
Abstract
Objective
Hypertonic injection of dextrose is an alternative treatment for reducing pain and increasing function in patients with knee osteoarthritis (OA). Dextrose prolotherapy (DP) is hypothesized to induce localized inflammation, leading to proliferation of cells in the joint space. This in vitro study explores how exposure to therapeutic doses of hypertonic dextrose affects fibroblast viability and proliferation, either directly or indirectly through exposure to secreted factors by dextrose-treated cells.
Design
MRC-5 fibroblasts exposed for 15 to 120 minutes to dextrose solutions at concentrations of 5%, 10%, 15%, 20%, or 25% were compared to a media-only control. Metabolic activity was measured by the XTT assay as an indicator of cell viability and proliferation.
Results
Fibroblasts exposed for any length of time at the highest concentration of dextrose (25%) or lower concentrations (10-20%) for longer durations exhibited significant reductions in cell viability compared to media controls. However, fibroblasts exposed to higher concentrations of dextrose (15-25%) for shorter durations (30-60 min) or lower concentrations (10%) for longer durations (120 min) exhibit an increased proliferative effect 48 hours after the initial experiment. Nascent fibroblasts exposed to supernatant fluid from cells directly treated with dextrose did not have a negative impact on cell viability compared to the media control.
Conclusions
These results suggest dextrose concentrations used for prolotherapy may stimulate proliferative responses in fibroblasts in support of theorized mechanisms of DP.
Keywords
Introduction
Osteoarthritis (OA) impacts millions of people a year and is the most prevalent joint disorder. 1 In OA, the cartilage within the joint degrades over time, causing a change in the structure of the underlying bone which leads to pain and reduced function. Tissue repair and healing is complicated by a limited vascular supply, inflammation from chronic use, and age-related changes. Despite the frequency of OA in the population, there is no established, standardized treatment to slow or reverse the progression of the disease. Clinical presentation of OA can vary considerably and range from minor (Stage 1), mild (Stage 2), moderate (Stage 3) to severe (Stage 4). 2 Treatments to manage the symptoms of mild to severe OA also vary, depending on the stage of OA and other host factors. These include lifestyle changes, physical therapy, and/or oral medications (e.g., nonsteroidal anti-inflammatory drugs [NSAIDs]) for less advanced stages of OA; however, as OA progresses, these may lose effectiveness, requiring opioids or eventually surgery (e.g., total joint replacement), which present increased cost, potentially significant adverse effects, and/or limited use in certain patients due to comorbidities and safety concerns.3,4
Prolotherapy, which involves injection of an irritant solution into ligaments, tendons, or joint spaces, is an alternative therapy for various musculoskeletal disorders including OA. In one of the earlier records of prolotherapy, Hackett 5 demonstrated ligaments could be strengthened by injection with a proliferative agent—hypertonic dextrose. Injection of dextrose, one of the more common agents used, has since been shown to reduce pain, increase joint function and mobility, and even slow disease progression, particularly for the mild-to-moderate stages of OA.6 -8 Although clinical outcomes from dextrose prolotherapy (DP) have demonstrated its safety and efficacy, the mechanism of action—theorized to induce mild inflammation that stimulates healing—is less understood.6,8,9
Many proliferative agents have been used since then; however, hypertonic dextrose solutions (>5% dextrose) of 12.5% and 25% are particularly popular due to dextrose’s relatively low cost, minimal side effects, and strong safety profile. 8 Although more robust clinical studies are needed, clinical use of DP has shown promising results in treating knee OA.7,9 -11 A pilot study with 7 participants showed reduced pain, improved walking speed, and a decrease in the Western Ontario and McMaster Universities Arthritis Index (WOMAC) scores. 7 WOMAC is a standardized index to evaluate patients with knee and hip OA; lower scores are indicative of reduced pain and improved function. 12 Another small study found individuals with knee OA had increased cartilage thickness and decreased WOMAC scores. 10 Multiple studies investigating the use of DP for OA in a variety of joints have shown high patient satisfaction after treatment.7,10,11 In a systematic review evaluating 33 studies, including randomized control trials, case series, and a case control study, DP performed better than injection controls. 9 This comprehensive analysis indicated there is evidence to support treatment of tendinopathies, knee OA, finger OA, and spinal/pelvic ligament dysfunction with DP. 9 However, clinical trials investigating DP have varied greatly in the injection procedure, resulting in varied treatment outcomes and the lack of a standardized protocol, and many studies lack a direct comparison to other therapeutic options such as lifestyle changes and analgesics. 6 Therefore, the 2019 American College of Rheumatology/Arthritis Foundation Guideline for the Management of OA of the Hand, Hip, and Knee conditionally recommends against prolotherapy of the hip and knee, pending more research. 3 As a result of this recommendation, DP is not typically covered despite promising results and low cost, highlighting the need for further investigation into the mechanism of DP and additional clinical studies.
While clinical research is limited, cell-based studies investigating the mechanism of prolotherapy are even more scarce. Currently, prolotherapy is theorized to induce an initial inflammatory response surrounding the injection site, stimulating cellular proliferation and tissue repair, ultimately resulting in improved strength, stability, and reduced pain. Dextrose is thought to work as an irritant on a variety of cells including fibroblasts and chondrocytes, stimulating them to release factors such as growth factors, cytokines, and other factors that may promote proliferative responses and tissue healing.6,13 -15 Fibroblasts secrete important components of the extracellular matrix, growth factors, and other factors such as cytokines associated with tissue repair.16,17 Fibroblasts also possess progenitor cell functions capable of supporting homeostasis and potentially remodeling. 17 Given the availability and frequent use of fibroblasts for in vitro experiments as well as their role in tissue repair, modulation of other cells, and collagen deposition, fibroblasts provide a good model for investigating cellular responses within the joint space.
In cellular models, dextrose has demonstrated the ability to enhance chondrogenesis or stimulate the formation of cartilage through various ways. Aggrecans are components of cartilage that resist compressive forces; in early stages of OA, aggrecan degradation can occur. Dextrose may induce aggrecan expression, thereby counteracting this degradation. 14 Hypertonic concentrations of dextrose have also been associated with increased metabolic activity and proliferation of chondrocytes along with increased collagen deposition. 15 Another study also showed that dextrose can suppress cytokines and their signaling pathways associated with the degradation of cartilage in OA. 18 However, tenocytes—cells that secrete the extracellular matrix within tendons—showed a decrease in metabolic activity and increase in pro-inflammatory cytokines following dextrose exposure. 19 Fibroblast studies have shown similar results where hypertonic concentrations of dextrose may decrease cell viability. 20 The limited number of cell-based studies and varied results prompted us to further explore the fibroblast response to dextrose. In this study, we explore the impact of varying concentrations of therapeutic doses of dextrose exposure on the cell viability and proliferative responses of fibroblasts. This was completed by evaluating the metabolic activity of fibroblasts exposed directly to dextrose immediately following exposure and then 48 hours later to determine the direct effects of dextrose on cell viability and proliferation, respectively. We also investigate how secreted factors from directly exposed fibroblasts impact metabolic activity of nascent fibroblasts (indirect effects). Therefore, we examine both the direct and indirect effects of dextrose on fibroblasts, across a range of exposure times and concentrations used clinically for DP.
Methods
Cell Culture Reagents
The MRC-5 cells, a human lung fibroblast cell line (ATCC; cat# CCL-171), were cultured in complete medium made from Eagle’s Minimum Essential Medium (ATCC; cat# 3002003) supplemented with 10% fetal bovine serum (Atlanta Biologicals; cat# S11150), 1% penicillin/streptomycin (ATCC; cat# 30-2300), and 2 mM L-glutamine (ATCC; cat# 30-2214) according to the protocol for subculture recommended by ATCC. Cells were maintained in 150 cm2 tissue culture flasks at 37°C supplemented with 5% CO2.
Preparation of Control and Dextrose Solutions
Dextrose solutions (5%, 10%, 15%, 20%, and 25%), based on relevant clinical concentrations used therapeutically in prolotherapy,6,9,10,21 were prepared using granulated dextrose (Acros Organics; cat# 170080010) dissolved in complete medium (as described above), maintaining sterile conditions. The complete medium without additional dextrose was used as the control condition. All solutions and controls were plated on a 6-well plate and incubated at 37°C, 5% CO2 overnight to allow pH equilibration prior to use in cellular assays.
Assays Measuring Cellular Metabolic Activity
The Invitrogen CyQUANT XTT Cell Viability Assay (Thermo Fisher Scientific; cat#: X12223) was used to measure cellular metabolic activity as an indicator of cellular viability. The XTT assay protocol was followed per manufacturer’s instructions to measure changes in metabolic activity across a range of incubation times using media without dextrose as controls. Measurements of optical density (OD) via absorbance at 450 and 660 nm were obtained using the Berthold Crocodile 5-in-One ELISA Mini Workstation spectrophotometer, correcting for background absorbance.
In vitro Experiments to Measure Fibroblast Response to Dextrose Treatments
To determine the immediate effect of directly exposing fibroblasts to dextrose concentrations used therapeutically for prolotherapy, cells were seeded at 20,000 cells per well in a 96-well plate. The cells were grown in 100 µL complete medium overnight to allow adherence to the plate. The next day the medium was removed from the cells and replaced with 100 µL of either complete medium (control) or a range (5%, 10%, 15%, 20%, and 25%) dextrose in complete medium. Exposure times of 15, 30, 60, and 120 minutes were evaluated for each dextrose concentration or control. After the respective times had elapsed, treatments were removed and replaced with 100 µL fresh complete medium and the XTT assay performed immediately. Samples were run in duplicate from 5 independent experiments.
To investigate the proliferative potential of directly exposing fibroblasts to dextrose concentrations, fibroblasts were exposed to varying concentrations of dextrose or control for 15 to 120 minutes as described above. Once the exposure time was reached, the dextrose treatment or control was removed and replaced with 100 µL fresh complete medium; then, the fibroblasts were incubated for an additional 48 hours at 37°C, 5% CO2. After 48 hours had elapsed, the XTT assay was performed. Samples were run in triplicate from 5 independent experiments. Values taken immediately following dextrose exposure were compared to values taken 48 hours following dextrose exposure. These values were normalized against the media control corresponding to the incubation time used for each condition to determine the percent change in metabolic activity as an indicator of proliferative responses by these cells.
To examine the indirect effects of dextrose exposure (i.e., how the supernatant fluid taken from directly dextrose-treated fibroblasts impacts cells that were not directly exposed to dextrose), fibroblasts were directly exposed to varying concentrations of dextrose or control for 15 to 120 minutes as described above. Once the exposure time was reached, the dextrose treatments or control were removed and replaced with fresh complete media; then, the directly treated cells were incubated for another 8 hours at 37°C, 5% CO2 to allow for any secreted factors in response to the dextrose treatment to be produced and accumulate within the supernatant fluid. After 8 hours, the supernatant fluid from the directly treated fibroblasts was collected, centrifuged at 500 × g for 10 minutes at 4°C to remove cellular debris, and then incubated at 37°C, 5% CO2 for pH equilibration prior to being applied to nascent fibroblasts, cells which had never been exposed to dextrose treatments. The nascent fibroblasts were plated at 20,000 cells per well on a 96-well plate in complete medium overnight at 37°C, 5% CO2. The next day, the complete medium was removed and replaced with 100 µL of the collected supernatant fluid from the dextrose-treated fibroblasts. After incubating for 48 hours at 37°C, 5% CO2, the XTT assay was performed on the nascent fibroblasts to investigate the indirect effects of dextrose exposure. Samples were run in triplicate from 5 independent experiments.
Statistical Analysis
Absorbance values of the repeated measures from each experiment were averaged to create a single value for each treatment and timepoint for each experiment. A 1-way analysis of variance (ANOVA) was used to compare each treatment at each timepoint to a matched media-only control. To account for type I error from multiple comparisons, P-values were adjusted using “P.adjust” with the False Discovery Rate (FDR) correction method, where significance is P < 0.05. The F-statistic, confidence interval (CI) values for the coefficient, and P-values are reported in
Summary Statistics for Fibroblasts Directly and Indirectly Exposed to Dextrose.
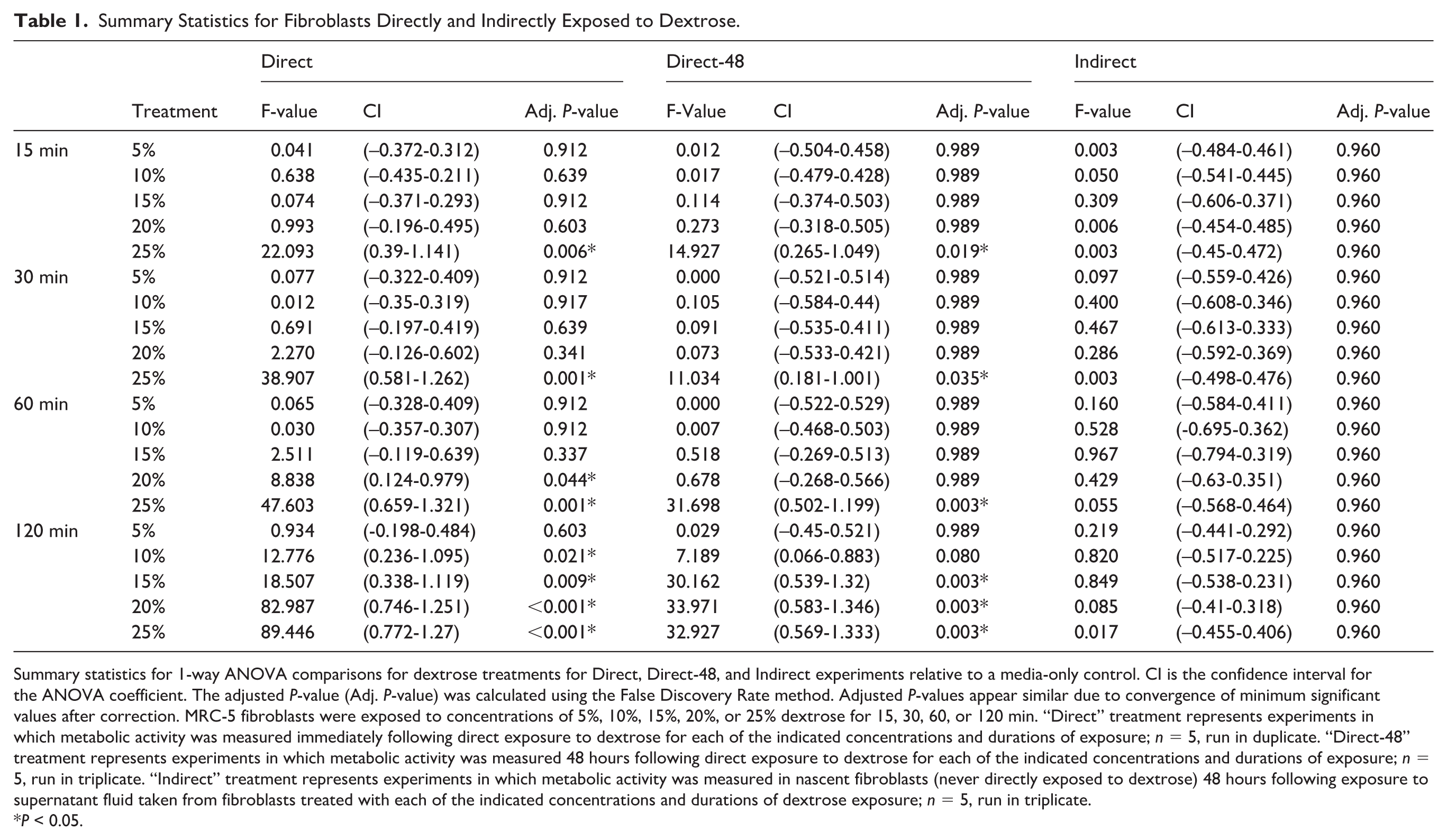
Summary statistics for 1-way ANOVA comparisons for dextrose treatments for Direct, Direct-48, and Indirect experiments relative to a media-only control. CI is the confidence interval for the ANOVA coefficient. The adjusted P-value (Adj. P-value) was calculated using the False Discovery Rate method. Adjusted P-values appear similar due to convergence of minimum significant values after correction. MRC-5 fibroblasts were exposed to concentrations of 5%, 10%, 15%, 20%, or 25% dextrose for 15, 30, 60, or 120 min. “Direct” treatment represents experiments in which metabolic activity was measured immediately following direct exposure to dextrose for each of the indicated concentrations and durations of exposure; n = 5, run in duplicate. “Direct-48” treatment represents experiments in which metabolic activity was measured 48 hours following direct exposure to dextrose for each of the indicated concentrations and durations of exposure; n = 5, run in triplicate. “Indirect” treatment represents experiments in which metabolic activity was measured in nascent fibroblasts (never directly exposed to dextrose) 48 hours following exposure to supernatant fluid taken from fibroblasts treated with each of the indicated concentrations and durations of dextrose exposure; n = 5, run in triplicate.*P < 0.05.
Results
Overall, when examining the viability of fibroblasts immediately after direct exposure to dextrose, we observed reduced cell viability at higher concentrations of dextrose or when cells were exposed to dextrose for longer periods of time. The highest concentration of dextrose (25%) reduced cell viability the most significantly regardless of the duration of exposure (
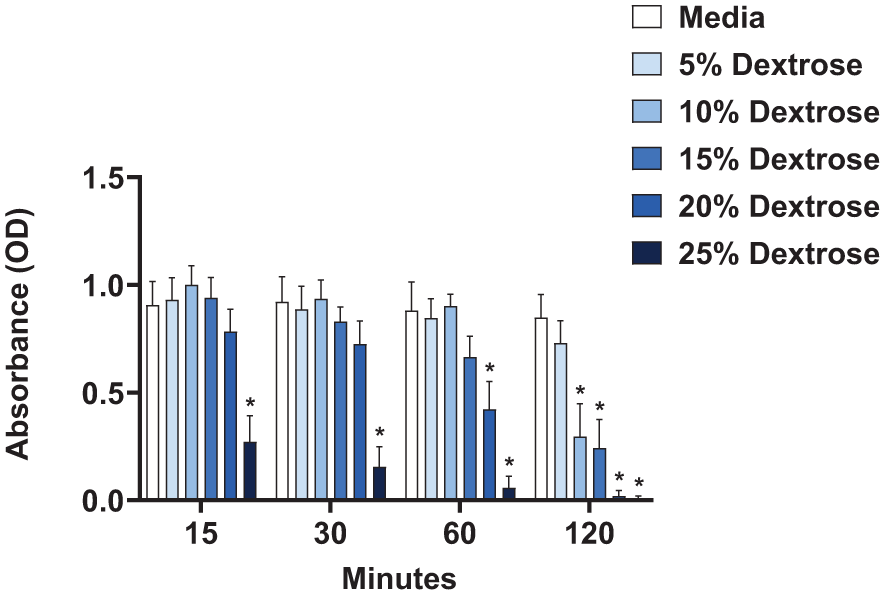
Direct effect of dextrose exposure on fibroblast metabolic activity immediately following treatment. XTT assay, displaying optical density (OD) readings of fibroblasts treated with 5%, 10%, 15%, 20%, and 25% dextrose compared to media control for 15, 30, 60, and 120 minutes of exposure, run in duplicate, n = 5, error bars represent SEM, *P<0.05.
Next, we investigated the recovery potential of cells directly exposed to dextrose at these various treatment conditions. After each dextrose exposure time was reached (5-120 min), the dextrose treatments were removed and replaced with fresh complete media; then, the cells were incubated for another 48 hours to grow and replicate. Fibroblasts exposed to the highest concentration of dextrose (25%) had reduced cell viability across all durations of initial exposure (5-120 min) (
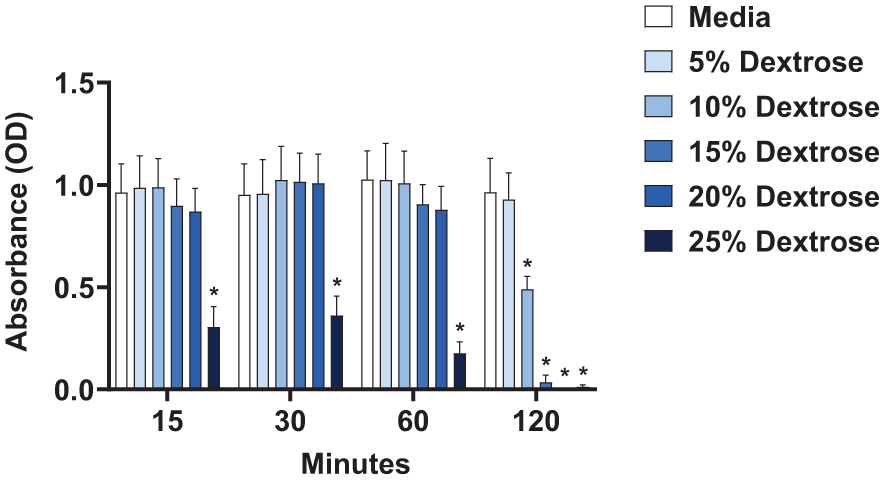
Direct effect of dextrose exposure on fibroblast metabolic activity 48 hours after treatment. After fibroblasts were treated with 5%, 10%, 15%, 20%, and 25% dextrose compared to media control for 15, 30, 60, and 120 minutes of exposure, the supernatant fluid was replaced with fresh media and the cells were maintained at 37°C, 5% CO2 for 48 hours prior to running the XTT assay, displaying optical density (OD) readings, run in triplicate, n = 5, error bars represent SEM, *P < 0.05.
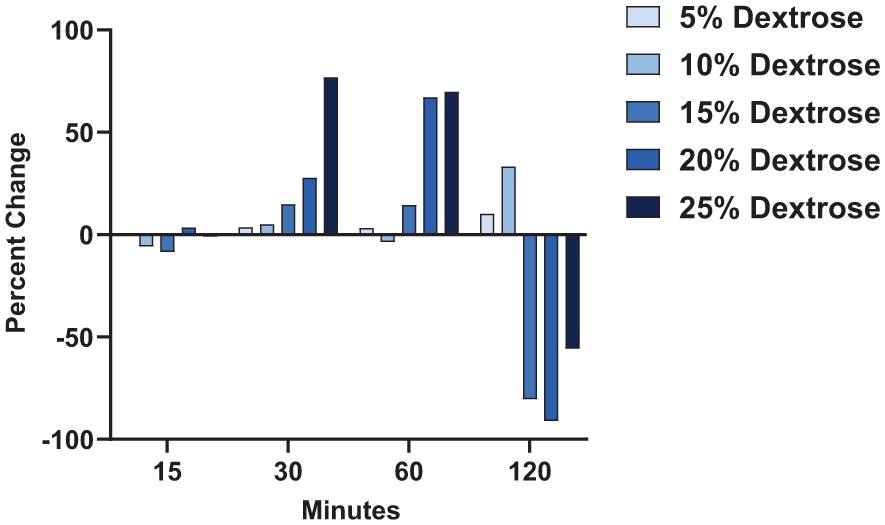
Percent change in fibroblast metabolic activity comparing immediately following dextrose treatment to 48 hours following treatment. MRC-5 fibroblasts were exposed to concentrations of 5%, 10%, 15%, 20%, or 25% dextrose for 15, 30, 60, or 120 min. Absorbance was measured by XTT assay immediately following these treatment conditions and then also 48 hours following each of these treatment conditions. Average absorbance values using the XTT assay were calculated for each type of treatment: (1) immediately following direct treatment with dextrose and (2) 48 hours following direct treatment. The percent change was calculated as a ratio of the 48-hour average absorbance value minus the immediate average absorbance value divided by the average absorbance value immediately following treatment for each matched dextrose concentration and exposure time. These values were normalized to the corresponding media control for each duration of dextrose treatment (15, 30, 60, or 120 min). Averaged values represent 5 independent experiments.
In the final set of experiments, we evaluated the effect of supernatant fluid, collected from dextrose-treated fibroblasts, on the viability of nascent fibroblasts (cells that had not been treated with dextrose). After treatment with 5% to 25% dextrose for 15 to 120 minutes, the dextrose-containing media was replaced with complete media and the fibroblasts were incubated for 8 hours to allow for the accumulation of secreted factors within the supernatant fluid. Then, the 8-hour supernatant fluid from these directly treated fibroblasts was applied to nascent fibroblasts (those with no exposure to dextrose). After 48 hours of exposure to this supernatant fluid, metabolic activity was measured in the nascent fibroblasts. In contrast to direct exposure to dextrose, supernatant fluid from dextrose-treated cells did not reduce cell viability in nascent fibroblasts at any concentration of dextrose or any length of exposure tested (
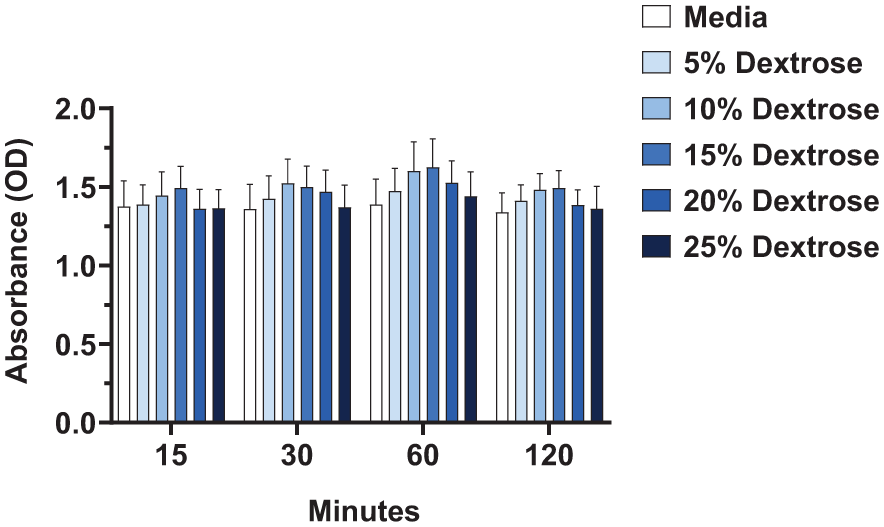
Indirect effect of dextrose exposure on the metabolic activity of nascent fibroblasts 48 hours after treatment. After fibroblasts were treated with 5%, 10%, 15%, 20%, and 25% dextrose compared to media control for 15, 30, 60, and 120 minutes of exposure, the supernatant fluid was replaced with fresh media and the cells were maintained at 37°C, 5% CO2 for 8 hours prior to collecting the supernatant fluid. Nascent fibroblasts (not exposed to dextrose) were then incubated with this supernatant fluid from dextrose-treated cells for 48 hours. XTT assay, displaying optical density (OD) readings of nascent fibroblasts 48 hours after being exposed to supernatant fluid taken from fibroblasts directly treated with dextrose, run in triplicate, n = 5, error bars represent SEM. *P < 0.05.
Discussion
OA impacts 1 in 7 adults in the United States 23 and currently has no cure. DP appears to improve pain and function in OA patients more than placebo in randomized controlled trials, but there is no conclusive evidence that it slows structural OA progression or promotes true joint healing. More high-quality mechanistic and imaging-based studies are needed. DP has been suggested to cause an initial inflammatory response that is followed by cellular proliferation and the production of growth factors.9,21,24,25 Using the in vitro fibroblast model, we found exposure to higher concentrations of dextrose for longer periods of time negatively impacted cell viability immediately following treatment. Comparing percent change in metabolic activity immediately following these exposures compared to 48 hours later, we observed exposure for 120 minutes had a sustained damaging effect with starting concentrations of 15% to 25% dextrose but not 5% to 10% dextrose, indicating that exposure at higher concentrations for long durations of time may be detrimental to fibroblasts. However, cells exposed to 20% to 25% dextrose for 30 to 60 minutes demonstrated notable increases in their metabolic activity at 48 hours despite initial damaging effects observed immediate post exposure. These results indicate that even though higher concentrations of dextrose may initially reduce cell viability, the cells remaining may benefit from an enhanced proliferative response if exposed for an hour or less in support of current proposed hypotheses for the mechanism of action by DP. The supernatant fluid produced from directly treated fibroblasts (8 hr post-treatment) neither enhanced nor reduced viability of nascent fibroblasts.
Findings from the present study provide support for the proposed mechanism of DP and suggest exposure to hypertonic solutions of dextrose can cause an initially damaging response followed by a proliferative response.9,21,24,25 Previous research has also shown that direct exposure to dextrose solutions can decrease fibroblast cell viability. 19 Interestingly, fibroblasts introduced to dextrose concentrations <25% for an hour or less, were able to fully recover as their metabolic activity was comparable to the media control at 48 hours post-treatment. Although initially there is cellular damage, this is followed by recovery and proliferation of cells to baseline levels even from those cells exposed to relatively high concentrations and for up to an hour. When adjusting for the initially damaging response, and therefore fewer starting number of cells immediately following dextrose treatment across various conditions, at 48 hours post-treatment, 20% to 25% dextrose for 30 to 60 minutes demonstrates the greatest increase in proliferative responses. A similar response has been reported in chondrocytes after exposure to hypertonic solutions of dextrose. 15 Furthermore, there may be an optimal concentration and duration of exposure that stimulates proliferative responses in fibroblasts; based on our results, we found starting concentrations of 20% to 25% dextrose exposure for 30 to 60 minutes to demonstrate the greatest increase in cellular activity indicative of proliferating cells. Although the supernatant fluid from directly exposed fibroblasts did not significantly impact the metabolic activity of nascent fibroblasts in our experiments, this still represents a potential area of the mechanism of DP worth investigating further. Overall, our research supports previous findings in similar studies while also providing further support of the currently proposed mechanism of DP. Our work highlights varying responses to dextrose depending on the concentration, duration, and degree of exposure.
This study tested a large range of dextrose concentrations (5-25%) in an effort to match the range of dextrose concentrations that are used clinically across this range.6,9,10,21 One of the challenges of the study and the clinical use of DP is knowing the half-life of intra-articular injections of dextrose solutions. However, based on its half-life in serum, we would expect the half-life within the joint space to be approximately 15 minutes or less but not more than 2 hours, as it is readily metabolized and dispersed.26,27 In support of this theory, a recent study measuring intra-articular injection of dextrose within human knees, demonstrated a significant drop in dextrose concentrations within the first 10 to 30 minutes, 28 supporting our use of short exposure times to dextrose in vitro. It is worth noting that normal metabolic processes of cells both under experimental conditions and in vivo would also contribute to decreasing concentrations of dextrose shortly after exposure; therefore, while we used initially 5% to 25% dextrose solutions, we do not expect that these concentrations were maintained over the duration of the exposure times but likely dropped as time progressed. In the clinical setting, therapeutic concentrations ranging from 10% to 25% are most commonly used—hence, their usage in this cellular model.6,9,10,21
The volume of the synovial fluid within the joint space is another important consideration. Based on other reports, the range of fluid within a normal joint varies significantly but can be greater with severe inflammation and pain.29,30 In clinical practice, the volume of intra-articular knee injections varies, but many clinicians utilize 5 to 10 mL.31 -34 Therefore, at the highest concentration used (25%) in this study and also clinically for DP, we would expect the concentration to quickly drop to <25% after injection as the dextrose solution mixes with the synovial fluid.6,21 Depending on the dilution factor upon injection which will be invariably influenced by patient factors (i.e., the size of the individual and inflammatory state of the knee joint) and the actual amount injected, the initial concentration of dextrose may be expected to minimally decrease or could be reduced by more than half within minutes of the injection. Hence, we used varying concentrations of hypertonic solutions of dextrose with the highest concentration at 25%; however, it is unlikely that cells would be exposed to this concentration for very long given the factors discussed above and a recent study describing the pharmacokinetics within the human knee joint. 28
While our findings support future work in this field and provide further insight into the mechanism of DP, there were some limitations to our study. The use of a cellular model and fibroblast cell line does not adequately represent the heterogeneity and complexity of the cells and tissues within the knee joint and biological system. However, it is a good model to investigate the response of individual components within the joint space to investigate the mechanism of DP at the cellular level. Although human cells were used, we utilized an immortalized lung fibroblast cell line commonly used in cellular studies; these cells have a greater proliferative profile than primary cells, so the response of primary cells and those within the joint space may be different. As discussed earlier, although we tried to account for the possible range of concentrations and exposure times to hypertonic concentrations of dextrose used for DP, the synovial fluid within individuals and amount injected in the clinical setting varies. The study also only evaluated the effects of commonly used therapeutic concentrations of dextrose solutions; however, clinically, dextrose solutions are often mixed with various concentrations of lidocaine. It is unclear how the combination of different concentrations of dextrose mixed with lidocaine may impact cells found within the joint space.
Further research is needed to fully understand the mechanism of DP and how dextrose exposure impacts the function and proliferative responses of other cell types found within the joint. It would also be beneficial to determine the half-life of hypertonic dextrose solutions at different therapeutic concentrations following intra-articular injections and determine the optimal concentrations for clinical use. This work and previous studies highlight a need to thoroughly evaluate how dextrose exposure impacts the production of secreted factors that regulate inflammatory and proliferative responses.
Conclusion
Cell viability and proliferative responses of fibroblasts varied depending on the concentration and duration of dextrose exposure. Immediately following exposure, higher concentrations of dextrose (25%) at all time points or lower concentrations (10-20%) at longer time points reduced cell viability in fibroblasts. This pattern was similar 48 hours after the experiment, with no significant differences in metabolic activity, which suggests that fibroblasts exposed to higher concentrations of dextrose for longer periods of time were not able to recover fully to baseline levels. In contrast, fibroblasts exposed to higher concentrations of dextrose (20-25%) for shorter durations (30-60 min) and lower concentrations (10%) for longer durations (120 min) exhibit significant proliferative responses 48 hours after initial dextrose treatment. These results suggest dextrose concentrations used for prolotherapy may stimulate proliferative responses in fibroblasts in support of theorized mechanisms of DP.
Supplemental Material
sj-docx-1-car-10.1177_19476035251408601 – Supplemental material for In Vitro Model Exploring the Mechanisms of Dextrose Prolotherapy: Fibroblasts Exposed to Clinical Concentrations of Dextrose Exhibit Significant Rebound Effects 48 Hours After Exposure
Supplemental material, sj-docx-1-car-10.1177_19476035251408601 for In Vitro Model Exploring the Mechanisms of Dextrose Prolotherapy: Fibroblasts Exposed to Clinical Concentrations of Dextrose Exhibit Significant Rebound Effects 48 Hours After Exposure by Bethany R. Harting, Sarah K. Fox, Adam D. Foster, Thomas M. Motyka and Amy N. Hinkelman in CARTILAGE
Supplemental Material
sj-xlsx-2-car-10.1177_19476035251408601 – Supplemental material for In Vitro Model Exploring the Mechanisms of Dextrose Prolotherapy: Fibroblasts Exposed to Clinical Concentrations of Dextrose Exhibit Significant Rebound Effects 48 Hours After Exposure
Supplemental material, sj-xlsx-2-car-10.1177_19476035251408601 for In Vitro Model Exploring the Mechanisms of Dextrose Prolotherapy: Fibroblasts Exposed to Clinical Concentrations of Dextrose Exhibit Significant Rebound Effects 48 Hours After Exposure by Bethany R. Harting, Sarah K. Fox, Adam D. Foster, Thomas M. Motyka and Amy N. Hinkelman in CARTILAGE
Footnotes
Acknowledgements
We thank Brooke Pritchett for her technical assistance and ongoing support in the research laboratory. We also acknowledge Cailee Dean for her tremendous effort in establishing the cellular protocols and experimental conditions, which were invaluable to this work.
Ethical Considerations
Not applicable.
Consent to Participate
Not applicable.
Consent for Publication
Not applicable.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This project was supported by intramural funding through the Campbell University Medical Research Fellowship established by a private philanthropic donation.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
All data for this study is available upon reasonable request.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.