analysis. This culture model was selected as it was previously shown that the cell phenotype, especially of fibroblasts, is highly dependent on it spatial arrangement and ECM density.24,33–36 For example, Kim et al. 37 showed that fibroblasts were activated when co-cultured with cancer cells on a 3D matrix. In addition, fibroblasts were shown to differently affect co-cultured cells, depending on the cultures’ 2D/3D setting.36–38 The explanation of those differences may be attributed to the fact that the 3D matrices are generally less rigid, 38 resulting in alternative signaling responses. 39 Therefore, we scanned all integrins following the 3D culture and found that while the ITGA5 in IPF-HLFs was in fact downregulated, the ITGA8 and ITGAV were significantly elevated. These results were then confirmed by western blot and qPCR that showed even higher levels of ITGA8 compared with ITGAV.
Integrin α8β1 is upregulated in conjunction with myofibroblast differentiation in pulmonary, hepatic, and cardiac fibrosis.20,40,41 In cardiac fibroblasts, α8β1 is induced by TGF-β and upregulated on myofibroblasts. 42 In the lung, ITGA8 expression is restricted to interstitial stromal cells, 43 and it was shown to be increased following bleomycin-induced fibrosis. 20 Moreover, α8β1 and αSMA are often co-localized during pulmonary fibrosis. Levine et al. showed that within areas of more advanced fibrosis, areas of dense α8-positive cellular infiltration were observed. 20 These results support our findings of increased ITGA8 in areas of dense fibrosis, while ITGA5 was limited to areas of ‘young’ loose fibrosis, localizing mainly in the FF, where cells are expressing relatively low levels of αSMA as well.
ITGA8 knockout animals experience early postnatal mortality due to defects in nephrogenesis, which limits their utility in mice models. 40 Hung et al. attempted to overcome this issue by knocking out the ITGA8 only in a subpopulation of PDGFRβ+ isolated stromal cells. 43 They showed that ITGA8 silencing led to an elevation in COL1A levels following TGF-β treatment. However, in the bleomycin-induced fibrosis model they did not find significant results. This could be due to their restricted cell selection.
Our study includes several limitations. The use of primary cell cultures from different patients and control donors led to high variability in the effects and, therefore, all results were normalized. In addition, the relatively short culture period of primary cells does not enable wide parallel comparisons between different donor cell lines, especially concerning cell phenotype. Thus, we could not compare several IPF-HLF lines with their relative ITGA5 expression and compare their response with the siRNA treatment. Finally, the mechanism by which ITGA5 silencing specifically affects IPF, but not normal HLFs was not fully elucidated. These findings warrant future research.
It is increasingly evident that the term ‘fibroblast’ covers a heterogeneous cell population.6,7 Therefore, only those HLFs that can generate myofibroblasts should be targeted, 33 as recently supported by studies on skin and heart fibrosis.44,45 Here, we showed at least two possible HLF populations with distinct characteristics. The first expressing high ITGA5 with high proliferation rates, which is located mainly at the FF, while the other – overexpressing ITGA8 with elevated αSMA and collagen 1a, with a more senescent phenotype that mostly resides in advanced stage fibrotic tissue.
In conclusion, ITGA5 silencing leads to elevated ITGA8 levels and myofibroblast differentiation. These results suggest the existence of several fibroblast populations with distinct characteristics during IPF progression and highlight both ITGA5 and ITGA8 as possible targets for IPF treatment. Further study is required to confirm these findings in other experimental systems.
Footnotes
Acknowledgements
The authors thank Ms Tatiana Epstein for the English editing.
Author contributions
GES drafted the manuscript, designed the experiments and analyzed the results, EB and BW performed the experiments and analyzed the results, EE and HG revised it critically for important intellectual content, while DS contributed to the conception and design, drafting the manuscript for important intellectual content and revised the final version.
Conflict of interest statement
The authors declare that there is no conflict of interest.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.