
Abstract
Purpose
The mechanobiological response of cartilage redox balance might change in response to altered substrate availability. The purpose of this study is to investigate effects of key metabolic substrates (glucose, glutamine, and oxygen) on the mechanoresponsiveness of cartilage redox balance using a label-free imaging technique that measures autofluorescence from endogenous redox cofactors.
Findings
Compared with room oxygen tension, low oxygen tension had higher autofluorescence intensity in green (FAD) channel after a single sub-failure tensile load. Cartilage explants cultured in high glucose medium with glutamine supply had higher autofluorescence intensity in both channels (FAD and NADH/NADPH) compared with low glucose with glutamine group or high glucose without glutamine group, while no difference was observed between the latter 2 groups.
Conclusions
Low oxygen tension or high glucose culture medium with glutamine supply increases the mechanoresponsiveness of cartilage redox balance induced by sub-failure mechanical loading. Glutamine appears to partially serve as a glucose substitute in this process.
Introduction
Redox balance, the balance between oxidative and reactive reactions, is crucial for cartilage homeostasis. Disruption of redox balance is a hallmark of many diseases including osteoarthritis (OA),1 -3 a prevalent degenerative joint disease that features damage in articular cartilage. Cartilage redox balance is known to shift in response to mechanical stimuli. For example, excessive mechanical loading promoted mitochondrial superoxide generation and downregulated the production of superoxide dismutase 2, together causing oxidative stress and cartilage degeneration. 4 The production of reactive oxygen species (ROS), a group of highly reactive molecules that cause oxidative stress, can be correlated to loading parameters. 5
Cellular redox balance fluctuates in adaptation to changes in availability of metabolic substrates such as glucose, glutamine, and oxygen, as they play key roles in intracellular biochemical reactions. Glucose is a fundamental nutrient and prior studies demonstrate that glucose level alters cartilage redox balance. Exposure to high glucose caused increased ROS production in human OA chondrocytes. 6 Oxidative stress induced by high glucose was also observed in rat chondrocytes. 7 When glucose availability is limited, other substrates such as glutamine can still fuel cellular energy metabolism. 8 Deletion of glucose transporter increased glutamine oxidation in a mouse model. 9 Furthermore, glutamine metabolism is essential for chondrocyte function and regulates its response to inflammatory stimuli. Arra et al. 10 highlighted that chondrocytes used glutamine for energy production by showing that inhibition of glutaminase decreased ATP level. The same study also showed that glutamine deprivation or inhibition of glutamate dehydrogenase reduced expression of inflammation markers in murine chondrocytes treated with pro-inflammatory cytokine. Furthermore, glutamine also regulates cellular redox balance and can protect chondrocytes in ROS-induced apoptosis. 11 Another substrate that plays a key role in cellular redox balance is oxygen.12,13 In cartilage, specifically, the protective role of hypoxia (low oxygen) is well documented. Monolayer chondrocytes cultured in low oxygen showed less oxidative DNA damage compared with that in high oxygen. 14 Mechanical stress significantly increased ROS production in cartilage explants cultured in 20% oxygen compared with 1% or 5% oxygen, 15 highlighting the potential protective effects of low oxygen.
In vitro mechanical loading is widely used when studying cartilage mechanobiological response and demonstrates that cartilage is responsive to various loading modes including but not limited to compression,5,15,16 tension, 17 shear, 16 and hydrostatic pressure. 18 Although in vitro loading models are informative, the confounding effect of environmental conditions during culturing and/or loading is often overlooked. Pathological joint changes, however, could alter substrate availability and the corresponding mechanoresponsiveness of cartilage redox balance. For example, fewer hypoxic cells were observed in all layers of OA cartilage compared with control in mice model, as measured by a hypoxyprobe (pimonidazole hydrochloride). 19 High glucose level, a feature of diabetes, can increase inflammatory response and damage cartilage matrix.20,21
This study aims to identify the effects of metabolic substrates (glucose, glutamine, and oxygen) on the mechanoresponsiveness of cartilage redox balance using optical redox imaging (ORI). ORI is a label-free imaging technique that measures the autofluorescence from flavin adenine dinucleotide (FAD) and nicotinamide adenine dinucleotide and its phosphate (NAD(P)H), vital cofactors in cellular redox reactions. 22 The findings shall deepen current understanding of cartilage mechanoresponsiveness and highlight the potential of ORI as a tool of evaluating cartilage redox balance.
Methods
Sample Collection
Female F2 line pigs from Swine Research and Teaching Center (Age 4–6 months) were used in this study. Right patellae were collected within 1 hour after sacrifice and stored in phosphate-buffered saline (PBS) with 1% penicillin-streptomycin (Quality Biological) during transportation. PBS was filtered and stored in autoclaved containers to minimize contamination. Full-thickness patellar cartilage was manually cut into approximately 0.3 mm x 12 mm strips. In total, 120 strips collected from 7 pigs were used. There were 6 groups that were subject to distinct nutrient and oxygen conditions with 20 strips in each group.
Tissue Culture
Cartilage strips were randomly assigned to 1 of 3 culture conditions: high glucose with glutamine (H+), high glucose without glutamine (H–), and low glucose with glutamine (L+). The components of the culture medium were the same for all 3 conditions except for the basal DMEM, 50 μg/mL
All samples were cultured at 37 °C in 5% O2 and 5% CO2 for at least 14 hours (overnight) to equilibrate before mechanical testing. All tests were done within 72 hours of culturing. Culture medium was changed at noon of day 2 and no sign of contamination was observed during tissue culturing. Samples stayed in the incubator until right before the test started and were left in PBS at room temperature throughout the test.
Mechanical Stimuli
A single tensile load was applied to 2 strips simultaneously using a custom-built microscope top loading device
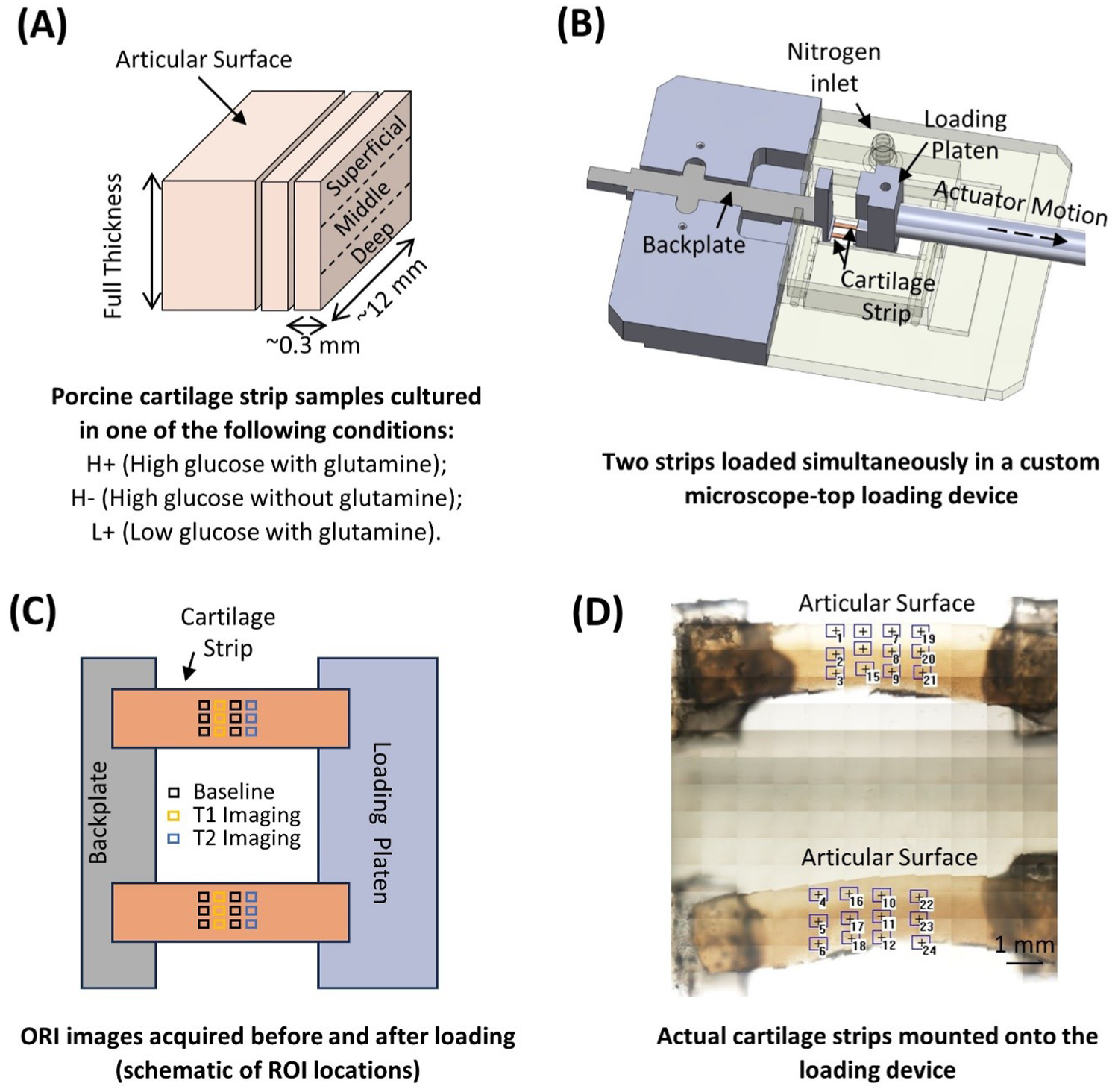
Overview of methods.
Optical Redox Imaging
ORI was done before, shortly after (within approximately 5 minutes, T1), and 30 minutes (T2) after the mechanical stimuli was applied. Briefly, 12 regions of interest (ROIs) were selected near the center of the sample
After all testing was complete, the mean intensity of the whole view in each ROI was measured using Matlab (R2023b). For each sample, the intensity of the 2 pre-loading ROIs in each zone was averaged to generate a baseline value, and the intensity of each post-loading ROI was normalized to the baseline value in the corresponding zone.
Oxygen Control
During imaging and mechanical loading, samples were randomly assigned to either room oxygen (RO2) or low oxygen (LO2). Low oxygen was achieved by a turning on a nitrogen tank connected to the chamber. A parafilm covered the chamber to reduce air flow. The dissolved oxygen in PBS was measured using an oxygen sensor (MW600 dissolved oxygen meter, Milwaukee Instruments). The oxygen concentration was 5.16 ± 0.37 mg/L and 2.03 ± 0.47 mg/L for HO2 and LO2 group, respectively. The equivalent oxygen tension in percentage of atmosphere was converted based on Henry’s Law:
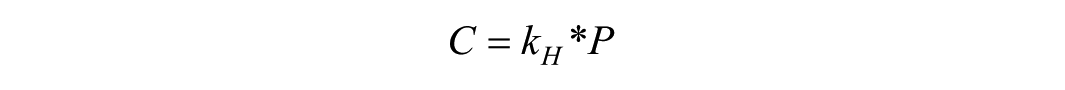
where C is the concentration of oxygen, kH is Henry’s constant, and P is the partial pressure of oxygen. Approximately, the oxygen tension for HO2 and LO2 group was 12.4%±0.9%, and 4.9%±1.1%, respectively.
Statistics
Statistical analysis was done in RStudio. For each time slot, Aligned Rank Transformed ANOVA was performed to analyze the effect of Nutrients (H+, H–, L+), Oxygen (NO2, LO2), Loading (0.1 s–1, 1.0 s–1), and Zone (SZ, MZ, DZ) and their interactions to the normalized autofluorescence intensity ratio in Green and DAPI channel, separately
Results
Effect of Substrate Availability
Nutrient was a significant main effect for green channel in both T1 and T2, and for the DAPI channel in T1
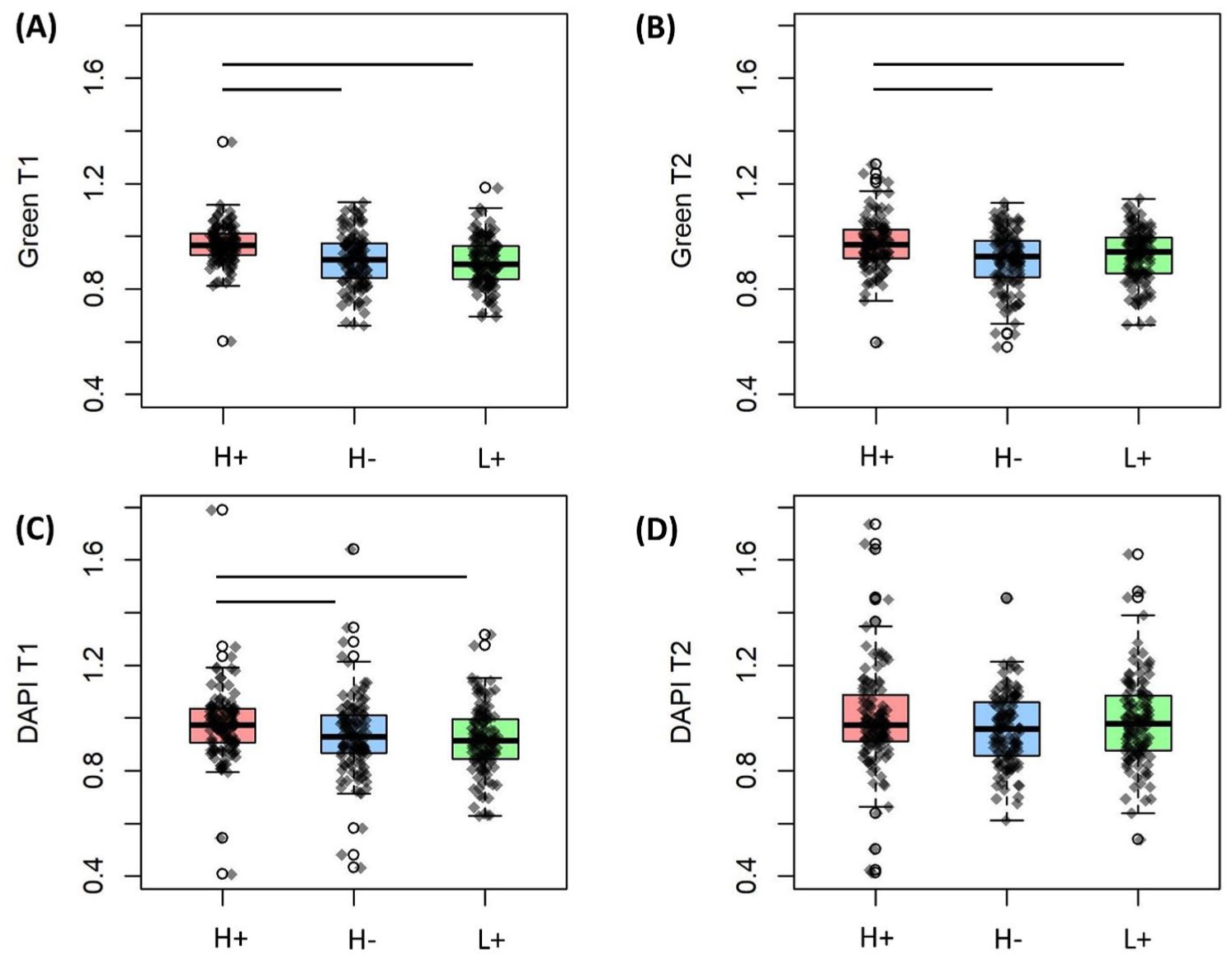
Effect of glucose (H or L) and glutamine (+ = with, – = without) on green channel intensity at
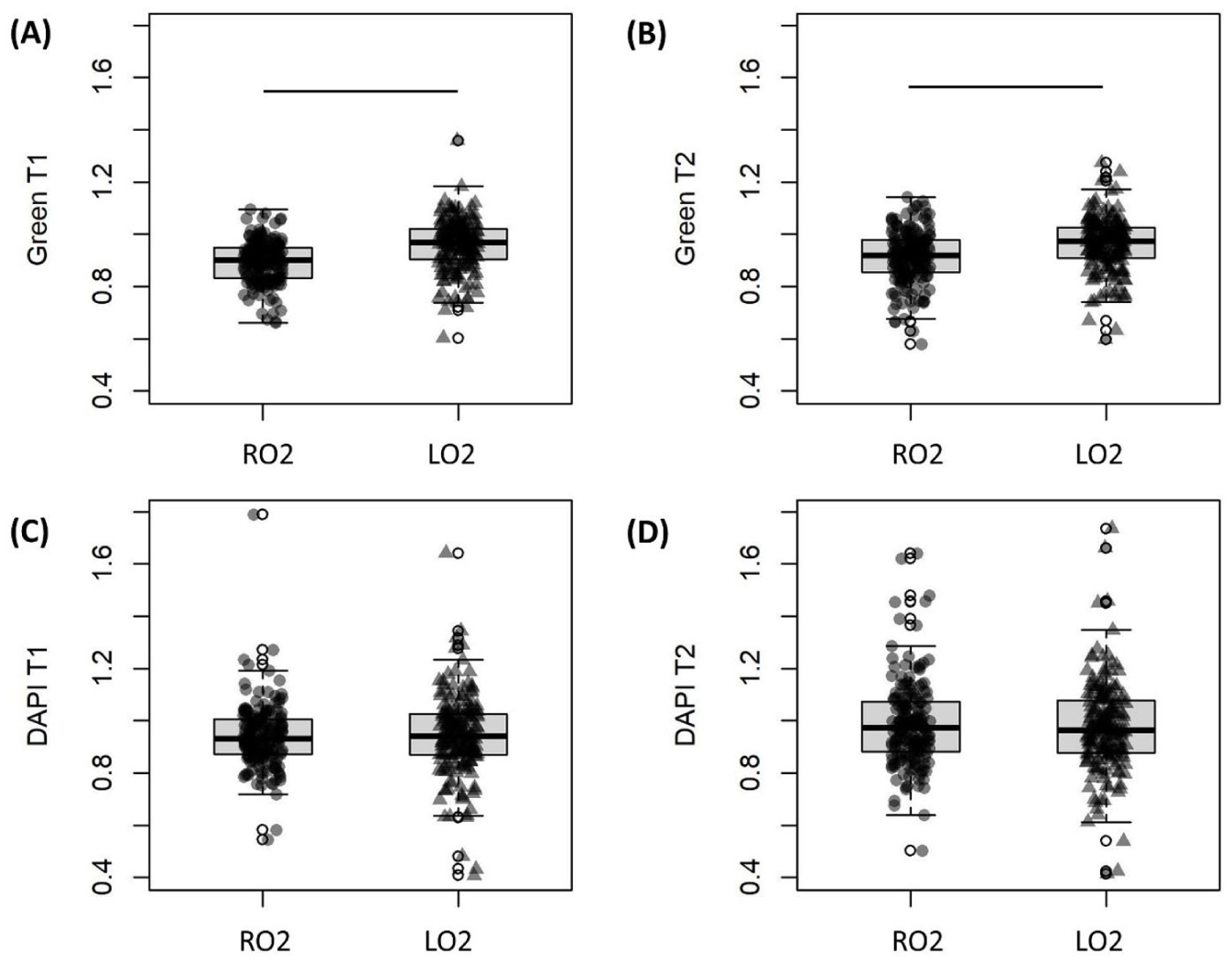
Effect of oxygen tension (RO2 = room oxygen tension, LO2 = low oxygen tension) on green channel intensity at
Interactive Effect of Nutrients Availability and Oxygen Level
Two-way interaction between nutrient and oxygen was significant for both channels in T2 but not T1
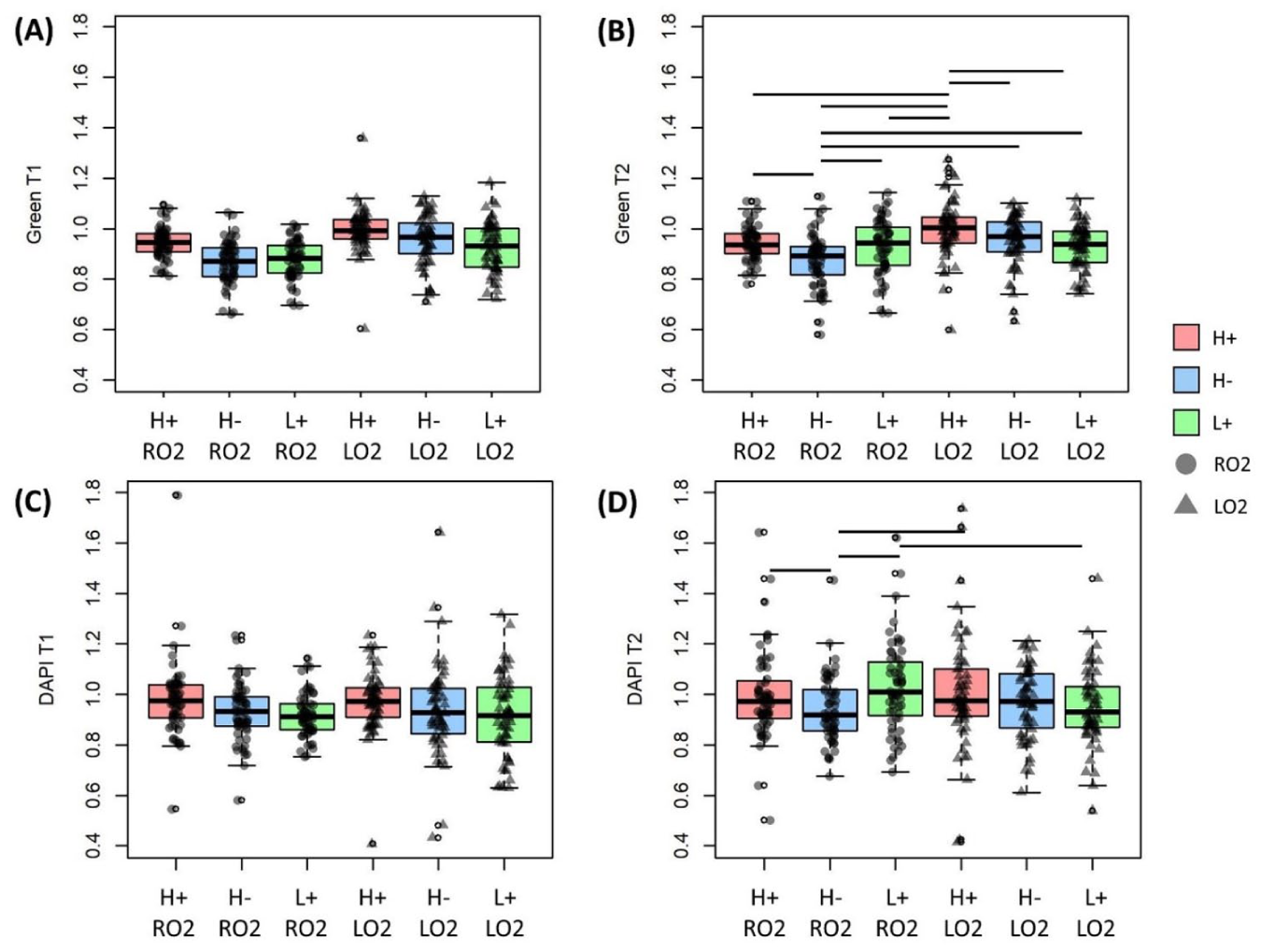
Interactive effect of nutrients (glucose, H or L, and glutamine, + or -) and oxygen tension (RO2 = room oxygen, LO2 = low oxygen) on green channel intensity at
Individual and Interactive Effects of Other Factors
Besides nutrients and oxygen, several other main effects and interactions were also significant. Zone was a significant main effect for the green channel in both time slots. In T1, normalized intensity in the superficial zone was higher than that in the middle zone. In T2, the superficial zone has a higher normalized intensity than both middle and deep zones
Discussion
This study applied sub-failure tensile loading to induce changes in autofluorescence intensity of redox cofactors in cartilage. Results demonstrate that availability of metabolic substrates altered the loading-induced change in cartilage redox balance. Specifically, culturing in high glucose (25 mM) with glutamine increased the autofluorescence intensity ratio of both FAD and NAD(P)H when compared with high glucose without glutamine or low glucose (5.5 mM) with glutamine. Compared with loading under high oxygen tension, loading in low oxygen tension increased FAD intensity but did not affect NAD(P)H intensity. The interaction effect between nutrients (glucose and glutamine) and oxygen was not observed immediately after loading, but was significant at ~30 minutes post-loading.
With glutamine, the higher FAD and NAD(P)H intensity of the high glucose group might be attributed to enhanced oxidative phosphorylation and glycolysis, respectively, as glucose is the fundamental substrate for both processes.25,26 Furthermore, environmental glucose can alter the mechanobiological response of cells. In mice podocytes, culturing in high glucose enhanced loading-induced increases in glucose uptake. 27 In this study, cartilage explants were cultured in high glucose medium for at least 14 hours before testing, which could elevate glucose uptake in response to mechanical stimuli and further augment the difference between low and high glucose conditions. There was no significant difference between H– and L+ groups, implying glutamine may partially substitute the role of glucose. In fact, this function of glutamine has been documented. Conversion of pyruvate from glucose is an important step for the tricarboxylic acid (TCA) cycle, a fundamental function of cellular energy metabolism. 13 C labeling showed that glutaminolysis pathway was significantly increased and contributed to TCA cycle in response to glucose deficiency. 28 Inhibiting transportation of pyruvate would also activate glutamine metabolism to generate oxaloacetate and acetyLCoA, maintaining the TCA cycle and improving cell survival. 29
Because oxygen directly participates in oxidative phosphorylation in which FAD is generated, 22 we expected to see higher FAD intensity in high oxygen group. The result, however, was contradictory to this hypothesis. This could be due to the fact that low oxygen tension better represents in vivo conditions for cartilage and is therefore favorable to cartilage mechanoresponsiveness. Supporting this concept is prior research, where mechanical cues increased COL2A gene expression in mice cartilage when loading was applied in low oxygen tension, but not in high oxygen. 30 In human inner meniscus tissues, which are similarly low in oxygen in vivo, it was also found that the effect of dynamic compression to up/downregulate certain genes was more prominent in low oxygen condition. 31
The interactive effect between oxygen tension and glucose availability was expected, as glucose transporters are responsive to oxygen sensing molecules (hypoxia-inducible factor alpha, HIFα) in cartilage. 32 For example, equine chondrocytes cultured in 1% oxygen had significantly higher glucose uptake and glucose transporter activity compared with that in 20% oxygen. 33 This established information was consistent with our finding that H+LO2 group had significantly higher green channel intensity compared with L+LO2, but there was no difference between H+RO2 and L+R02, as increased glucose intake allows for fuel for oxidative phosphorylation. Surprisingly, a similar effect was not observed for NAD(P)H. Because NAD(P)H participates in multiple metabolic process,34 -36 it is hard to interpret changes in NAD(P)H intensity or attribute it to a certain pathway.
This study came with several limitations. The samples were manually cut, which could introduce sample-to-sample variations in thickness and geometry. As autofluorescence from cells that were out of focus might still contribute to the measurement, this variation would elevate the noise within the data. Besides, consistent cell density was assumed based on tissue source and health, but the number of cells in each ROI was not quantified and this could introduce variation in measured intensity. Some samples were tilted before loading applied and/or detached during loading, making the loading profile not the exact same for all samples. This discrepancy could make interpretation of the effect of loading rate challenging and contribute to the noise as well. Apoptosis as a result of experimental conditions may also influence results. Cutting causes mechanical injury that is known to induce chondrocyte apoptosis.37,38 However, this effect would be consistent across groups because all samples were cut to the same shape using the same methods. Based on a prior study showing that high glucose (30 mM or 40 mM) upregulated apoptosis in human chondrocytes, 39 there may have been increased apoptosis in H+ and H– groups. This discrepancy between high and low glucose groups might confound the observed interaction effect between nutrient and mechanical stimuli to ORI metrics to some extent, and a control group without loading would help to clarify the effect of glucose level on baseline ORI. However, this confounding effect from glucose was unlikely dominating the results, because the documented effect of high glucose was observed after 72 hours of treatment, which is the upper limit of tissue culturing in this study. Autofluorescence intensity measured in this study was the average intensity of the whole field. We assumed consistent cell density across samples given the tissue source, but the variation in total cell numbers in each field could still contribute to the measurements. Both FAD and NAD(P)H participate in multiple metabolic pathways, and not only their intensity, but also their lifetime are indicators of cellular redox balance.22,40,41 However, we were not able to distinguish autofluorescence signals from cofactors in different pathways or measure fluorescence lifetime in the current setup, which limits the richness of the data. In addition to FAD and NAD(P)H intensity, optical redox ratio, FAD / (FAD + NAD(P)H), could also be used to provide a global view of redox balance. However, in this study we analyzed FAD and NAD(P)H intensity separately to investigate the potential difference in the effect of loading to cartilage autofluorescence.
Despite the limitations, this study provides new information on an aspect of cartilage mechanobiology that is often overlooked, and showed that availability of metabolic substrates affects cartilage redox balance in response to mechanical stimuli. Specifically, adding glucose or glutamine alone increased post-loading autofluorescence intensity of both FAD and NAD(P)H, and the effect was more notable for FAD and immediately after loading. Post-loading FAD intensity was higher in the low oxygen group compared with high oxygen. Furthermore, there were temporal changes in this effect, as the interaction between nutrients and oxygen was only significant 30 minutes after loading. Given the difference between in vivo conditions and commonly used in vitro culture conditions, including but not limited to the level of glucose, glutamine, and oxygen, this study implies that caution should be taken when interpreting results of cartilage loading models, and attention should be paid to culture conditions.
Supplemental Material
sj-docx-1-car-10.1177_19476035251377532 – Supplemental material for Cartilage Redox Balance Is Influenced by Substrate Availability
Supplemental material, sj-docx-1-car-10.1177_19476035251377532 for Cartilage Redox Balance Is Influenced by Substrate Availability by Jingyi Wang and Corinne R. Henak in CARTILAGE
Footnotes
Ethical Considerations
This study was exempt from ethical approval by the Institutional Animal Care and Use Committee (IACUC) because of the purchase of cadaveric tissues.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Funding from NSF CMMI BMMB 2237707 is gratefully acknowledged.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.