
Abstract
Objective. Caspases, cysteine proteases traditionally associated with apoptosis and inflammation, have recently been identified as important regulators of autophagy and reported within the growth plate, a cartilaginous part of the developing bone. The aim of this research was to identify novel autophagy-related molecules affected by inhibition of pro-apoptotic caspases in chondrocytes. Design. Chondrocyte micromasses derived from mouse limb buds were treated with pharmacological inhibitors of caspases. Autophagy-related gene expression was examined and possible novel molecules were confirmed by real-time polymerase chain reaction and immunocytofluorescence. Individual caspases inhibitors were used to identify the effect of specific caspases. Results. Chondrogenesis accompanied by caspase activation and autophagy progression was confirmed in micromass cultures. Expression of several autophagy-associated genes was significantly altered in the caspases inhibitors treated groups with the most prominent decrease for Pik3cg and increase of Tnfsf10. The results showed the specific pro-apoptotic caspases that play a role in these effects. Importantly, use of caspase inhibitors mimicked changes triggered by an autophagy stimulator, rapamycin, linking loss of caspase activity to an increase in autophagy. Conclusion. Caspase inhibition significantly affects regulation of autophagy-related genes in chondrocytes cultures. Detected markers are of importance in diagnostics and thus the data presented here open new perspectives in the field of cartilage development and degradation.
Introduction
Autophagy is involved in degradation of cytosol components and organelles 1 and thus represents an essential mechanism for either cell survival or death associated with tissue/organ development and maintenance.2,3 Autophagy is currently a hot topic in research promoted by the Nobel Prize award in physiology and medicine 2016, 4 which allows for a wide range of clinical applications in this field. In bone development (osteogenesis), autophagy is induced during differentiation of osteoblasts and participates in bone formation and homeostasis.5,6 In keeping with these functions, impaired regulation of autophagy is associated with several bone and cartilage disorders such as osteoporosis and osteoarthritis. 7 Recently, autophagic molecules are in focus as targets of osteoarthritic treatment. 8
In long bones, arising by endochondral ossification, a transient growth plate composed of chondroblasts/chondrocytes and their extracellular matrix allows for elongation of the bone. 9 Growth plate maturating chondrocytes display an autophagic phenotype. 10 Autophagy in these cells was recently shown to regulate secretion of collagen type II, 11 the major component of cartilage extracellular matrix. Since cartilage is mostly an avascular structure, cartilage cells grow in an under nourished, hypoxic environment. The nutrition of cells thus has to be ensured by alternative mechanism, such as autophagy, which has been proposed to allow hypertrophic cartilage cells to survive. 12 Autophagy also serves as a protective mechanism in adult cartilage 13 and is likely to precede the programmed cell death of chondrocytes.14,15 Beclin-1, one of the main autophagy markers, maintains chondrocyte survival, and its absence results in enhanced chondrocyte death. 16 Cleaved Beclin-1 loses its autophagic activity and can induce apoptosis, 17 and vice versa, overexpressed Beclin1 can inhibit apoptosis. 18
Programmed cell death (apoptosis) is mediated by caspases, a group of cysteine proteases. Two distinct pathways of caspase activation, intrinsic (mitochondrial) and extrinsic (receptor mediated), are evident. These pathways are initiated by apical caspases (-8, -9) and executed by a trio of death caspases (-3, -6, -7). 17 Notably, activation of pro-apoptotic caspases was recently demonstrated to occur also in non-apoptotic growth plate chondrocytes, 19 and in developing Meckel’s cartilage. 20 In keeping with this non-apoptotic role, caspase inhibition in vitro changed expression of osteogenic markers in developing chondrocytes.19,21 This is in agreement with caspases having novel non-apoptotic roles in addition to their role in cell death. 22 Along with engagement in apoptosis, cell cycle regulation, and cell differentiation, there is increasing evidence that pro-apoptotic caspases participate in autophagy and in autophagy-apoptosis cross-talk.18,23 Among pro-apoptotic caspases, the central caspase-3 was demonstrated as an inhibitor of autophagy in most studies. 24 The other 2 members of the “death trio,” caspase-6 and caspase-7, are also associated with a negative impact on autophagy.25,26
Application of autophagy on cartilage treatment is conditioned by understanding of autophagic processes. In the presented investigation, we aimed to define the effect of caspases inhibition to autophagy in chondrocytes. Therefore, chondrocytes were exposed to pharmacological caspase inhibitors, unique research tool, 27 to follow whether this affected the regulation of autophagy-related genes and to identify which pro-apoptotic caspase was engaged.
Material and Methods
Tissue Preparation and Immunohistochemistry
Paraffin-embedded specimens of mouse fore limbs (CD1 strain) were prepared 28 and processed to achieve 5-µm-thick histological sections. Primary antibodies (anti-Beclin, ab62557, Abcam, dilution 1:300, anti-LC3b cs3868, Cell Signaling, dilution 1:100) were applied overnight following antigen retrieval. The chromogenic reaction was mediated by a peroxidase-conjugated streptavidin biotin system (Vectastain) and substrate diaminobenzidine (Dako) to visualize positive cells as brown. Slides were counterstained by hematoxylin to distinguish the cell nuclei. In the case of immunofluorescence, Alexa Fluor 488 (Thermo Fisher Scientific) was applied as a secondary antibody. Nuclei were counterstained by ProLong Gold Antifade Mountant with DAPI (Thermo Fisher Scientific).
Micromass Cultures
Micromass cultures were utilized as a common in vitro method to investigate chondrogenesis. 29 Cells were obtained from mouse fore limbs at E12 by a standard protocol 30 and left until the following morning in order to attach to the surface. For caspase inhibition, pharmacological inhibitors: Pan Caspase Inhibitor Z-VAD-FMK, Caspase-2 Inhibitor Z-VDVAD-FMK, Caspase-3 Inhibitor Z-DEVD-FMK, Caspase-6 Inhibitor Z-VEID-FMK, Caspase-8 Inhibitor Z-IETD-FMK, Caspase-9 Inhibitor Z-LEHD-FMK, Pan Caspase OPH Inhibitor Q-VD (FMK001, FMK003, FMK004, FMK006, FMK007, FMK008, OPH001, R&D Systems) were applied to the micromass cultures at a concentration of 100 µM, according to the manufacturer’s recommendation and previous studies.21,31 In the controls, dimethyl sulfoxide (DMSO), the inhibitor vehicle, was added and the control micromass cultures were run in parallel. Autophagy was induced by addition of 500 nM rapamycin (R8781, Merck) to the micromass cultures. The medium, including DMSO, rapamycin, and caspase inhibitor, was changed every second day. 30 The experiments were performed in at least 3 replicates. The research was performed under the project GACR 19-12023S approved and supervised by the Czech Sciences Foundation including all ethics issues.
RNA Isolation, PCR Array, and Real-Time PCR
The cultured cells were harvested into 350 µL RLT lysis buffer (Qiagen, Valencia, CA) with β-ME (Sigma-Aldrich). RNeasy Mini Kit (Qiagen) was used for RNA isolation, Super Script VILO (Invitrogen) for cDNA transcription. PCR Arrays were applied to analyze 84 genes connected with autophagy and autophagy-apoptosis cross-talk (PAMM-084Z, Qiagen). Real-time polymerase chain reaction (PCR) was performed for expression analysis of Pik3cg (Mm00445038_m1, Thermo Fisher Scientific), Tnfsf10 (Mm01283606_m1, Thermo Fisher Scientific), and Ctss (Mm01255859_m1, Thermo Fisher Scientific). Reaction mixture contained the one-step master mix gb Ideal PCR Master Mix (Generi Biotech). Expression levels were calculated normalized against the level of actin (Mouse Actb, Mm02619580_g1, Thermo Fischer Scientific).
Immunocytofluorescence and TUNEL Assay
For immunocytofluorescence, micromass cultures were grown on cultivation glass and fixed by 4% PFA. Alcian blue staining was used for visualization of chondrogenic differentiation. Primary antibodies (Beclin, ab62557, Abcam; LC3b cs3868, Cell Signaling, Sox9, HPA001758, Sigma Aldrich), Pik3cg (MA5-26087 Thermo Fisher), Cleaved caspase-2 (PA5-39872, Thermo Fisher Scientific), Cleaved caspase-3, 9664, Cleaved caspase-6, 9761, Cleaved caspase-7, 9491, Cleaved caspase-8, 8592, and Cleaved caspase-9, 9509, all from Cell Signaling, were applied overnight/4°C. Alexa Fluor 488 or 568 (A11034, A10037, Thermo Fischer) was diluted 1:200 and then applied for 40 minutes at room temperature. Cytoskeleton was visualized by ActinGreen 488 ReadyProbes Reagent (Thermo Fischer); nuclei were detected by ProLong Gold Antifade reagent with DAPI (Thermo Fischer). For detection of apoptosis, ApopTag Fluorescein in Situ Apoptosis Detection Kit (S7110, Merck) was used. Externalization of phosphatidylserine was detected by Annexin V-FITC Apoptosis Staining/Detection Kit (ab14085, Abcam).
Statistical Analysis
PCR array data were statistically evaluated using the Qiagen Data Analysis Center (available online). The significance was determined as P < 0.05 and the threshold was established as −2/+2 fold-regulation. Three biological samples were used for all tested groups. Expression levels from real-time PCR were calculated by ΔΔCT method. Results were analyzed using 2-tailed t test or, in the case of specific caspase inhibitors, using ANOVA with Dunnett’s post hoc test.
Results
Beclin-1 and LC3b Are Expressed in the Prenatal Growth Plate
To evaluate autophagy during chondrogenesis in relation to limb development in vivo, immunohistochemistry was applied on histological sections of the mouse fore limb at different stages of development from embryonic (E) 12 (the stage corresponding to that used for micromass preparation) to E18 (when all zones of the growth plate are clearly detectable).
At E12 (
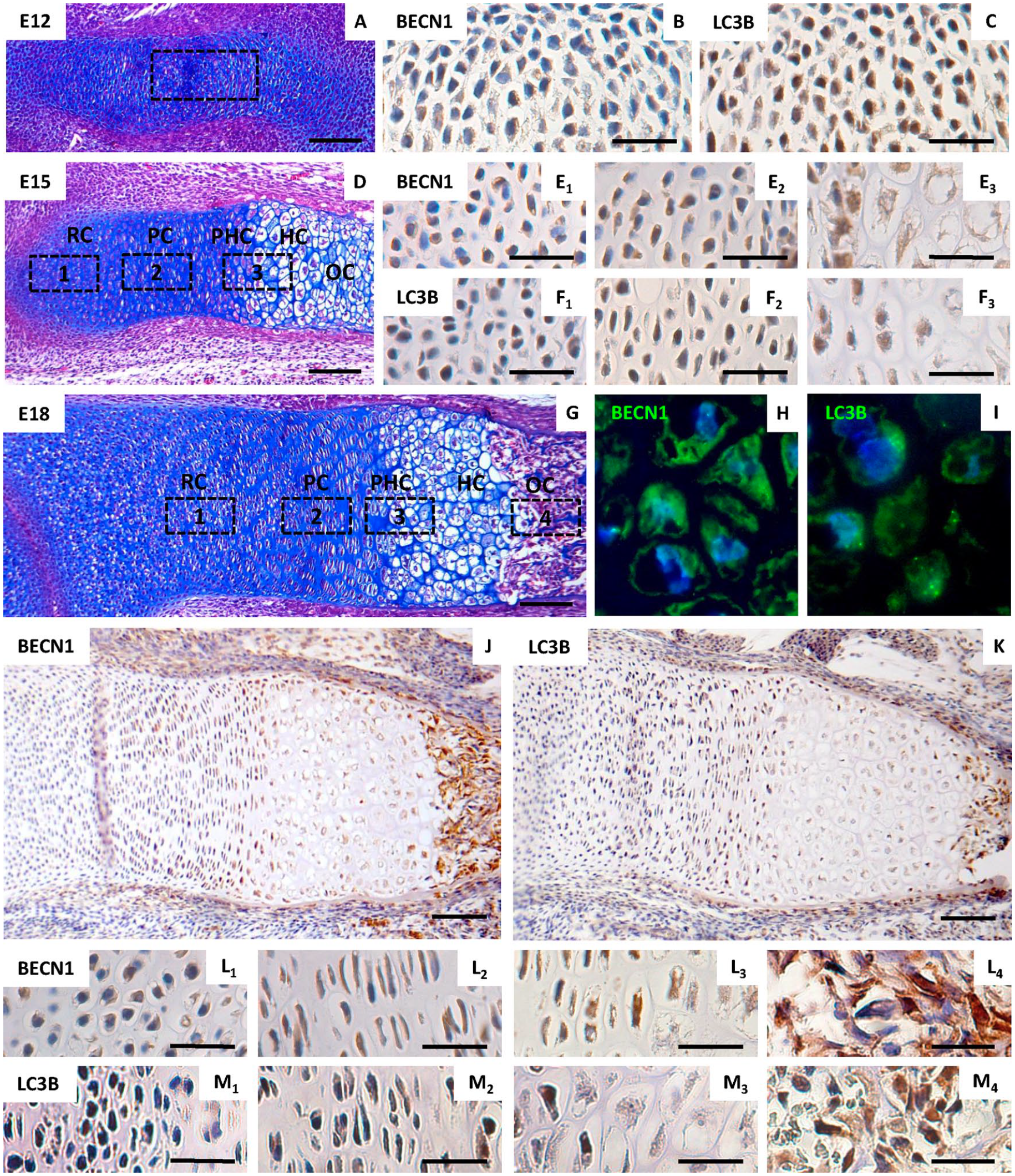
Expression of of Beclin-1 and LC3B in developing prenatal growth plate. Morphological appearance (stained by hematoxylin/eosin/Alcian blue) of forelimb at developmental stage E12 (
Autophagy-Related Molecules Are Expressed During Micromass Progression
A well-established in vitro system of micromass cultures
32
was used to follow spontaneous expression of autophagic molecules during cultivation and to allow for targeted modulation of active caspases. Expression of autophagy related genes was followed during the course of standard micromass cultivation. By comparison of day 2 and day 6 (
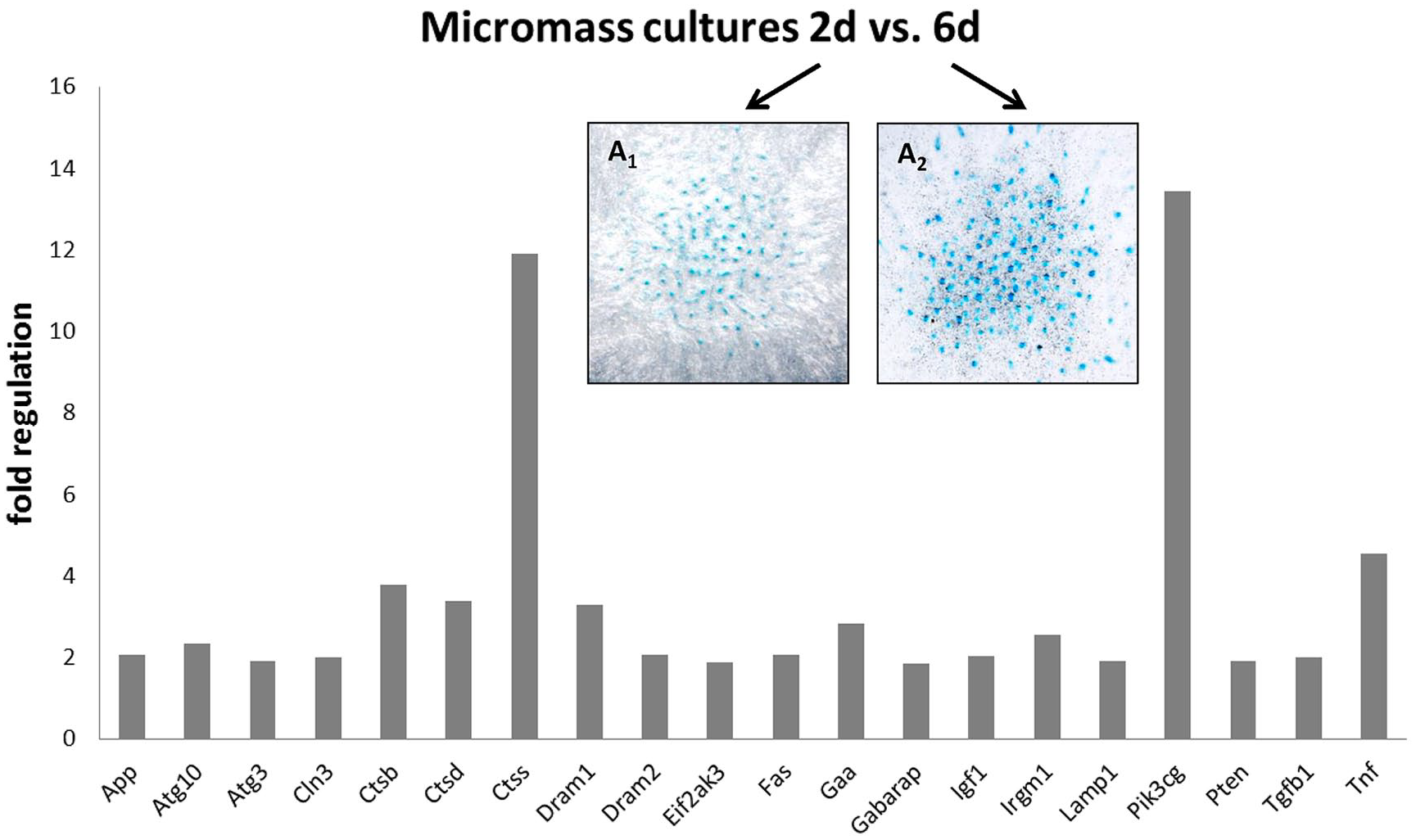
PCR array analysis of autophagy-related gene expression during micromass cultivation from 2 to 6 days (P < .05). Micromass cultures stained by Alcian blue binding to factors of extracellular matrix of cartilage (e.g., mucopolysaccharides) after 2 (
Pro-Apoptotic Caspases Are Activated in Cartilage Nodules in Both Apoptotic and Non-Apoptotic Patterns of Expression
To evaluate the relationship between chondrogenesis, autophagy, and caspase activation, micromass cultures were examined after 6 days of cultivation. By this time point, micromass cultures formed complex structure and formed nodules as documented by visualization of actin filaments (
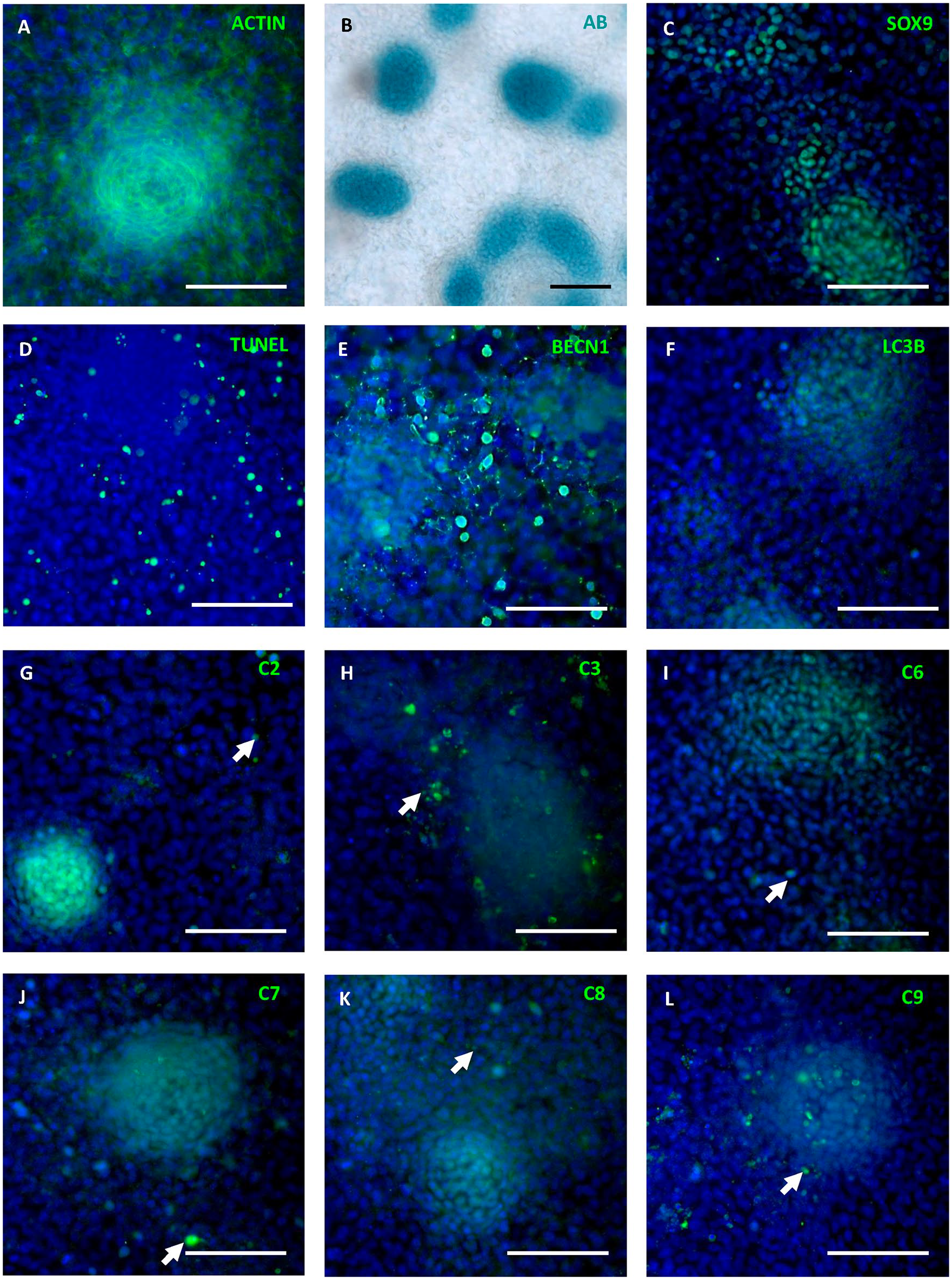
Detection of apoptosis, chondrogenic differentiation, autophagy, and caspase activation in micromass cultures. Alcian blue staining of micromass cultures (
Caspase Inhibition Impacts on Autophagy in Micromass Cultures
To test the role of caspases in regulating autophagy, all caspases were inhibited using a general FMK caspase inhibitor. Compared to control cultures, 6 out of 84 tested genes displayed significant alterations after 6 days of cultivation with the general caspase inhibitor. The most striking change (decrease of 38 times) was shown for Pik3cg. The other affected genes included increased expression of Cdkn2a, Irgm1, Tmem74, Tnfsf10, and decreased expression of Ctss (
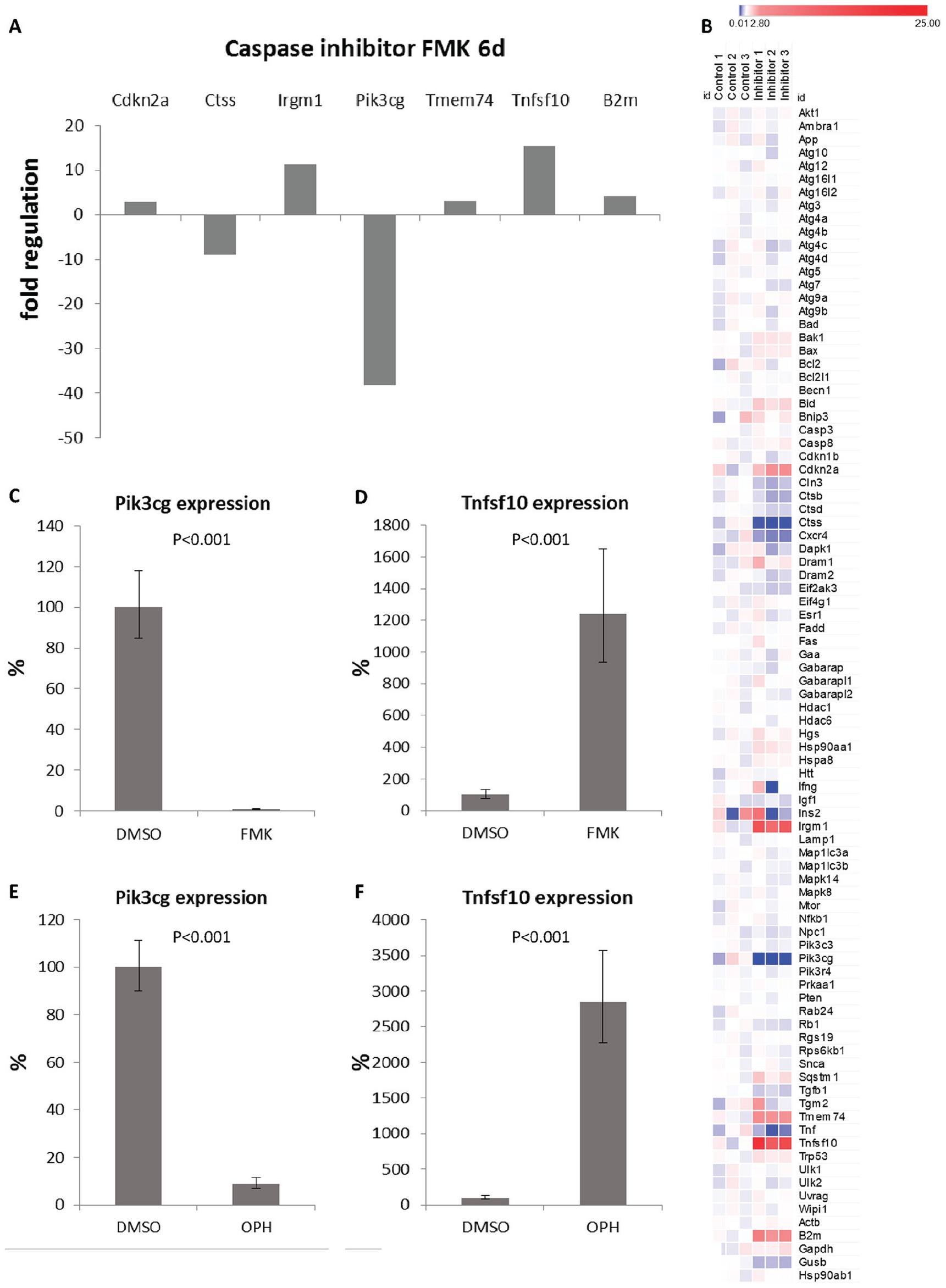
PCR arrays evaluation (
The decrease of Pik3cg in micromass cultures was visible at the protein level after both types of general caspase inhibitors (
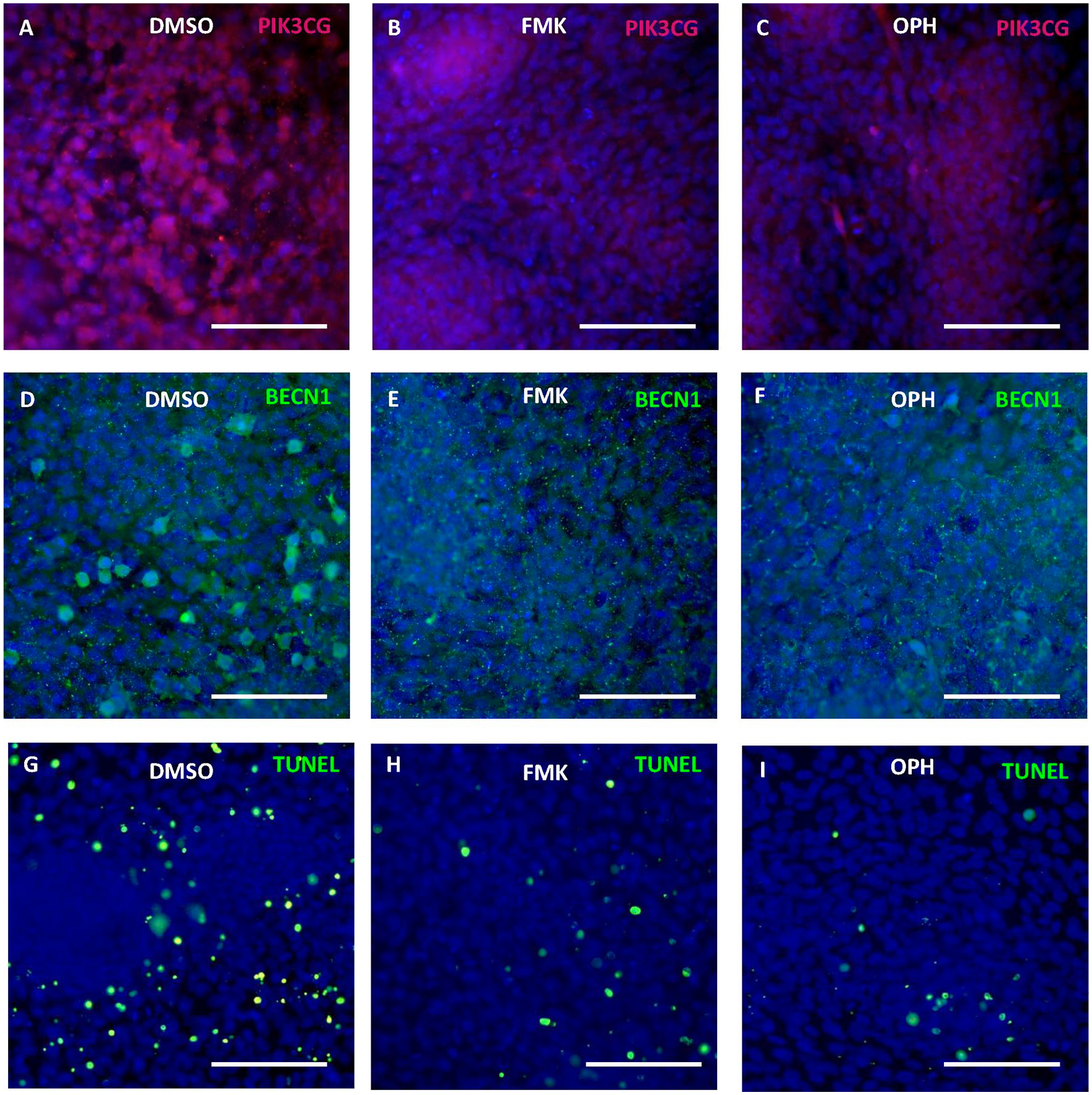
Modulation of apoptosis and autophagy through caspase inhibition. Immunofluorescent detection of Pik3cg and Beclin-1 and apoptosis in micromass cultures after 6 days of cultivation with FMK inhibitor (
To confirm that the changes in expression detected after caspase inhibition were connected with autophagy, we induced autophagy by addition of rapamycin. After 6 days of cultivation, expression of Pik3cg was significantly decreased to 6% (P < 0.001), expression of Tnfsf10 increased to 620% (P < 0.001), and expression of Ctss decreased to 5% (P < 0.001) (
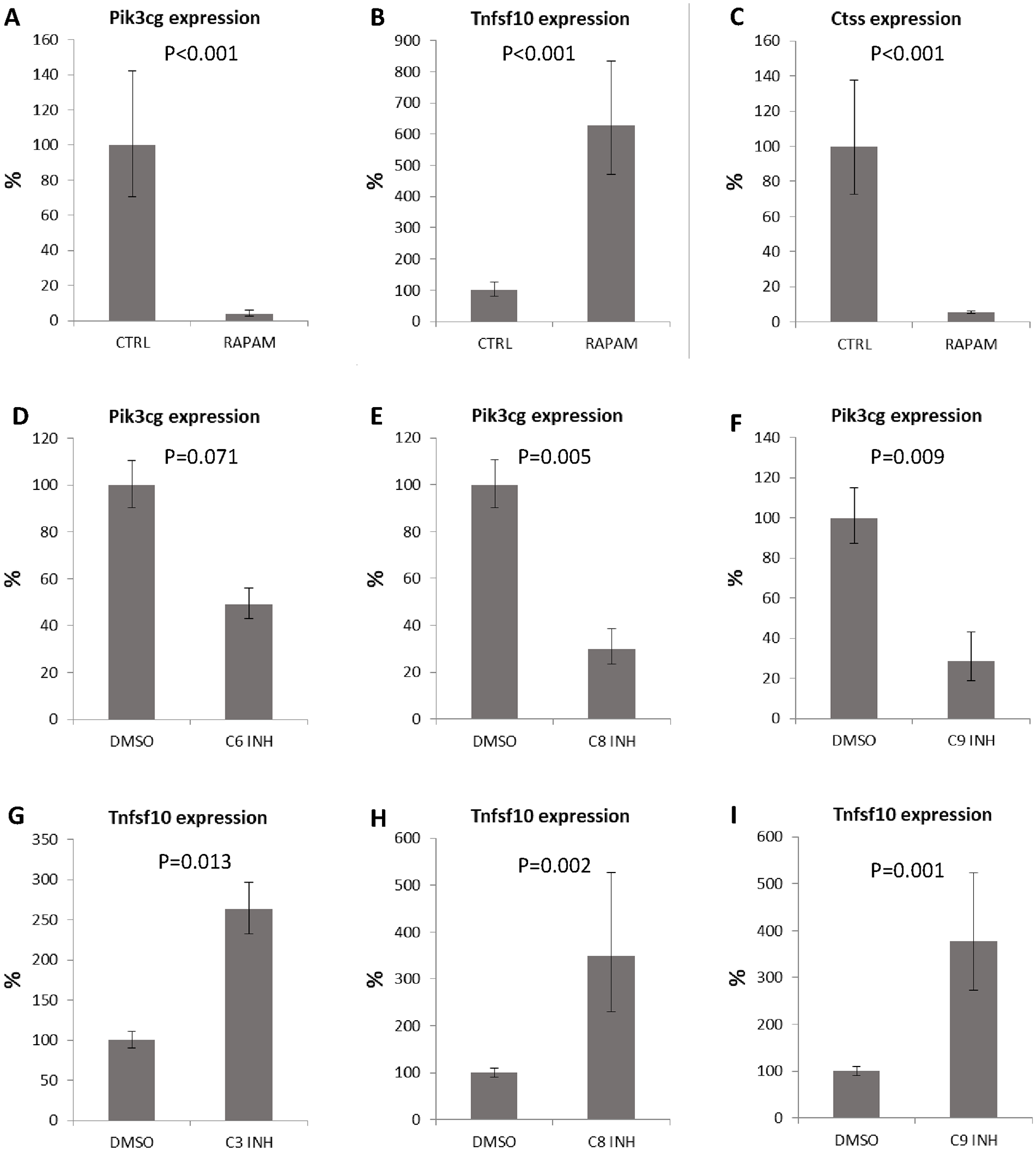
Expression of autophagy related genes after rapamycin treatment and individual caspase inhibition. Expression of Pik3cg (
Individual Pro-Apoptotic Caspase Inhibition Impacts on the Expression of Pik3cg and Tnfsf10
To assess which caspase might be responsible for the most prominent expression changes (such as in Pik3cg and Tnfsf10), caspases-2, -3/7, -6, -8, and -9 were inhibited individually. Unfortunately, due to overlapping substrates, separate pharmacological inhibition of caspase-3 and caspase-7 is not possible. Expression of Pik3cg showed decreasing trend after inhibition of caspase-6 (
Discussion
Apoptosis and autophagy are both engaged in physiologic cell removal, tightly regulated by mutual molecular crosstalk. 36 Novel functions of pro-apoptotic caspases in autophagy have been discussed recently. 23 Since apoptosis and autophagy are associated with long bone growth plate, and caspases are activated in several zones within the growth plate, the presented investigation focused on chondrocytes.
In vivo, autophagy progression is accompanied by the expression of several genes whose specific ablation can cause serious defects in a range of critical developmental steps. 37 Beclin-1 belongs to major markers of autophagy and knockout mice die at E8.5.36,37 We detected Beclin-1 in the prenatal growth plate prominently present in the hypertrophic chondrocytes and in the osteogenic zone. These data correspond with the pattern of Beclin-1 in the postnatal growth plate. 38 In the Beclin-1 positive areas, the expression of pro-apoptotic caspases has been previously described in non-apoptotic cells within the prenatal growth plate. 19 These data might point to caspase-mediated cleavage of Beclin-1, as previously demonstrated for caspase-3. 2 Caspases may thus work as a regulatory switch between autophagy, apoptosis and chondrocyte survival. 23
To follow the expression of autophagy related-genes specifically in chondrocytes, an in vitro micromass culture system was applied. The most dramatic changes observed during the culture period were for the expression of Pik3cg and Ctss. Pik3cg is an intracellular signal transducer enzyme involved in many cellular functions including regulation of autophagy. 39 Ctss is a molecule which has been be connected with apoptosis 40 as well as autophagy. 41
To investigate the effect of caspases on the expression of autophagy genes in chondrocytes, pharmacological inhibitors of caspase-activity were utilized in the micromass cultures. The most significant decrease was detected in expression of Pik3cg and the most apparent increase applied to Tnfsf10. These two markers were further investigated in detail by using inhibitors specific to individual caspases.
For Pik3cg, specific caspase inhibition identified engagement of caspase-8, -9, and probably caspase-6 but not caspase-2. Notably, apoptotic caspases, except caspase-2, are able to cleave Beclin-1 and thus these inhibitors have the potential to decrease autophagy. 23 Here we show that similarly, caspases were also able to regulate levels of Pik3cg expression, an interaction not previously reported. Pik3cg affects autophagy in a negative way by modulation of Akt signaling followed by mTOR activation. 39 This pathway is associated with growth factor mediated processes 42 and FGF signaling was recently suggested as a regulator of bone growth through autophagy. 11
Caspase-8 is one of the key apoptotic components shown to be involved in autophagy 43 and is present also in non-apoptotic chondrocytes, 20 including the growth plate. 19 Inhibition of caspase-8 in micromass culture caused a 3-fold increase in Tnfsf10 (Trail) expression. Trail promotes autophagy by induction of autophagosome formation. 44 Moreover, Trail-mediated signaling can activate caspase-8. The regulation of autophagy by caspase-8 was proposed via its substrates including Atg3, Atg5, and Beclin-1.26,45,46
A similar impact on Tnfsf10 expression was achieved in the case of inhibition of the other apical pro-apoptotic caspase, caspase-9. As for caspase-8, caspase-9, regulation of autophagy is thought to occur via Atg factors (5,7) and Beclin-1.47-49 Compared to caspase-8 and -9, inhibition of caspase-3/7 had a slightly lower effect while there was no significant difference in controls and cultures treated with caspase-2 or caspase-6 inhibitors. Among the executive caspases in autophagy, caspase-6, unlike caspase-3 and -7, can, along with ATGs and Beclin-1, cleave p62, which links the autophagy pathway and the ubiqutin–proteasome system. 50 The effect of pro-apoptotic caspases on expression of Tnfsf10 is here presented for the first time and an understanding of the mechanism may be of high importance also for TRAIL/caspase-based therapies. 51
Among upregulated genes after pan caspase inhibition, significant differences in expression after caspase inhibition were, along with Tnfsf10, observed in the case of Cdkn2a, Irgm1, and Tmem74. Cdkn2a/p16 prohibits progression of cells from G1 phase to S phase and plays a role in the induction of autophagy.52,53 The mechanism of Irgm1 action is not completely known, but Irgm was reported to interact with Ulk1 and Beclin 1 to govern their co-assembly and thus can play a direct role in the core autophagy machinery. 54 Tmem74 has been demonstrated to co-localize with apoptosis-related protein Bik in subcellular organelles and inhibit Bik-induced apoptosis. 55
Notably, there was an increased expression of B2m, beta-2-microglobulin, one of the housekeeping genes. Despite the fact that this gene is not considered in the panel of autophagy-related molecules, anti-B2m antibodies as apoptosis enhancers were recently demonstrated to inhibit autophagy. 56 Additionally, B2m may contribute to pathogenesis of osteoarthritis. 57
Knowledge about mechanisms of caspases engagement in autophagy is not yet complete. Z-FMK pan caspase inhibitors were earlier reported as modulators of autophagy 58 but the mechanism was explained particularly by inhibition of Beclin-1 cleavage and keeping retention of its autophagic properties. 24 Beclin-1 cooperates with Pik3 molecules in the formation of autophagosomes. 39 During chondrocytic cultivation, mRNA expression of Beclin-1 was kept at high levels, which was maintained even after inhibition of caspases ( Table 1 ). In addition to autophagy, Beclin-1 plays an important role during engulfment of apoptotic cells, since the process was abolished in Beclin-1 knockout cells. 59 This may explain why some Beclin-1-positive cells clearly apparent in the control cultures are almost not visible after caspase inhibition. This Beclin-1-positive cell population displays phosphatidylserine translocation rather than TUNEL positivity, indicating that they are probably at the beginning of the apoptotic process. Caspase inhibition therefore resulted in a significant reduction of apoptosis and thus possibly disruption of Beclin-1 participation during apoptotic cell engulfment.
Autophagy-Related Genes Expressed at High Levels (Same Range as Housekeeping Genes of the Panel) After 6 Days of Cultivation of Micromass Cultures.
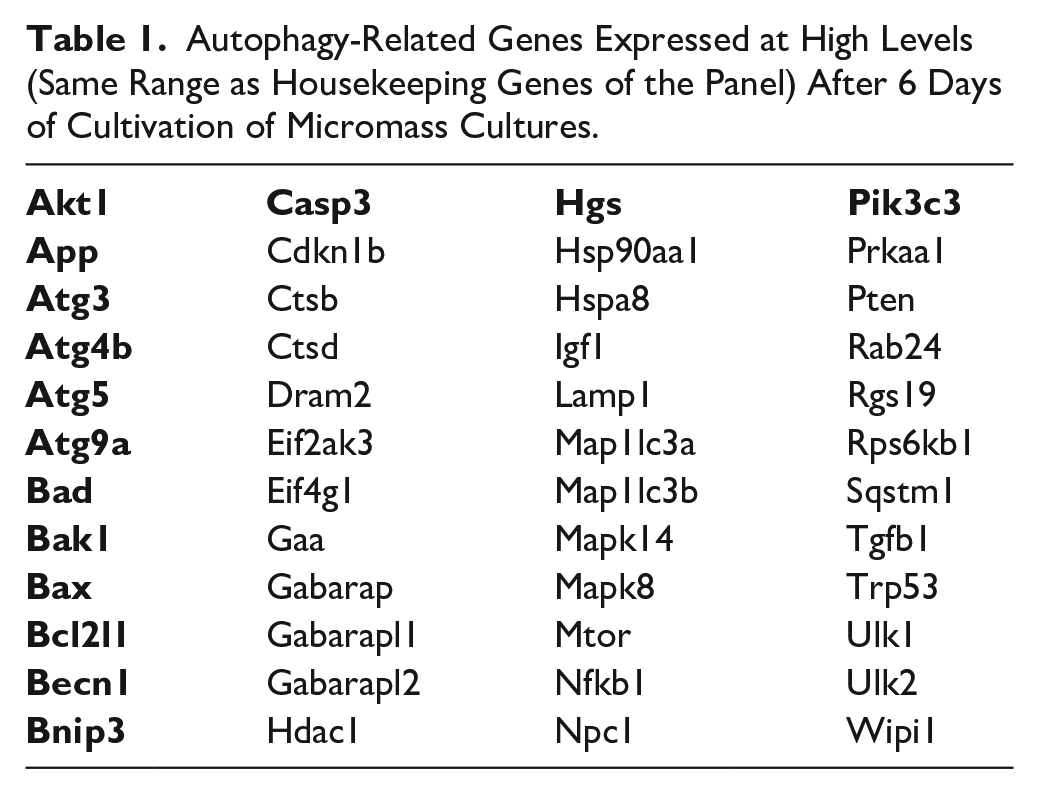
The findings are in agreement with the fact that the decision point regulated by Beclin-1 happens at the protein level as mRNA of Becn1 was constantly high. In addition, the impact of caspase inhibition on the transcriptome must be mediated by protein changes.
Importantly, the major changes in gene expression triggered by inhibition of caspases were mimicked by stimulation of autophagy using rapamycin. Inhibition of caspases in chondrocytes therefore leads to stimulation of autophagy. Increased levels of Trail after caspase inhibition and after rapamycin treatment also supports a hypothesis that caspases can regulate autophagy through expression of Trail.
In the current investigation, novel pieces of the network puzzle connecting apoptosis and autophagy by caspases were revealed. Although autophagy was already described in cartilage, this is the first time when caspases were identified as regulators of autophagy related genes in chondrocytes as a part of physiological cartilage development. Furthermore, several novel caspase downstream factors were identified here. The results were confirmed by two different types of general caspase inhibitors (FMK and OPH) and compared to changes after rapamycin treatment. Separate inhibition of specific caspases allowed for identification of individual pro-apoptotic caspases responsible for the major effects of the inhibition. Notably, several genes whose expression was modulated by caspase inhibition are of importance in diagnostics and clinically oriented studies focused on osteoarthritis. 60 With regard to the critical function of autophagy for metabolism of chondrocytes, caspases might be potentially engaged in repair of the chondrocyte damage or slowing down of their aging. Thus, these data open new possibilities for cartilage reparation.
Supplemental Material
19-5-2020_Supplementary_figure_1 – Supplemental material for Caspase Inhibition Affects the Expression of Autophagy-Related Molecules in Chondrocytes
Supplemental material, 19-5-2020_Supplementary_figure_1 for Caspase Inhibition Affects the Expression of Autophagy-Related Molecules in Chondrocytes by Barbora Vesela, Eva Svandova, Alice Ramesova, Adela Kratochvilova, Abigail S. Tucker and Eva Matalova in CARTILAGE
Footnotes
Supplemental Material
Author Contributions
EM designed the study. BV planned experiments. BV, AR, ES, and AK performed experiments. ES and AT analyzed the data. BV, EM, and AT wrote the paper.
Acknowledgment and Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The research was granted by the Czech Science Foundation, Project GA ČR 19-12023S. The CR-UK cooperation was supported by the Inter-COST project LTC18 (“Caspases as Novel Regulators in Osteogenic Cellular Networks,” running under the COST Action CA15214), the Inter-Excellence Programme provided by the Ministry of Education of the Czech Republic (![]() ).
).
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethics Approval
Ethical approval was not sought for the present study. The project GACR 19-12023S was approved and supervised by the Czech Sciences Foundation including all ethics issues.
Animal Welfare
The present study followed international, national, and/or institutional guidelines for humane animal treatment and complied with relevant legislation.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.