
Abstract
Objective
To investigate the novel role of miR-708-5p in osteoarthritis (OA) and its potential as a therapeutic target through regulation of NOX4/NF-κB signaling.
Methods
Expression levels of miR-708-5p were analyzed in OA cartilage using GEO datasets and validated in interleukin (IL)-1β-treated primary human chondrocytes. Gain- and loss-of-function experiments were performed using miR-708-5p mimics and inhibitors to evaluate its effects on inflammation, extracellular matrix metabolism, apoptosis, and oxidative stress. Direct targeting of NOX4 by miR-708-5p was confirmed through bioinformatic prediction, luciferase reporter assays, and rescue experiments.
Results
miR-708-5p was significantly downregulated in OA cartilage and IL-1β-treated chondrocytes. Overexpression of miR-708-5p attenuated IL-1β-induced inflammatory responses by suppressing pro-inflammatory cytokines (IL-1β, IL-6, tumor necrosis factor [TNF]-α), inhibiting matrix-degrading enzymes (MMP3, ADAMTS-4), and enhancing anabolic factors (COL2A1, SOX9). miR-708-5p protected against chondrocyte apoptosis by regulating Bcl2/BAX and caspase-3 expression. It also increased chondrocyte proliferation in EdU assays and reduced reactive oxygen species (ROS) production. Mechanistically, miR-708-5p directly inhibited NOX4, reducing ROS generation and nuclear factor kappa B (NF-κB) activation. NOX4 overexpression reversed the protective effects of miR-708-5p, confirming the functional significance of this regulatory axis.
Conclusion
miR-708-5p is downregulated in OA and exerts chondroprotective effects. These findings suggest that restoring miR-708-5p expression may effectively suppress the NOX4/NF-κB axis and modulate chondrocyte inflammation, oxidative stress, apoptosis, and matrix degradation.
Introduction
Osteoarthritis (OA) is a prevalent chronic degenerative joint disorder characterized by progressive cartilage loss, synovial inflammation, abnormal subchondral bone remodeling, and osteophyte formation.1-3 Current therapeutic approaches remain primarily palliative, ranging from early-stage anti-inflammatory medications to advanced disease surgical interventions.4,5 The molecular mechanisms underlying OA pathogenesis remain incompletely understood, and effective disease-modifying treatments are lacking, highlighting the urgent need for novel therapeutic strategies.
MicroRNAs (miRNAs) are small, single-stranded, non-coding RNAs, typically 20 to 25 nucleotides long, which serve as crucial post-transcriptional regulators of gene expression. These molecules bind to complementary sequences in the 3'-untranslated regions (3'-UTRs) of target messenger RNAs (mRNAs), leading to either mRNA degradation or translational suppression. 6 Despite the current lack of U.S. Food and Drug Administration (FDA) approval, miRNAs represent promising therapeutic candidates for OA treatment. 7 Their ability to simultaneously target multiple genes makes them uniquely suited to address OA’s complex pathology, while their endogenous nature and flexible targeting suggest a favorable safety profile. 8 These characteristics and growing evidence of their regulatory roles in OA strongly support their continued therapeutic development.9-11
miR-708-5p has emerged as a potential regulatory factor in joint diseases, with conflicting roles reported across different models. In rheumatoid arthritis (RA), miR-708-5p was downregulated in synovial tissues, where its supplementation induced synoviocyte apoptosis through Wnt3a/β-catenin pathway inhibition in rats. 12 Conversely, elevated miR-708-5p was observed in rabbit knee OA, and its repression promoted cartilage repair via SATB2 targeting. 13 Recent temporomandibular joint (TMJ) studies found that miR-708-5p deficiency contributed to OA through TLR4/NF-κB pathway activation. 14 These divergent findings across different joint conditions highlight the context-dependent nature of miR-708-5p regulation.
Inflammation, central to OA, is primarily mediated through the cytokine interleukin-1β (IL-1β).15,16 IL-1β is one of the major catabolic factors involved in OA, and many of its biological effects, such as suppression of extracellular matrix (ECM) synthesis and cell apoptosis, are mediated through reactive oxygen species (ROS) generation. NOX4, a major producer of ROS in chondrocytes, has emerged as a crucial mediator of oxidative stress and inflammation in OA pathogenesis. 17 IL-1β stimulation significantly upregulates NOX4 activity, leading to increased ROS production, which subsequently activates redox-dependent transcription factors such as NF-κB and AP-1. 18 This activation triggers matrix catabolism and chondrocyte death through the increased expression of matrix metalloproteinases (MMPs), ADAMTS proteins, and caspase-3.19,20 Importantly, NOX4 activation promotes increased production of inflammatory cytokines and leads to the persistence of inflammation, hence sustaining pro-catabolic pathways causing cartilage degradation. Recent studies have demonstrated that NOX4 deficiency protects against cartilage degradation and osteophyte formation in experimental OA models, highlighting its potential as a therapeutic target.17,21
In this study, we conducted analyses of GEO datasets, revealing significant downregulation of miR-708-5p expression in OA articular tissues compared with normal controls. Subsequently, we investigated the functional role of mir-708-5p in an IL-1β-induced human chondrocyte model of OA. We identified NOX4 as a novel direct target of miR-708-5p through computational prediction and experimental validation. We used multiple molecular approaches to establish that miR-708-5p suppresses NOX4 expression and attenuates chondrocyte inflammatory responses. These findings reveal a novel miR-708-5p/NOX4 regulatory axis in OA pathogenesis and suggest miR-708-5p as a potential therapeutic target.
Materials and Methods
Materials
Recombinant human IL-1b protein was obtained from LEADGENE BIOMEDICAL, Inc. (Taipei, Taiwan). Antibodies against ADAMTS-4, SOX9, ACAN, MMP-3, MMP-13, iNOS, BAX, caspase-3, NOX4, and β-Actin were purchased from Proteintech Group (USA). Anti-COL2A1 was obtained from Arigo biolaboratories Corp (Taiwan). Antibodies against IL-1β, IL-6, and TNF-α were from Boster biological technology (USA). MMP-3 IL-6 and tumor necrosis factor-alpha (TNF-α) enzyme-linked immunosorbent assay (ELISA) kits were purchased from ABclonal Technology (Palo Alto, CA, USA). Polymerase chain reaction (PCR) Master Mix and Lipofectamine® 2000 were bought from Thermo Scientific. Total RNA was extracted using the RNAzol or TOOLSmart RNA Extractor reagent (BIOTOOLS) (Foster City, CA, USA). Protease and phosphatase inhibitors were purchased from BIOTOOLS (Taiwan).
Isolation and Culture of Primary Human Chondrocytes
Following Institutional review Board (IRB) approval (Blinded for peer review), human chondrocytes from individuals with OA (3 females, ages 80, 72, and 65 years) were extracted during total knee arthroplasty procedures. To isolate the chondrocytes, the articular cartilage slices were chopped finely with a scalpel blade. Cartilage tissue was digested overnight with 0.01% (w/v, 0.166 U/mL) collagenase P in a medium containing 10% fetal calf serum (FCS) at 37°C. Afterward, the freed cells were separated from tissue debris by filtration through a 70 µm nylon cell strainer and collected from the filtrate to remove the undigested cartilage. The chondrocytes were cultured with DMEM (high glucose) supplemented with 10% FCS,
Analysis of the GEO Database
Gene-level profiles in OA and normal articular cartilage and synovial tissue were analyzed using 2 GEO datasets: GSE105027 (comprising 12 OA and 12 normal samples) and GSE143514 (comprising 5 OA patients and 3 healthy controls).22,23 These datasets analyzed miR-708-5p expression levels between OA and normal tissues.
DNA Constructs
PCR amplified the full-length cDNA of the human NOX4 gene. The PCR fragments were then inserted into the KpnI and ApaI sites of the pcDNA3.1/myc-His A expression vector containing the cytomegalovirus (CMV) promoter. PCR amplified the full-length 3’UTR of the human NOX4 gene. The PCR fragments were cloned into the pmirGLO Dual-Luciferase miRNA Target Expression Vector (Promega, Madison, WI, USA). Mutant-type NOX4 3’-UTR (miR-708-5p binding site mutant) luciferase reporters were purchased from MDBio, Inc.
Luciferase Reporter Gene Assay
Chondrocytes were placed in 12-well plates and were transfected with the plasmid and miRNA or expression plasmids using Lipofectamine 2000. Forty-eight hours after transfection, the cells were collected and lysed. Then, the measurement of luciferase activity was described previously by Huang et al. 24 The luciferase activity was determined and normalized with the amount of total protein. Continuous data are presented as mean values and standard deviations (SDs) for 3 determinations.
Transfection
MiR-708-5p mimics and inhibitors are chemically synthesized in MDBio, Inc. (Taipei, Taiwan). The sequences of miR-708-5p mimics were 5’- AAGGAGCUUACAAUCUAGCUGGG-3’ for sense and 5’-CAGCUAGAUUGUAAGCUCCUUUU-3’ for antisense. The sequences of miR-708-5p mimics NC were 5’-UCAGAGAAAUCAUCAGGGCUUGG-3’ for sense and 5’-AAGCCCUGAUGAUUUCUCUGAUU-3’ for antisense. The sequence of miR-708-5p inhibitors was 5’-CCCAGCUAGAUUGUAAGCUCCUU-3’. The sequence for miR-708-5p inhibitors NC was 5’-AUAUGUGGACGUCCCUCCCUUAA-3’. Human chondrocytes in 6-well plates grown to about 80% confluence, 10 nM miR-708-5p mimics, 10 nM mimics negative control (mimics NC), 10 nM miR-708-5p inhibitor, and 10 nM inhibitor NC or 2 μg pcDNA-NOX4 or pcDNA vector were transfected into cells at 37 °C for 24 h, using Lipofectamine 2000 (Invitrogen). After transfection, the cells were stimulated with IL-1β for 24 hours and then utilized in subsequent experiments.
qRT-PCR
According to the manufacturer’s protocol, total RNA was extracted from chondrocytes with the RNAzol reagent (MOLECULAR RESEARCH CENTER, INC). Reverse transcription was synthesized using the Thermo Scientific Maxima First Strand cDNA Synthesis Kit (ThermoFisher, Waltham, MA, USA), respectively. The primers for qRT-PCR analyses were as follows: miR-27a forward: 5’-TTCACAGTGGCTAAG-3’; miR-27a reverse: 5’-GTGCAGGGTCCGAGGT-3’; U6 forward: 5’-TGCGGGTGCTCGCTTCGCAGC-3’; U6 reverse: 5’-CCAGTGCAGGGTCCGAGGT-3’; Odz4 forward: 5’-CACCCTTCG GATTCTGTACGAC-3,’ reverse: 5’-AGCTCCATGTCTTCCCATCAGC-3,’ COL2A1 forward: 5’-ACATGGAGACTGGCGAGACTTG-3,’ reverse: 5’- GGACAGCAGGCGTAGGAAGGTC-3’; SOX9 forward: 5’-CGGTCAGCCAGGTGCTCAAAGG-3,’ reverse: 5’-GAGCTTGCC CAGCGTCTTGCTG-3,’ MMP3 forward: 5’-GAACCTGTCCCTCCAGAACCTG- 3,’ reverse: 5’-TATGCGGCATCCACGCCTGAAG-3,’ ADAMTS4 forward: 5’-CAAGCATCCGCAATCCTGTCAG-3,’ reverse: 5’- ACACAGGTCCTGACGGGTAAAC-3,’ NOX4 forward:5’- TCTTGGCTTACCTCCGAGGATC-3,’ reverse: 5’-GTTGAGGGC ATTCACCAGATGG-3,’ IL-1β forward: 5’-TTCAGGCAGGCCGCGTCAGTTG-3,’ reverse: 5’-CGGAGCGTGCAGTTCAGTGATC-3,’ IL-6 forward: 5’-CCAGAGCTGTGCAGATGAGTAC-3,’ reverse: 5’- CAGGCTGGACTGCAGGAACTCC-3,’ BCL2 forward: 5’ -AGCTGCACCTGACGCCCTTCAC-3,’ reverse: 5’-GTGCCGGTT CAGGTACTCAGTC-3,’ caspase-3 forward: 5’- ATTGTGGAATTGATGCGTGA -3,’ reverse: 5’-CAACGATCCCCTCTGAAAAA- 3,’ GAPDH forward: 5’-GGCCAAGGTCATCCATGACAAC-3,’ GAPDH reverse: 5’-TCCCGTTCAGCTCAGGGATGAC-3.’ To measure miR-708-5p, reverse transcription was performed using the miRCURY LNA miRNA PCR Starter Kit (Qiagen, Hilden, Germany), and relative quantification was determined by normalization to U6 or GAPDH. The qRT-PCR assays were performed in triplicate, and the relative expression levels were calculated based on the 2−ΔΔCt method.
Western Blot
The cells were washed thrice with cold phosphate-buffered saline (PBS), and cell lysates were extracted with RIPA buffer (50 mM Tris–HC1, 150 mM NaCl, 1% IGEPAL CA-630, and 0.5% sodium deoxycholate) containing the protease and phosphatase inhibitor (BIOTOOLS, Taiwan). The total protein concentration of various lysates was determined according to the Bio-Rad Bradford Protein Assays kit using bovine serum albumin (BSA) as a standard. A semidry apparatus separated Equal amounts of proteins on 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to a polyvinylidene difluoride membrane. Primary antibodies against the following proteins were used: MMP-3, MMP-13, ADAMTS-4, SOX9, iNOS, BAX, IL-1β, IL-6, TNFα, COL2A1, ACAN, caspase 3, NOX4 and β-Actin were employed as primary antibodies. The immunoblot analysis was performed using a secondary mouse or rabbit immunoglobulin G (IgG) antibody coupled with horseradish peroxidase, which was detected by the Millipore ECL kit (Thermo Fisher Scientific, UK). The densities of the immunoblots were determined using an image analysis system installed with the BIO-ID software (VilberLourmat, France).
Edu Assay
We used the Click-iT™ EdU Cell Proliferation Kit (Invitrogen, ThermoFisher) to evaluate cell proliferation. Briefly, after transfection with miR-708-5p mimics, miR-708-5p inhibitor, or corresponding negative controls for 48 hours, chondrocytes were exposed to 20 μM EdU for 2 hours and fixed with 4% formaldehyde in PBS for 20 minutes. Thereafter, the cells were stained with Hoechst 33342, while proliferative cells were labeled with EdU staining in green. Afterward, they were photographed with fluorescence microscopy. EdU-positive (EdU+) cells were imaged using a fluorescence microscope and counted under a microscope.
ELISA Assay
Cells were transfected with various plasmids, pcNOX4 or miR-708-5p, after which they were exposed to IL-1β (10 ng/ml) for 24 or 72 hours. Then, the levels of IL-6 (cat no. KE00139, Proteintech, Rosemont, USA) and TNF-α (cat no.KE00068, Proteintech, Rosemont, USA) in the culture supernatants were detected using the commercial ELISA kits per the manufacturer’s standard protocols.
ROS Detection
Human chondrocytes were transiently transfected with the 10 nM miR-708-5p mimics, 10 nM mimics negative control (mimics NC), 10 nM miR-708-5p inhibitor, and 10 nM inhibitor NC for 24 hours, using Lipofectamine 2000 (Invitrogen). Then, the cells were stimulated with or without 10 ng/ml IL-1β for 24 hours. The cells were then incubated with 10 μM DCFH-DA in DMEM for 30 minutes at 37°C washed with ice-cold PBS. They detected the fluorescence via fluorescence microscopy at 488 nm excitation and 525 nm emission.
Statistical Analysis
All experiments were performed at least 3 times independently and data are presented as mean ± standard deviation (SD). Statistical analyses were conducted using GraphPad Prism software (version 10.0). For comparisons between 2 groups, Student’s t-test was applied. For multiple group comparisons, 1-way analysis of variance (ANOVA) was performed. For time-dependent experiments, 2-way ANOVA was used. A p value <0.05 was considered statistically significant.
Results
MiR-708-5p is Downregulated in OA Cartilage and IL-1β-Treated Human Articular Chondrocytes
Gene Expression Omnibus (GEO) database records (GSE105027, GSE143514) were first analyzed to evaluate the clinical relevance of miR-708-5p in OA pathogenesis. This analysis revealed significantly lower levels of miR-708-5p in cartilage and synovial tissues collected from OA patients compared with those from healthy controls
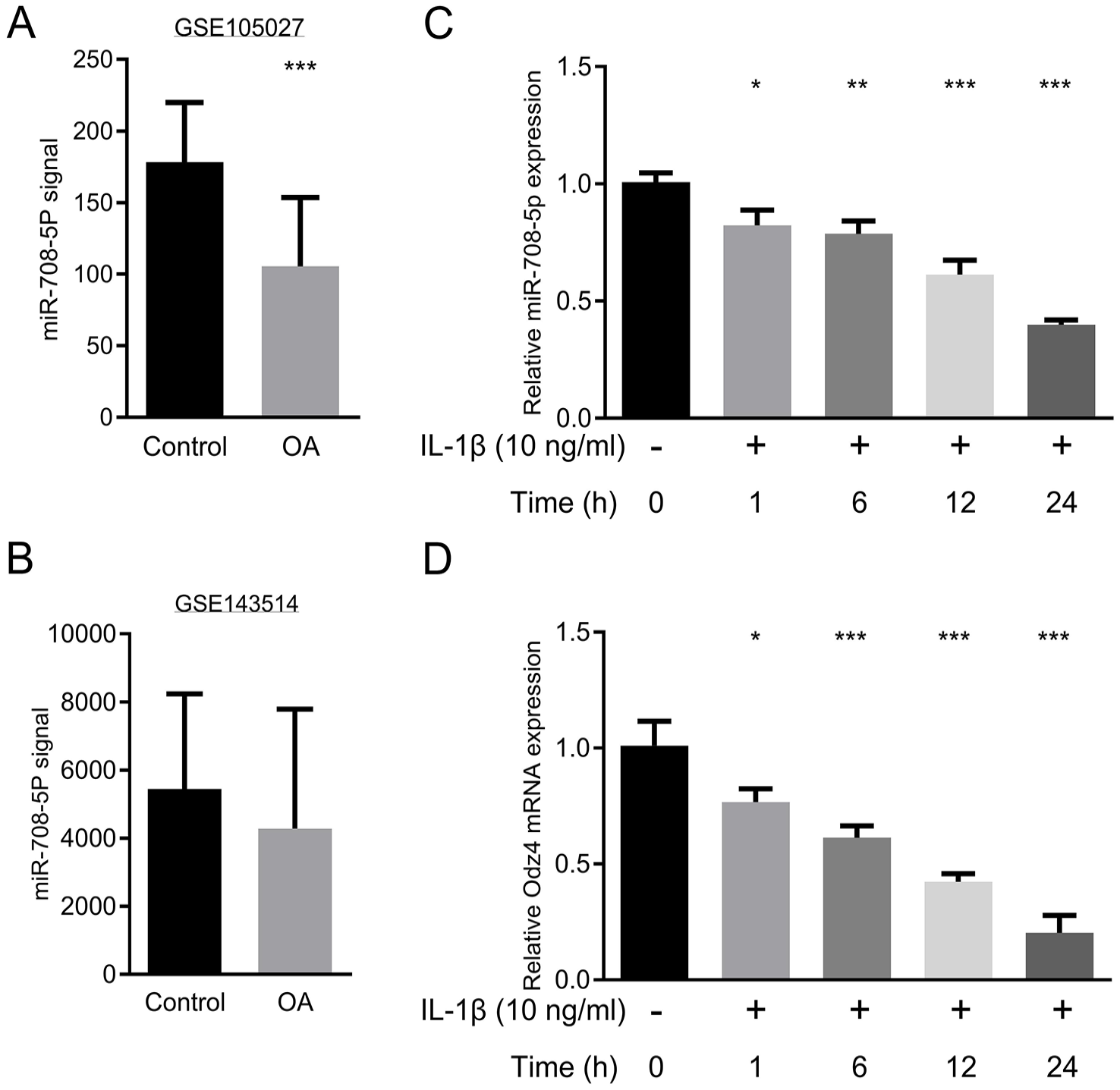
miR-708-5p expression is downregulated in osteoarthritic cartilage and IL-1β-stimulated chondrocytes.
To validate these clinical findings and investigate the regulatory mechanisms, we investigated miR-708-5p expression in an IL-1β-induced OA cell model. Primary human chondrocytes were treated with 10 ng/ml IL-1β at 0, 1, 6, 12, or 24 hours before evaluating miR-708-5p expression. IL-1β treatment significantly suppressed miR-708-5p expression in a time-dependent manner compared with the untreated controls
miR-708-5p Overexpression Attenuated IL-1β-Mediated Inflammation
Proinflammatory cytokines, including IL-1β, IL-6, and TNF-α, contribute to OA pathogenesis by enhancing proteolytic enzyme activity that degrades the ECM of cartilage.
26
To investigate the regulatory effects of miR-708-5p on IL-1β-induced inflammatory response in OA, human chondrocytes were transfected with miR-708-5p mimics or inhibitors to achieve overexpression or suppression, respectively. Transfection efficiency was verified by RT-qPCR
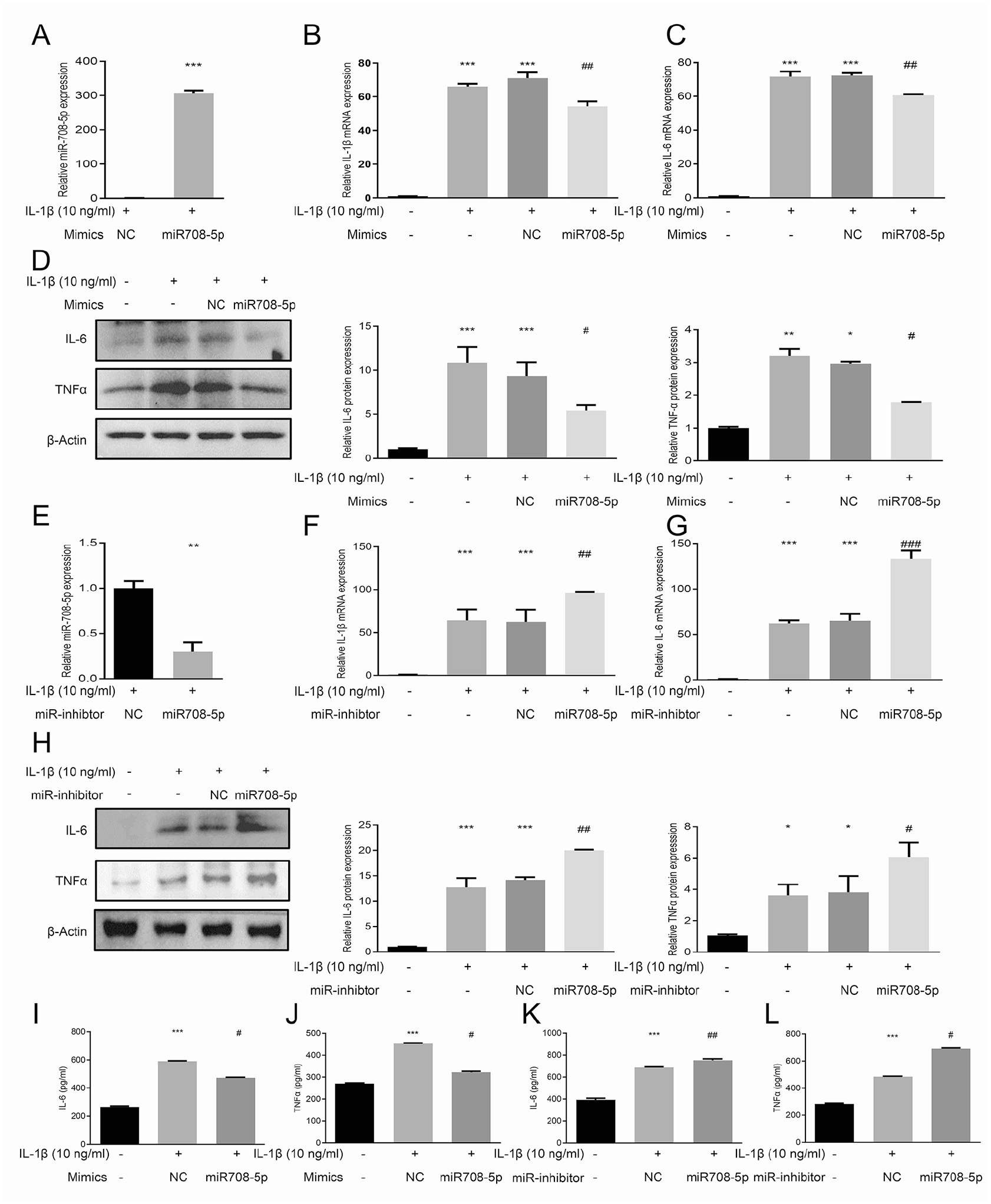
miR-708-5p inhibits IL-1β-induced inflammatory responses in human chondrocytes.
To further characterize the anti-inflammatory effects of miR-708-5p, we measured the secreted protein levels of IL-6 and TNF-α in cell culture supernatant through ELISAs. IL-1β treatment significantly increased IL-6 and TNF-α secretion compared to untreated controls
MiR-708-5p Modulates ECM Homeostasis by Regulating Anabolic and Catabolic Gene Expression in IL-1β-Induced OA
In OA, ECM degradation occurs through the breakdown of collagen and proteoglycan, primarily mediated by MMPs and ADAMTS proteases. We investigated the regulatory role of miR708-5p on cartilage ECM homeostasis by examining its effects on key anabolic factors (COL2A1 and SOX9) and catabolic enzymes (MMP3 and ADAMTS-4)
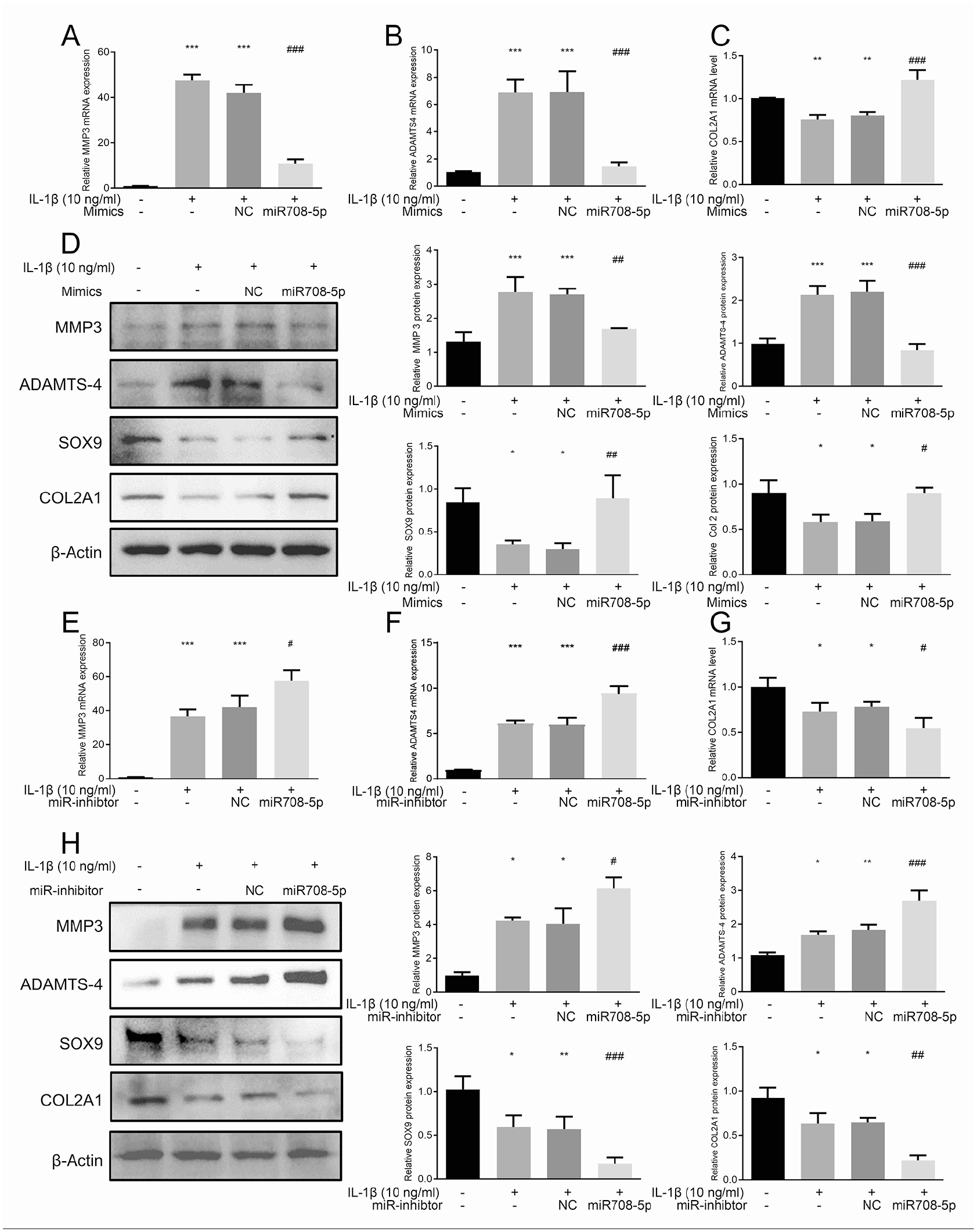
miR-708-5p regulates extracellular matrix homeostasis in IL-1β-stimulated chondrocytes.
IL-1β treatment for 24 hours significantly altered ECM component expression in human chondrocytes. At the mRNA level, IL-1β induced downregulation of anabolic factors COL2A1 while upregulating catabolic enzymes MMP3 and ADAMTS-4
Conversely, inhibition of miR-708-5p amplified the IL-1β-mediated effects, further elevating catabolic enzyme expression and enhancing suppression of anabolic markers
miR-708-5p Attenuates IL-1β-Induced Chondrocyte Apoptosis and Proliferation Impairment
We conducted gain- and loss-of-function analyses to examine miR-708-5p’s effects on chondrocyte survival. IL-1β treatment induced significant apoptosis, as demonstrated by the elevated expression of BAX and cleaved caspase-3, with concurrent bcl2 downregulation
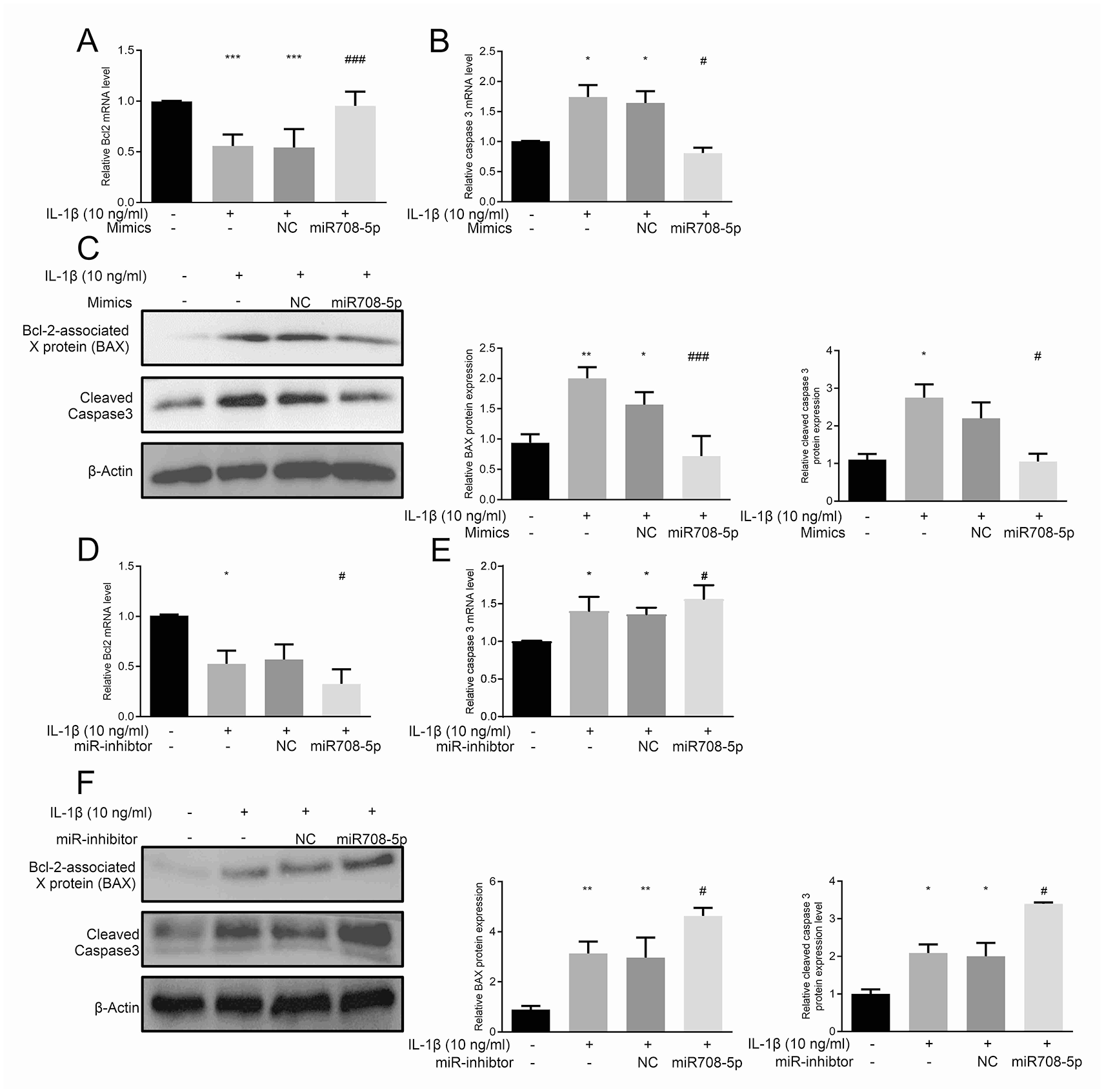
miR-708-5p modulates IL-1β-induced chondrocyte apoptosis.
Cell proliferation analyses using EdU incorporation assays demonstrated that IL-1β significantly suppressed chondrocyte proliferation, reducing EdU+ cell populations

miR-708-5p regulates chondrocyte proliferation under IL-1β stimulation.
Identification and Functional Validation of NOX4 as a Direct Target of miR-708-5p
To elucidate the molecular mechanism by which miR-708-5p attenuates IL-1β-induced cartilage degradation and inflammatory response in chondrocytes, we used computational algorithms (miRWalk) to predict the targets of miR-708-5p. Bioinformatic analyses revealed a specific binding site within the NOX4 3'-UTR that corresponds to the miR-708-5p sequence
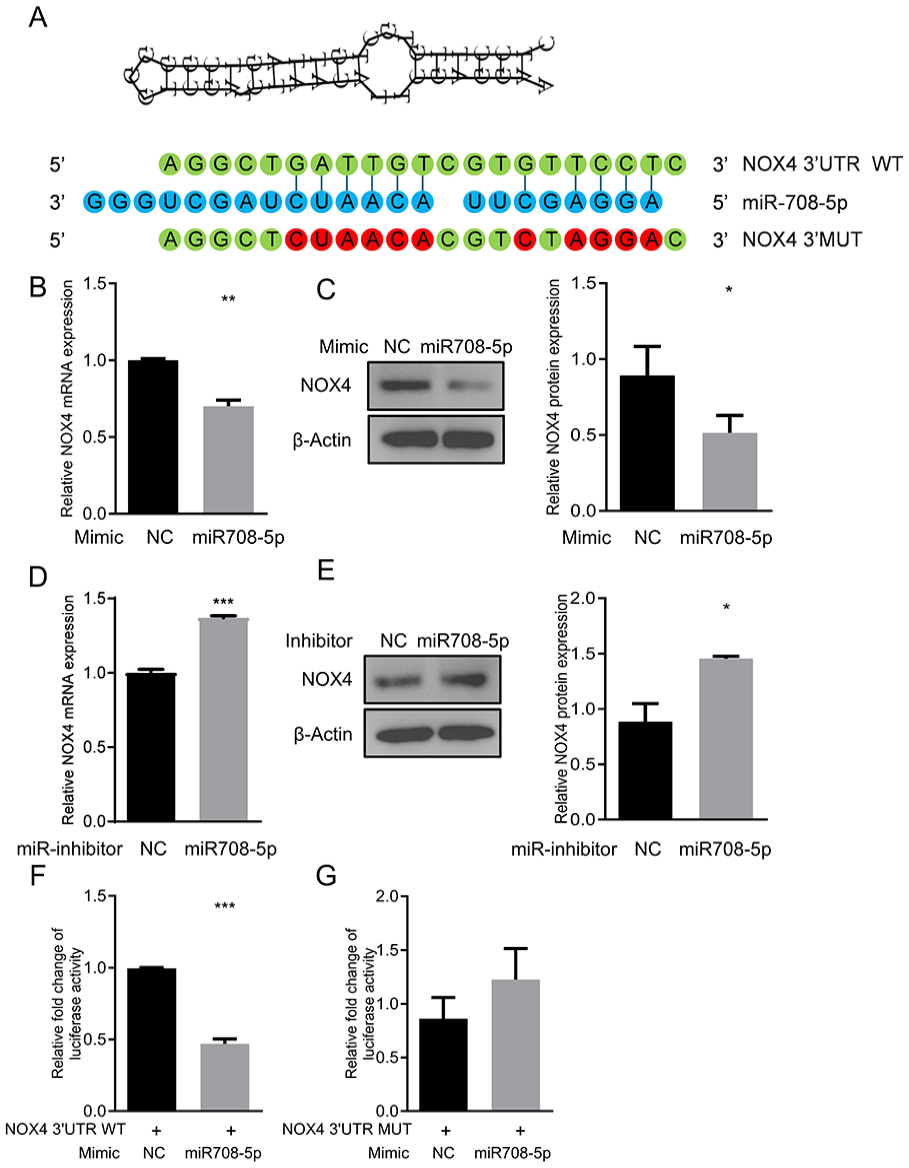
NOX4 is a direct target of miR-708-5p.
To validate NOX4 as a direct target of miR-708-5p, we performed a luciferase reporter plasmid carrying the wild-type (WT) or mutant (MUT) NOX4 3’-UTR, which were co-transfected with the miR-708-5p mimics or mimics-NC
miR-708-5p Upregulation Suppressed IL-1β-Induced NOX4 Expression, Oxidative Stress Response, and ROS Production in Chondrocytes
ROS and oxidative stress are crucial in cartilage homeostasis and OA pathogenesis.
27
Using the DCF-DA assay, we observed that IL-1β significantly elevated intracellular ROS levels, which was effectively suppressed by miR-708-5p overexpression
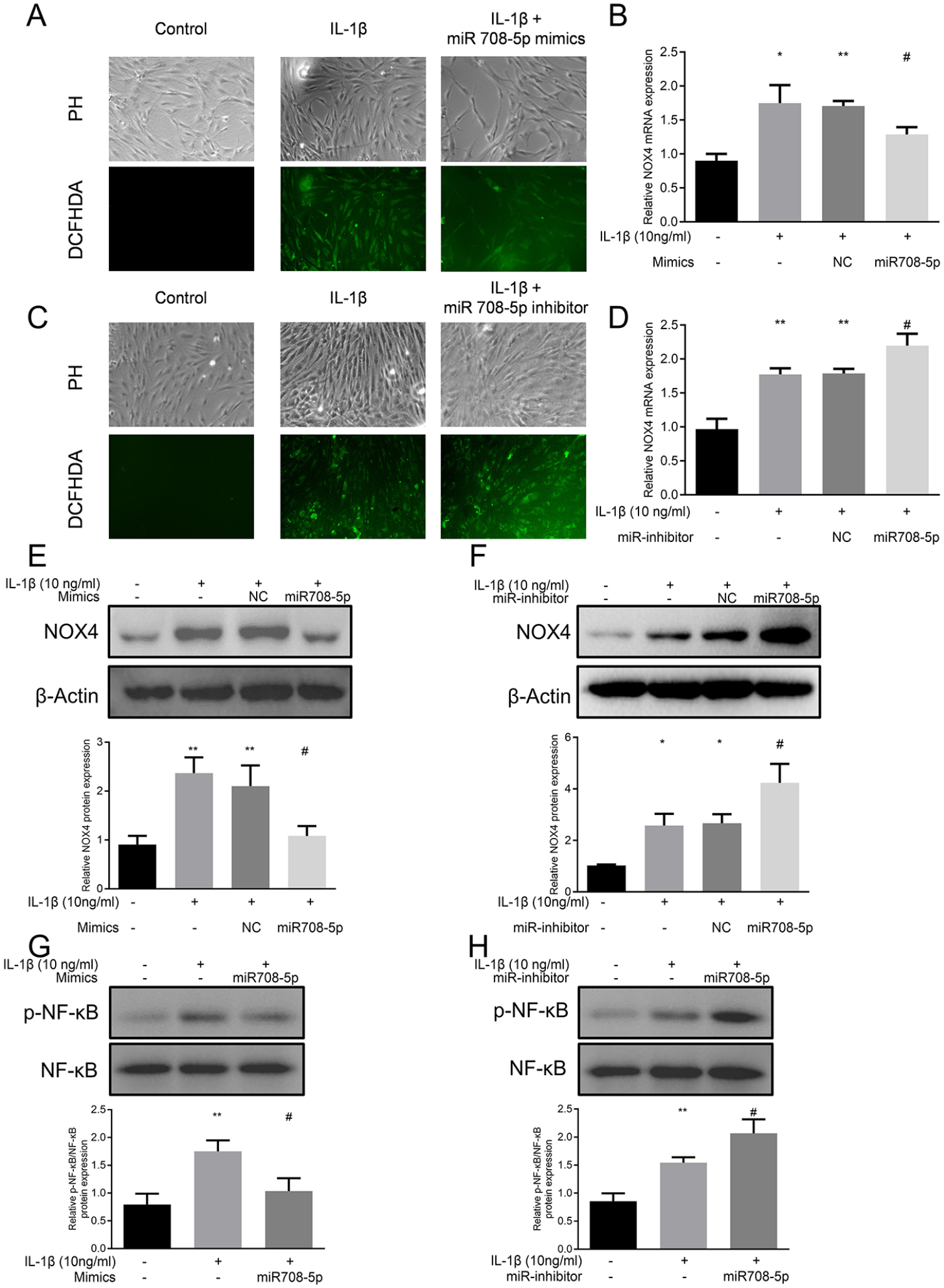
miR-708-5p modulates IL-1β-induced oxidative stress and NOX4/NF-κB signaling in chondrocytes.
At the molecular level, IL-1β treatment substantially upregulated NOX4 expression. qRT-PCR analysis revealed that this IL-1β-mediated induction of NOX4 mRNA was significantly suppressed by miR-708-5p overexpression
Given NOX4’s role in NF-κB signaling, we examined this downstream pathway activation. IL-1β stimulation significantly increased NF-κB phosphorylation, which was markedly diminished by miR-708-5p overexpression
The miR-708-5p/NOX4 Axis Modulated IL-1β-Induced Chondrocyte Damage
To confirm the functional significance of the miR-708-5p/NOX4 regulatory axis, we examined the effects of NOX4 overexpression in combination with miR-708-5p
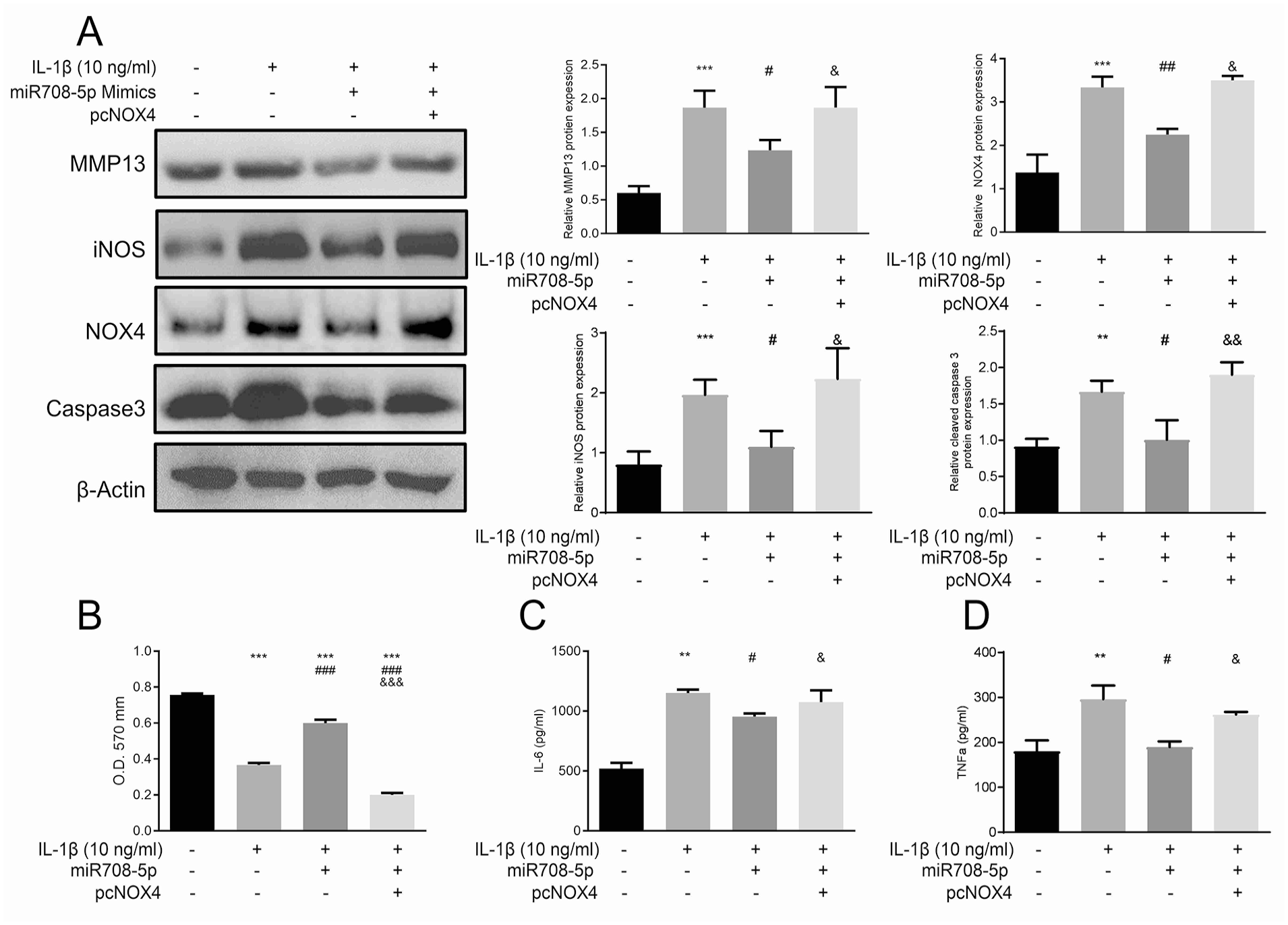
The miR-708-5p/NOX4/NF-κB axis modulates IL-1β-induced inflammatory and catabolic responses in chondrocytes.
ELISA analysis further confirmed the role of the miR-708-5p/NOX4 axis in regulating inflammatory cytokine production
Discussion
The present study identified miR-708-5p as a critical regulator of OA pathogenesis through the comprehensive analysis of GEO databases and in vitro models. Our investigations revealed significant decreases in miR-708-5p expression in OA patients’ articular cartilage tissues and IL-1β-treated chondrocytes. Using computational algorithms (miRWalk), we identified NOX4 as a direct target of miR-708-5p. Moreover, overexpression of miR-708-5p alleviated IL-1β-induced inflammatory response, apoptosis, cartilage degradation, and ROS production through the NOX4/NF-κB pathway in chondrocytes. Our data indicate miR-708-5p as a promising therapeutic modulator of the NOX4/NF-κB signaling axis, with a multifaceted role in maintaining cartilage homeostasis.
Aberrant expression of miRNAs is closely correlated with various diseases, including OA.28-30 miRNAs exert important functions in chondrocyte homeostasis by regulating the inflammatory responses, oxidative stress, ECM-related protein production, apoptosis, and senescence.31,32 Recent studies have revealed diverse roles of miR-708-5p in tissue homeostasis and stress responses, highlighting its potential significance in joint diseases.12,13,33 To validate the role of miR-708-5p in OA and address the knowledge gap in human studies, we investigated its function using primary human chondrocytes and clinical sample databases. Our study in human knee OA chondrocytes revealed decreased miR-708-5p expression in OA conditions, similar to Xu’s findings in rodent TMJ. 14
While Xu et al. 14 investigated miR-708-5p’s role in mechanically-induced TMJ OA through the TLR4/NF-κB pathway in rodent models, our study uniquely demonstrates that miR-708-5p regulates oxidative stress-mediated cartilage degradation through the NOX4/NF-κB axis in human chondrocytes. The consistency between our findings and Xu’s observations in TMJ regarding miR-708-5p’s protective role suggests a common regulatory mechanism in inflammatory conditions. However, Wang et al. 13 reported contrasting findings in their rabbit knee OA model, where miR-708-5p was upregulated in the chondrocytes and exhibited detrimental effects through targeting SATB2. This apparent dichotomy in miR-708-5p function likely reflects fundamental differences between the studies in experimental contexts, tissue characteristics, and disease progression stages.
Several key biological and methodological factors may explain these divergent findings across experimental models. First, the cellular response to OA varies significantly depending on the joint type, disease stage, and the degree of cartilage degeneration. Our samples were derived from end-stage OA patients undergoing total knee arthroplasty, where the cartilage may have undergone phenotypic transformation toward fibrocartilage—more like TMJ tissue than to the early-stage cartilage in Wang’s experimental model. Second, the disease models employed in each study triggered distinct pathogenic mechanisms. Wang et al. focused on post-surgical regeneration involving stem cell responses, while Xu et al. examined age-related mechanical stress, and our study investigated inflammatory/oxidative stress conditions. Third, these different disease triggers likely activate distinct molecular pathways. SATB2-mediated stem cell differentiation in the rabbit model versus stress response pathways leading to NF-κB activation in TMJ and human chondrocytes. The consistency between our findings in human chondrocytes and Xu’s observations in TMJ regarding miR-708-5p’s protective role through NF-κB regulation suggests this may be the predominant pathway in inflammatory and mechanical stress conditions. However, the contrasting results from Wang’s regeneration model highlight how miR-708-5p’s function can shift when the cellular context changes from stress response to repair. These context-dependent differences suggest therapeutic strategies targeting miR-708-5p must carefully consider the specific disease mechanism and joint type involved. These model-specific difference in miR-708-5p function underscored the need to identify its direct molecular targets in human OA, which would explain its protective effects in our experimental system.
Our identification of NOX4 as a direct target of miR-708-5p reveals a novel regulatory axis in OA that modulates oxidative stress and inflammatory responses. However, the relationship between miR-708-5p and ROS signaling is highly tissue-specific and disease-dependent. The relationship between miR-708-5p and ROS displays remarkable context specificity across different tissues and disease models.34-36 In osteoblasts, miR-708-5p protects against oxidative stress-induced apoptosis by targeting PTEN. 34 It also suppresses hypoxia/reoxygenation-induced inflammation and ROS generation in cardiomyocytes. 36 However, overexpression of miR-708-5p in cancer cells promoted ROS-induced apoptosis.37,38 Our study has established that in human chondrocytes, miR-708-5p specifically targets NOX4, a critical upstream regulator of ROS, thereby exerting protective effects against oxidative stress in OA pathogenesis.
After identifying the context-dependent nature of miR-708-5p, we sought to elucidate its molecular mechanism in human OA chondrocytes. IL-1β, a pivotal inflammatory mediator in OA pathogenesis, triggers a cascade of cellular responses in chondrocytes, including upregulation of NOX4 and p22phox proteins, leading to increased NOX4-derived ROS generation.39,40 This IL-1β-mediated NOX4 activation induces significant upregulation of matrix-degrading enzymes (MMPs and ADAMTs4) while simultaneously suppressing the synthesis of critical cartilage components like type II collagen and aggrecan, primarily through the activation of NF-κB, AP1, and HIF-1α signaling pathways.19,41-43 Using luciferase reporter assays, we confirmed miR-708-5p’s ability to suppress IL-1β-induced NOX4 expression, ROS production, and NF-κB pathway activation in chondrocytes. Furthermore, our rescue experiments revealed that NOX4 overexpression could reverse the protective effects of miR-708-5p on cell viability and inflammatory responses, confirming that the miR-708-5p/NOX4 axis plays a crucial role in regulating IL-1β-induced chondrocyte injury in OA.
While our findings establish NOX4 as the key target in chondrocytes, miR-708-5p exhibits diverse roles in inflammation across different tissues. It protects cardiomyocytes against hypoxia/reoxygenation-induced inflammation by targeting HMGB1 signaling and inhibits alcohol-induced liver inflammation through ZEB1/AKT/mTOR pathway regulation.36,44 Conversely, in dental pulp stem cells, miR-708-5p knockdown reduces LPS-induced inflammation and apoptosis. 45 Our study reveals that miR-708-5p specifically enhanced cartilage-specific matrix markers while reducing inflammatory mediators, suggesting its therapeutic potential in cartilage protection.
OA represents a highly intricate disorder affecting the entire joint, involving multiple tissues rather than cartilage. 46 The pathogenesis involves a complex interplay between cartilage, synovium, and subchondral bone that significantly affects disease progression and therapeutic outcome. While our study primarily demonstrates miR-708-5p’s protective effects in human chondrocytes through NOX4/NF-κB pathway regulation, emerging evidence indicates that miR-708-5p may exert pleiotropic effects across various tissues. The synovium contributes significantly to OA through its inflammatory response, producing pro-inflammatory mediators and proteolytic enzymes that drive cartilage degradation and joint tissue damage. 47 Notably, miR-708-5p has been found to play a protective role in the synovium by increasing cell apoptosis, inhibiting cell proliferation, and decreasing the migration capacity of synovial fibroblasts in rat RA models. 12 While these findings are promising, future studies are needed to investigate the therapeutic effect of miR-708-5p in the human OA models.
Moreover, subchondral bone remodeling is important in OA development and progression. 48 Research has revealed that miR-708-5p protects osteoblasts and may prevent osteoporosis. 34 However, in bone marrow stromal cells (BMSC), miR-708-5p negatively regulates osteoblast differentiation in BMSCs, and inhibition of miR-708-5p rescued the suppressive effects of glucocorticoids on BMSC function.49,50 It is important to note that these findings regarding miR-708-5p’s effects on bone cells and BMSCs were derived from non-OA models. Further investigations are warranted to elucidate the specific role of miR-708-5p in OA-related bone remodeling, as the pathological microenvironment in OA may significantly influence miR-708-5p’s functional outcomes.
Although miR-708-5p represents a promising therapeutic candidate for OA, its clinical translation remains challenging due to inherent delivery limitations. Unmodified miRNAs are unstable in physiological environments, prone to nuclease degradation, rapid clearance, and poor cellular uptake due to their size and negative charge. 51 These issues are further intensified by the dense, avascular structure of articular cartilage, which restricts molecular diffusion and limits retention. 52 Nevertheless, various emerging delivery strategies have shown promise to enhance stability, bioavailability, and targeted delivery of miRNAs. 53 These include viral vectors, lipid and polymer-based nanoparticles, inorganic carriers, extracellular vesicles, and peptide-based systems.53-55 These advances may offer a foundation for overcoming existing barriers and may facilitate the clinical translation of miR-708-5p–based OA therapy.
This study has several limitations. First, in vitro experiments were based on chondrocytes from only 3 elderly female OA patients, which limits generalizability to other age groups, sexes, or disease stages. Second, the GEO analysis used only 2 datasets with small sample sizes, reducing the strength of transcriptomic comparisons. Third, the IL-1β-induced cell model does not capture the full complexity of in vivo OA. Lastly, we did not assess in vivo efficacy or delivery strategies for miR-708-5p, which are critical for clinical translation. These limitations highlight the need for future studies utilizing in vivo models and clinical samples to validate our findings and establish the therapeutic potential of miR-708-5p in OA treatment.
Conclusion
Our study establishes miR-708-5p as a crucial regulator in OA pathogenesis through the NOX4/NF-κB signaling pathway. The consistent downregulation of miR-708-5p in both OA clinical samples and IL-1β-stimulated chondrocytes, coupled with its demonstrated ability to preserve cartilage integrity, suppress inflammatory responses, reduce oxidative stress, and maintain ECM homeostasis in IL-1β-stimulated chondrocytes, positions this microRNA as a promising therapeutic target. Future translational work should focus on targeted delivery of miR-708-5p to cartilage tissue and evaluate its therapeutic efficacy in OA animal models.
Footnotes
ORCID iDs
Ethical Considerations
This study was approved by the Institutional Review Board of Kaohsiung Medical University Hospital (KMUHIRB-F(I)-20220166).
Consent to Participate
Not applicable.
Consent for Publication
Not applicable.
Author Contributions
ZM Liu and SH Huang devised the project, the main conceptual ideas, and the proof outline. CC Wang and ZM Liu worked out almost all of the technical details and performed the numerical calculations for the suggested experiment. PC Shen and SJ Chen contributed to the final version of the manuscript. PY Tu and LM Chen contributed to the data analysis and figure visualization. CC Lu and YC Tien supervised the project.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by Kaohsiung Medical University Memorial Hospital (grant no: KMUH-113-3M35).
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
The corresponding author can provide the data supporting the present study upon reasonable request.