
Abstract
Objective
The production of extracellular matrix is a necessary component of articular cartilage repair. Gene transfer is a promising method to improve matrix biosynthesis by articular chondrocytes. Gene transfer may employ transgenes encoding regulatory factors that stimulate the production of matrix proteins, or may employ transgenes that encode the proteins themselves. The objective of this study was to determine which of these 2 approaches would be the better choice for further development. We compared these 2 approaches using the transgenes encoding the structural matrix proteins, aggrecan or type II collagen, and the transgene encoding the anabolic factor, insulin-like growth factor I (IGF-I).
Methods
We transfected adult bovine articular chondrocytes with constructs encoding type II collagen, aggrecan, or IGF-I, and measured the expression of type II collagen (COL2A1) and aggrecan (ACAN) from their native genes and from their transgenes.
Results
IGF-I gene (IGF1) transfer increased the expression of the native chondrocyte COL2A1 and ACAN genes 2.4 and 2.9 times control, respectively. COL2A1 gene transfer did not significantly increase COL2A1 transcripts, even when the transgene included the genomic COL2A1 regulatory sequences stimulated by chondrogenic growth factors. In contrast, ACAN gene transfer increased ACAN transcripts up to 3.4 times control levels. IGF1, but not ACAN, gene transfer increased aggrecan protein production.
Conclusion
Taken together, these results suggest that the type II collagen and aggrecan production required for articular cartilage repair will be more effectively achieved by genes that encode anabolic regulatory factors than by genes that encode the matrix molecules themselves.
Introduction
Articular cartilage damage is a major cause of pain and disability due to trauma and osteoarthritis. 1 Articular chondrocytes have a limited capacity for restoring the cartilage matrix that serves as the gliding surface for joints. As a result, cartilage damage is generally persistent and progressive. 2 Current treatments for joint trauma and osteoarthritis address the symptoms of the disease. Treatments are needed that can address the cartilage loss itself. The restoration of matrix molecules in the articular cartilage will be a necessary feature of any treatment for articular cartilage damage. Various polypeptide growth factors have been identified that promote matrix production by articular chondrocytes. In vitro and in vivo studies suggest that some of these factors have therapeutic potential for articular cartilage repair.3 -13
The effort to deliver anabolic growth factors together with cells to restore lost cartilage tissue has led to the application of gene transfer to cell-based tissue repair. One approach to such gene therapy is indirect; it delivers genes that encode regulatory molecules to increase the expression of chondrocyte genes that encode the desired matrix proteins. A second approach directly delivers the genes that encode the desired matrix proteins. Multiple studies have shown that delivery of genes that encode regulatory molecules, such as growth factors, can stimulate the production of new matrix by articular chondrocytes.14 -20 To our knowledge, no studies have investigated the delivery of the genes that encode the matrix molecules themselves. This direct approach offers theoretical advantages for articular cartilage repair. First, it avoids nonspecific growth factor actions, including stimulation of undesired molecules such as type I collagen. Second, it prevents off-target effects on noncartilage tissues such as synovium. Third, it avoids the potential risk of uncontrolled growth factor–induced cell proliferation.
Articular cartilage matrix is composed primarily of type II collagen and aggrecan. The production of both molecules by articular chondrocytes is regulated, in large measure, by polypeptide growth factors. Prominent among these is insulin-like growth factor I (IGF-I). Delivery of IGF-I as an exogenous protein, or by gene transfer, stimulates the gene expression of both type II collagen and aggrecan in vitro 21 and the production of new cartilage matrix containing type II collagen and aggrecan in vitro and in vivo.5,6,12,14,16 -18,22 -24 For these reasons, we chose genes encoding type II collagen (COL2A1) and aggrecan (ACAN) as the structural matrix genes, and the gene encoding IGF-I (IGF1) as the cell-regulatory gene for these studies.
Type II procollagen is produced in 2 forms from 2 mRNA splice variants of the COL2A1 gene. The type IIA (COL2A1A) variant contains the full sequence, including exon 2 and is synthesized by chondroprogenitor cells. The type IIB (COL2A1B) variant lacks exon 2 and is produced by mature chondrocytes. 25 Type II collagen is responsible for the tensile strength of articular cartilage and its loss during osteoarthritis is associated with advancing disease. Aggrecan is a high molecular mass (~2 × 106 Da) proteoglycan composed of a core protein modified by the addition of glycosaminoglycan chains. 26 When bound to hyaluronic acid, aggrecan is responsible for the compressive strength of articular cartilage and its loss is a hallmark of early osteoarthritis. Both type II collagen and aggrecan are synthesized by articular chondrocytes and serve as markers of the chondrocyte phenotype.
Despite their importance as the principal structural components of articular cartilage, it is not known whether the use of the genes encoding these matrix molecules is an effective approach to increasing aggrecan and type II collagen gene expression. Furthermore, gene transfer may, instead, employ transgenes encoding regulatory factors that stimulate the expression of these matrix molecule genes. The objective of this study was to test the hypothesis that 1 of these 2 approaches is superior to the other as a candidate for further development of gene therapy for articular cartilage repair.
Method
Vector Preparation
Adeno-associated virus-based plasmid vectors were created to deliver cytomegalovirus (CMV)-driven genes encoding human IGF-I, type IIA procollagen, type IIB procollagen, and aggrecan. These vectors are designated pAAV-IGF-I, pCMV-COL2A1A, pCMV-COL2A1B, and pCMV-ACAN respectively. In addition, a vector containing the human COL2A1 coding sequence and its native regulatory elements, designated pNR-COL2A1, was created to enable the expression of human type II procollagen (both type IIA and type IIB) that is regulated by the native promoter and enhancer of the human COL2A1 gene ( Fig. 1 ) (Detailed methods in Supplementary Material).
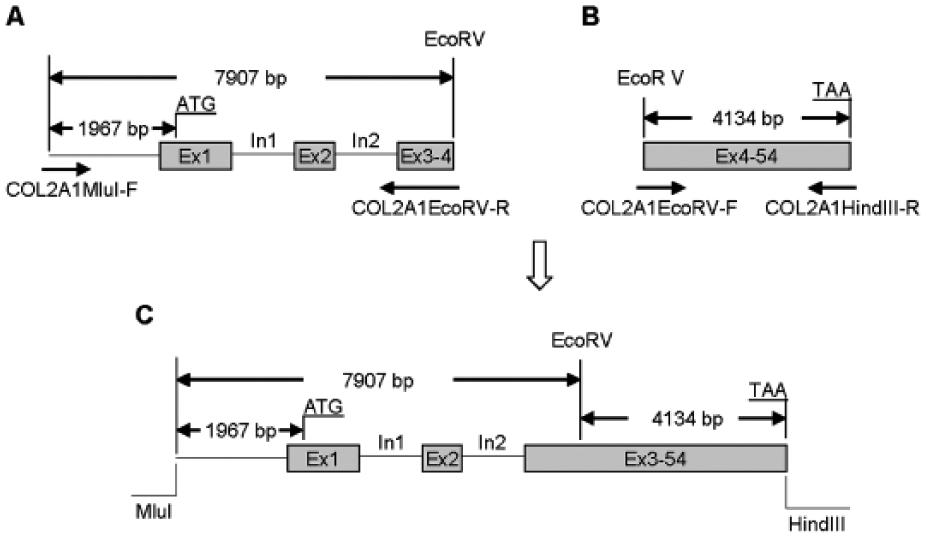
Schematic illustration of the construction of the genomic type II collagen transgene (NR-COL2A1). (
Chondrocyte Cell Culture and Transfection
Basal medium was prepared with Dulbecco’s modified Eagle medium, 100 U/mL penicillin, 100 μg/mL streptomycin, 2 mM glutamine (Invitrogen, Carlsbad, CA) and 50 μg/mL ascorbic acid (Sigma, St. Louis, MO). Complete medium was prepared with basal medium and 10% fetal bovine serum. Bovine articular chondrocytes were isolated as previously described. 27 Briefly, carpal articular cartilage was harvested from skeletally mature (growth plates closed) bovids. Chondrocytes were isolated and placed in primary monolayer in 6-well plates in complete medium. After 3 days of culture, the medium was replaced with fresh complete medium and the cells were transfected using 6 µL FuGENE 6 (Roche Applied Science, Indianapolis, IN) with 2 µg of each plasmid DNA per well. After 16 hours, transfection was stopped by replacing the medium with fresh complete medium. On days 2, 4, or 6 following transfection, cultures were terminated or given fresh basal medium. At the time of termination, the cell layer was lysed in RLT lysis buffer (RNeasy Mini kit, Qiagen, Germantown, MD) and the samples submitted for total RNA purification and real-time polymerase chain reaction (PCR) analysis.
Stimulation of the Genomic Type II Collagen Transgene with Exogenous Growth Factor Proteins
To stimulate the expression of the genomic NR-COL2A1 transgene with chondrogenic growth factor(s), the medium was replaced with fresh complete medium supplemented with 200 ng/mL IGF-I (Peprotech, Rocky Hill, NJ) or the combination of 200 ng/mL IGF-I, 100 ng/mL bone morphogenetic protein-2 (BMP-2) (R&D Systems, Minneapolis, MN), and 100 ng/mL bone morphogenetic protein-7 (BMP-7) (R&D Systems, Minneapolis, MN), and the cells were transfected as above with pNR-COL2A1 or empty vector. This growth factor combination was selected based on prior studies demonstrating synergistic stimulation of both COL2A1 and ACAN gene expression when the genes encoding these growth factors were delivered to adult bovine articular chondrocytes. 21 After 16 hours, the transfection was stopped by replacing the medium with fresh complete medium supplemented with the same growth factor(s). On days 2, 4, or 6 following transfection, cultures were terminated or given fresh basal medium containing 0.1% bovine serum albumen (Sigma) and the same growth factor(s). Chondrocytes transfected by empty vector without growth factor treatment served as control.
RNA Purification, Reverse Transcription, and Real-Time PCR Analysis
RNA purification and reverse transcription were performed as previously described. 27 Briefly, total RNA was prepared using the RNeasy Mini kit (Qiagen, Germantown, MD). On-column DNase digestion was performed to remove any residual DNA. Reverse transcription was performed using the High-capacity cDNA Archive Kit (Applied Biosystems, Foster City, CA). Reverse transcription was terminated by heating at 95°C for 20 minutes. cDNA samples were diluted 1:20 for real-time PCR analysis.
To distinguish and compare expression of the chondrocytes’ bovine native COL2A1 gene from that of the human COL2A1 transgenes, two different sets of real-time PCR primers were designed to quantify the transcripts from these 2 gene sources: (1) primers: HB-COL2A1-F and HB-COL2A1-R, which detect both human and bovine COL2A1 mRNA and (2) primers: H-COL2A1-F and H-COL2A1-R, which detect only human COL2A1 mRNA. Similarly, 2 different sets of real-time PCR primers were designed to distinguish and compare expression of the chondrocytes’ bovine native ACAN gene from that of the human ACAN transgene. HB-ACAN primers, HB-ACAN-F and HB-ACAN-R, detect both human and bovine ACAN mRNA while H-ACAN primers, H-ACAN-F and H-ACAN-R, detect only human ACAN mRNA ( Table 1 ).
Primers Used for PCR and Real-Time PCR.

PCR = polymerase chain reaction; F = forward; R = reverse; HB = human and bovine; H = human.
Human ACAN mRNA, total ACAN mRNA including both human and bovine ACAN mRNA, human COL2A1 mRNA, and total COL2A1 mRNA including both human and bovine COL2A1 mRNA, were measured using SYBR Green Master Mix (Applied Biosystems, Foster City, CA) and specific primer sets described above, respectively. The standard curve method was used to calculate the expression of target genes. The standard curves were prepared using 10× serial dilutions of pCMV-ACAN and pCMV-COL2A1A plasmids respectively. The concentration of plasmid DNA was measured by A260 and converted the absolute number of copies according to the size of the plasmid. Data are expressed as times change in the number of transcripts compared with control after normalization to bovine 18S rRNA levels. Bovine 18S rRNA was measured by real-time PCR as previously described. 21
Glycosaminoglycan Assay
Glycosaminoglycan content of the cells and surrounding matrix was measured by dimethylmethlyene blue (DMMB) assay as previously described. 20
Statistical Analysis
Cell isolates from 3 different animals (N = 3) were used in 3 independent experiments. Analyses of COL2A1 and ACAN mRNA (dependent variables) were performed on the ratios of expression by cells transfected with the designated transgenes to expression by cells transfected with empty vector (controls). One-sample t tests were used to determine the significance of the ratio to the control for each group. Group and day effects were evaluated using mixed-model analyses of variance (ANOVAs) with fixed effects for group, day, and their interaction and a random effect to correlate samples over time. All analyses were performed after a natural log transformation of the data to satisfy the normality assumption for the ANOVA.
Results
Regulation of Type II Collagen Gene Expression by IGF-I Gene Transfer
In this model, the cells transfected with empty vector, or pAAV-IGF-I, express only the native bovine COL2A1 gene. This expression is represented by real-time PCR measurement using HB-COL2A1 primers. Articular chondrocytes carrying IGF1 transgenes increased COL2A1 mRNA from the native bovine COL2A1 gene up to 2.4 times (P < 0.01) stimulation compared with mock-transfected chondrocytes. Expression peaked at 4 days and remained elevated at the 6-day end-point of the experiments ( Fig. 2 , Set 2).
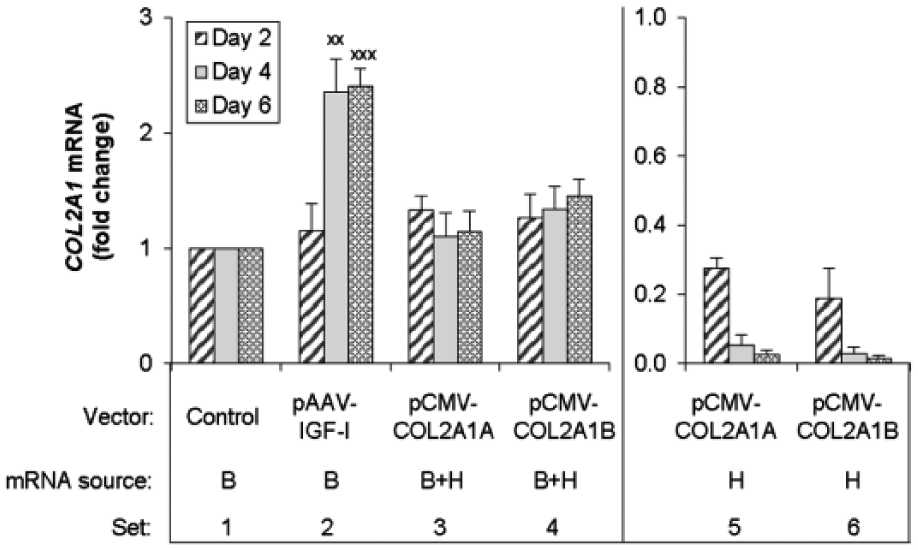
Effect of the transgene encoding human IGF-I, type IIA procollagen, or type IIB procollagen on COL2A1 transcripts. Chondrocytes were transfected with the designated vectors, or empty vector (Control), and cultured for the designated time periods. The number of COL2A1 transcripts was measured by real-time polymerase chain reaction (PCR) using species-specific primers as described in Materials and Methods. The source of mRNA was the bovine native gene (B), both the bovine native gene and the human transgenes (B + H) or the human transgenes only (H). Data are expressed as times change in the number of COL2A1 transcripts compared with the control after normalization to bovine 18S rRNA levels. The y-axis scale for Sets 1-4 is larger than that for Sets 5 and 6, reflecting a much greater abundance of transcripts from the native bovine gene than from the human transgenes. Data represent the mean of 3 independent experiments. Data are expressed as mean ± SD of times change in transcript number compared with control.
The Type II Collagen Native Gene and Transgene Are Differentially Expressed
The cells transfected with pCMV-COL2A1A or pCMV-COL2A1B, express both the bovine native COL2A1 gene and the human COL2A1 transgene. The total transcripts from both the human transgene and the bovine native gene are represented by real-time PCR measurement using the HB-COL2A1 primers. The transcripts from the human COL2A1 transgene were measured by real time PCR using the human-specific H-COL2A1 primers.
Transfection with pCMV-COL2A1A or pCMV-COL2A1B did not increase total COL2A1 transcripts compared to mock transfected controls at any time point tested ( Fig. 2 , Sets 3 and 4). By comparing the HB-COL2A1 primer data (total transcripts) with H-COL2A1 primer data (human transcripts), we found that the number of human transcripts represented less than 21% of the total COL2A1 transcripts in pCMV-COL2A1A or pCMV-COL2A1B transfected cells at day 2, and the expression of these transgenes declined faster than that of the native bovine COL2A1 gene. As a result, by the end of culture at 6 days, less than 3% of total COL2A1 transcripts were generated from human COL2A1 transgenes ( Fig. 2 , Sets 3 and 5, Sets 4 and 6).
Growth Factors Differentially Stimulate Type II Collagen Expression from the Native Type II Collagen Gene and from a Human Type II Collagen Transgene Driven by Native Regulatory Elements
A theoretical advantage of the human genomic DNA vector, pNR-COL2A1, is the inclusion of the native regulatory sequences of the human COL2A1 gene through which growth factors may stimulate transgene expression. Because BMP-2 and BMP-7 have been shown to interact with IGF-I to synergistically stimulate bovine articular chondrocyte expression from its native COL2A1 gene, 21 we employed the three factors together to test the hypothesis that NR-COL2A1 transgene expression can be augmented by these chondrogenic growth factors. Articular chondrocytes carrying the NR-COL2A1 transgenes were treated with 200 ng/mL IGF-I alone or in combination with 100 ng/mL each of BMP-2 and BMP-7. To compare the regulation of the NR-COL2A1 transgenes with that of the native bovine COL2A1 gene, we also delivered these growth factors to control chondrocytes transfected by the empty vector.
In control cells, data from HB-COL2A1 primers showed that treatment with IGF-I progressively increased native bovine COL2A1 expression in a time-dependent fashion to nearly 3.5 times control (P < 0.001) at 6 days of culture ( Fig. 3 , Set 2). The combination [IGF-I + BMP-2 + BMP-7] augmented this stimulation to 13.7 times (P < 0.001) ( Fig. 3 , Set 3).
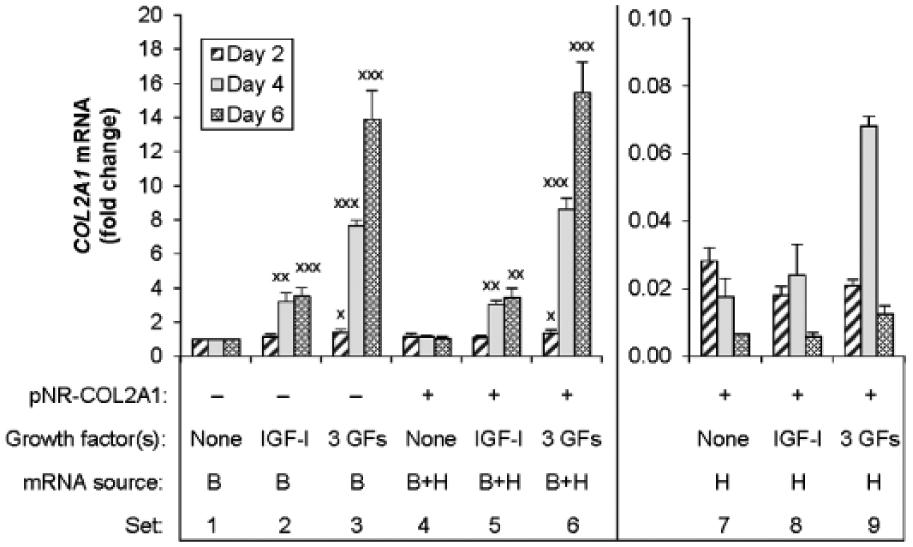
Effect of the transgene encoding human type II procollagen under the control of native regulatory elements on COL2A1 transcripts. Chondrocytes were transfected with pNR-COL2A1 (
In pNR-COL2A1 transfected cells, the data from HB-COL2A1 primers showed that the addition of NR-COL2A1 transgenes did not increase total COL2A1 transcripts compared with its control ( Fig. 3 , Sets 1 and 4). The data also showed that NR-COL2A1 did not increase total COL2A1 transcripts compared to the cells that were treated with IGF-I ( Fig. 3 , Sets 2 and 5) or the combination, [IGF-I + BMP-2 + BMP-7] ( Fig. 3 , Sets 3 and 6). Comparing the data from HB-COL2A1 primers (total transcripts) with that using H-COL2A1 primers (human transcripts) showed that expression from the NR-COL2A1 transgenes was approximately an order of magnitude less at all time points than expression from the native bovine gene, with or without growth factor stimulation ( Fig. 3 , Sets 7-9).
To determine whether IGF-I or [IGF-I + BMP-2 + BMP-7] stimulates NR-COL2A1 transgene expression via its native regulatory elements, the data from H-COL2A1 primers were normalized to the expression from pNR-COL2A1 transfection without growth factors. The results showed that IGF-I transiently increased the expression of the NR-COL2A1 transgenes 1.4 times control (P < 0.01) at day 4, and the 3 growth factors in concert increased this expression 4.1 times (P < 0.01) and 1.9 times (P < 0.05) at days 4 and 6, respectively (Supplementary Table S1). These data indicate that the NR-COL2A1 transgenes are responsive to these chondrogenic growth factors, but that the magnitude of transgene stimulation is much less than that of the native gene. Taken together, the data indicate that the contribution of the transgenes to the total COL2A1 transcripts is minimal, even in the presence of highly stimulatory growth factors.
The Aggrecan Transgene and Native Aggrecan Gene Are Differentially Expressed
We distinguished expression of the bovine chondrocytes’ native ACAN gene from that of the human ACAN transgenes by using 2 different real-time PCR primer sets. Data from the HB-ACAN primers represent the total transcripts from both the human ACAN transgenes and the bovine native ACAN gene. Data from the human-specific H-ACAN primers represent transcripts from the human ACAN transgene.
Articular chondrocytes carrying the IGF1 transgenes increased ACAN mRNA from the native bovine ACAN gene 3.0 times (P < 0.001) and 2.6 times (P < 0.001) compared with control chondrocytes at days 4 and 6, respectively ( Fig. 4 ). Delivery of human ACAN transgenes increased total (human + bovine) ACAN transcripts 2.5 times (P < 0.001), 4.7 times (P < 0.001), and 2.1 times (P < 0.01) compared with controls at days 2, 4, and 6, respectively ( Fig. 4 ). By comparing the HB-ACAN primer data (total transcripts) with H-ACAN primer data (human transcripts), we found that the human ACAN transgenes generated a similar number of ACAN transcripts as the bovine native ACAN gene at days 2 and 6, and 3.4 times as many transcripts as the bovine native ACAN gene at day 4 ( Fig. 4 ). Furthermore, chondrocytes treated with human ACAN transgene produced 2.1 times (P < 0.001) and 1.6 times (P < 0.01) more ACAN transcripts at days 2 and 4 respectively, than chondrocytes treated with IGF1 transgenes ( Fig. 4 ). Thus, unlike type II collagen gene transfer, aggrecan gene transfer significantly increased the number of transcripts compared to the respective bovine native genes alone. The data further show that aggrecan gene transfer generated more ACAN transcripts than did stimulation of the native ACAN gene by IGF-I gene transfer.
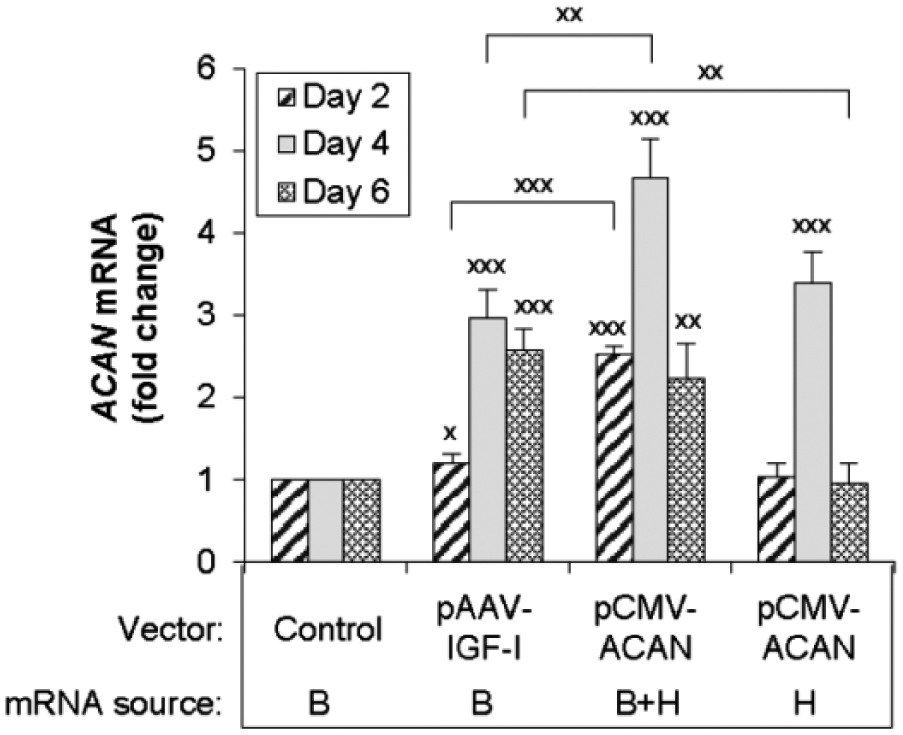
Effect of the transgene encoding human IGF-I or aggrecan on ACAN transcripts. Chondrocytes were transfected with the designated vectors, or empty vector (Control) and cultured for the designated time periods. The number of ACAN transcripts was measured by real-time polymerase chain reaction (PCR) using species-specific primers as described in Materials and Methods. The source of mRNA was the bovine native gene (B), both the bovine native gene and the human transgenes (B + H) or the human transgenes only (H). Data are expressed as times change in the number of ACAN transcripts compared with the control after normalization to bovine 18S rRNA levels. Data represent the mean of 3 independent experiments. Data are expressed as mean ± SD of times change in transcript number compared with control.
Glycosaminoglycan Production
The glycosaminoglycan content of the cells and surrounding matrix did not differ significantly among control cells treated with empty vector and those treated with the ACAN transgene. In contrast, cells treated with the IGF1 transgene produced 2.2 times (P < 0.001) more glycosaminoglycan than did control cells ( Fig. 5 ). These data indicate that the increase in ACAN transcripts that occurred in response to ACAN gene transfer was not reflected in a commensurate increase in aggrecan in the cells or matrix.
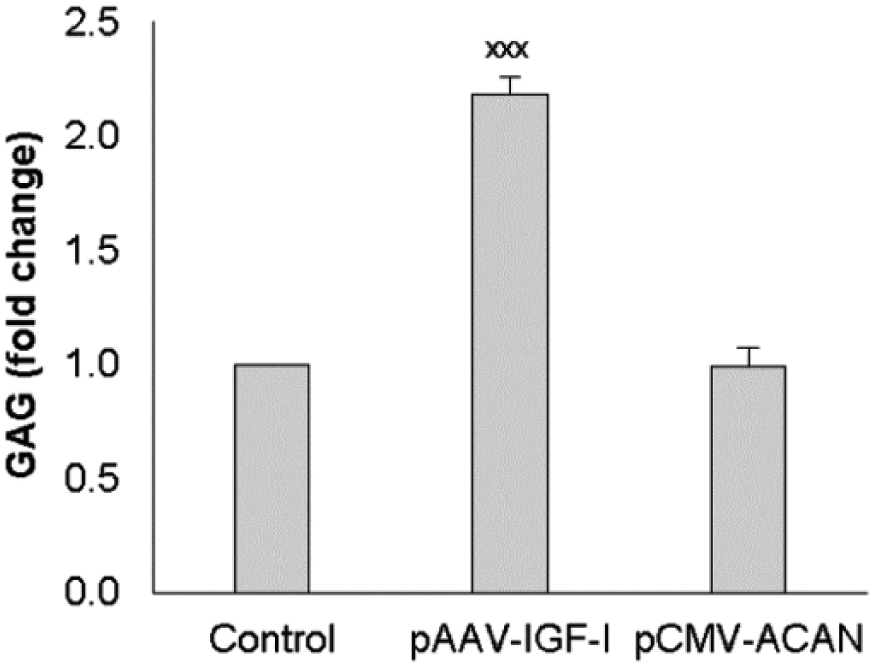
Effect of the transgene encoding human IGF-I or aggrecan on glycosaminoglycan production (GAG). Chondrocytes were transfected with the designated vectors, or empty vector (Control). After 16 hours, transfection was stopped by replacing the medium with fresh complete medium. On days 2 and 4 after transfection, the medium was replaced by basal medium. On day 6 after transfection, culture was terminated and the cell layer was digested with proteinase k after the medium was removed. GAG in cell layer was measured by dimethylmethlyene blue (DMMB) assay of proteinase k cell layer digest. Data are expressed as fold change compared with the control. Data represent the mean ± SD of 3 independent experiments.
Discussion
These studies address a fundamental question regarding the selection of therapeutic genes for cartilage repair: whether the production of the cartilage matrix required for cartilage repair is better achieved by gene therapy employing transgenes that encode cartilage structural proteins, or alternatively, by gene therapy employing transgenes that encode factors that regulate these structural proteins’ native genes. In the case of type II collagen, the data indicate that using growth factor transgenes to augment expression from its native gene is considerably more effective than increasing the number of gene copies to be transcribed. In the case of ACAN expression, treatment by aggrecan gene transfer is more effective than treatment by IGF1 gene transfer in increasing ACAN transcripts in chondrocytes ( Fig. 4 ).
The large difference in the number of transcripts generated from the chondrocytes’ native bovine COL2A1 and ACAN genes compared with their respective human transgenes may reflect, in part, the high baseline levels of expression from the native COL2A1 gene and the relatively lower levels of expression from the native ACAN gene (Supplementary Table S2). Thus, the transgenes contributed a much smaller proportion of the total COL2A1 transcripts than of the total ACAN transcripts. Even though IGF-I and the combination [IGF-I + BMP-2 + BMP-7] stimulated the expression of the NR-COL2A1 transgenes driven by native regulatory elements, this stimulation was minimal compared to the stimulation of the native COL2A1 gene. Even though the NR-COL2A1 transgene contains the native promoter before Exon 1 and putative enhancer in intron 1 ( Fig. 1C ), it may be that other regulatory elements are also involved in the native gene. An enhancer element has recently been identified in intron 7 of the type II collagen gene, 28 and this is not present in the plasmid pNR-COL2A1 ( Fig. 1C ). In addition, the length of the COL2A1 and ACAN transgenes could limit their effectiveness, because comparatively fewer of them can be delivered in a given amount of plasmid DNA (2 µg in these studies) than for most transgenes, including the IGF1 transgene. However, increasing the number of transgenes by increasing plasmid DNA from 2 µg to 4 µg did not significantly increase the number of either COL2A1 or ACAN transcripts (data not shown), further suggesting that multiple mechanisms regulate transcription from these transgenes.
A limitation of this study is its focus on just the type II collagen and aggrecan genes. These genes were selected because they encode the predominant structural components of articular cartilage. The genes encoding the variety of lower abundance molecules present in cartilage matrix may behave differently when transferred to chondrocytes. An additional limitation of the study is its focus on IGF-I. IGF-I was selected for its unusual combination of anabolic and anticatabolic effects on cartilage matrix homeostasis by articular chondrocytes,6,22,23 including stimulation by IGF1 gene transfer of COL2A1 and ACAN expression under similar experimental conditions. 21
Similarly, the combination of exogenous proteins, [IGF-I + BMP-2 + BMP-7] was selected to stimulate the genomic NR-COL2A1 transgene because this combination of growth factor genes generated the maximal stimulation of COL2A1 expression by adult articular chondrocytes among multiple individual and 2– or 3–growth factor gene combinations of 5 chondrogenic growth factor genes in prior studies using similar experimental conditions. 21 Nevertheless, many other growth factors regulate articular chondrocyte function and the results of the present studies cannot be extrapolated to those other factors.
An additional limitation is the limited information available on transfection efficiency. Since the absolute transfection efficiency cannot be measured for these gene products, the transfection efficiency among vectors was estimated to be similar on the basis of the similar vector design. The relative transfection efficacy among experiments was assessed by measuring IGF-I levels in response to pAAV-IGF-I transfection (Supplementary Table S3). The very small SD in IGF-I values among experiments suggests that the transfection efficiency among experiments was relatively constant. Furthermore, the mean concentration of IGF-I in the conditioned medium of the transfected cells during the first 4 days of culture was similar to the concentration in the medium of cells treated with exogenous IGF-I (Supplementary Table S3).
Chondrocytes transfected with pCMV-COL2A1A or pCMV-ACAN generated a similar number of human COL2A1 and ACAN transcripts from these transgenes (Supplementary Table S2). In contrast to the transgenes, the control chondrocytes produced more transcripts from the native bovine COL2A1 gene than from the native bovine ACAN gene. In addition, the number of bovine COL2A1 transcripts from the native COL2A1 gene in the chondrocytes transfected by empty vector was much larger than the number of human COL2A1 transcripts from the human COL2A1 transgene in pCMV-COL2A1A transfected chondrocytes (4, 19, and 40 times increase at days 2, 4, and 6, respectively). Thus, the human COL2A1 transcripts from the human COL2A1 transgene contributed only a small proportion of the total COL2A1 transcripts in pCMV-COL2A1A transfected cells. Specifically, the human transgene contributed 20%, 5%, and 2% of the total COL2A1 transcripts at days 2, 4, and 6, respectively. In contrast, the number of bovine ACAN transcripts from the native bovine ACAN gene in the control chondrocytes was similar to, or less than, that of human ACAN transcripts from human ACAN transgene in pCMV-ACAN transfected cells. Specifically, the human ACAN transgene contributed 41%, 72%, and 42% of the total ACAN transcripts at days 2, 4, and 6, respectively. These relationships account for the observation that the human aggrecan transgene significantly increased total ACAN transcripts, but the human type II collagen transgene did not significantly increase total COL2A1 transcripts, compared with native bovine gene expression.
Glycosaminoglycans associated with the chondrocytes and their surrounding matrix was used as an index of aggrecan production. Interestingly, the increase in aggrecan transcripts did not lead to an increase in glycosaminoglycans. Although this observation remains unexplained. The ACAN transgene sequence contained a few nucleotide base substitutions, but these did not cause a frame shift or premature termination. It may reflect the use of the human ACAN transgene, rate-limited translation of the transcripts to form the aggrecan core protein, posttranslational modifications, or other reasons. Whatever the underlying mechanism, the data suggest that growth factor stimulation of the native aggrecan gene is superior to adding additional aggrecan transgenes as an approach to the formation of new cartilage matrix. Given the very low expression of the COL2A1 transgene, no measurement of collagen protein was undertaken. The appearance of the chondrocytes generally corresponded to the gene expression or exogenous growth factor levels (Supplementary Figs. S1-S3).
Taken together, these results support the hypothesis that there is a difference between the 2 tested approaches to augmenting matrix molecule gene expression. They suggest that the application of gene therapy to articular cartilage repair may be better achieved by selectively combining genes that encode regulatory molecules than by employing genes that encode the regulated structural molecules.
Footnotes
Acknowledgments and Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported in part by National Institutes of Health, National Institute of Arthritis and Musculoskeletal and Skin Diseases Grant AR047702, US Department of Veterans Affairs, BLR&D Merit Review Award I01 BX000447, and the Indiana University School of Medicine Research Infrastructure Fund. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health. The contents do not represent the views of the U.S. Department of Veterans Affairs or the United States Government.
Declaration of Conflicting Interests
The author(s) declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: Dr. Trippel is a consultant for ELK Orthobiologics.
Ethical Approval
Ethical approval was not sought for the present study because the work does not involve animal or human research.
Trial Registration
Not applicable.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.