
Abstract
Aim
In the osteoarthritis (OA) disease, all structures of the joint are involved. The infrapatellar Hoffa fat pad is rich in macrophages and granulocytes, which also represents a source of adipose mesenchymal progenitor cells (ASC) cells. In our study, we analyze how OA affects the ability of ASC-derived from Hoffa’s fat pad to differentiate into chondrocytes.
Material and methodology
We took knee Hoffa’s pad samples and adipose tissue from the proximal thigh from 6 patients diagnosed with severe OA and from another 6 patients with an anterior cruciate ligament (ACL) rupture without OA. From all the patients, we took subcutaneous adipose tissue from the thigh, as the control group. Samples of synovial fluid (SF) were also extracted. The gene expression was analyzed by real-time quantitative polymerase chain reaction.
Results
PTH1R and MMP13 expression during chondrogenic differentiation were similar between OA and ACL groups, while the expression of OPG, FGF2, TGFβ, MMP3 were significantly lower in the OA group. Exposure of differentiated ASC to OA SF induced an increase in the expression of OPG, PTH1R, and MMP13 and a decrease in the expression of FGF2 in cell culture of the ACL group. However, expression of none of these factors was altered by the OA synovial fluid in ASC cells of the OA group.
Conclusion
OA of the knee also affects the mesenchymal stem cells of Hoffa fat, suggesting that Hoffa fat is a new actor in the OA degenerative process that can contribute to the origin, onset, and progression of the disease.
Keywords
Introduction
Osteoarthritis (OA) leads to progressive destruction of the joint cartilage and also affects the subchondral bone, which is subject to bone remodeling involving osteophyte formation, sclerosis, and subchondral cysts. Moreover, it is also associated with inflammation of the synovial membrane. The human knee contains 1 to 5 mL of synovial fluid (SF), secreted by the synovial membrane, a lubricant that reduces friction, but which is also a vehicle for molecules that protect or destroy the cartilage. 1 The knee joint has also a periarticular adipose tissue and behind the patellar ligament has an area of dense adipose tissue in contact with the anterior cruciate ligament (ACL) and anterior meniscal insertions, which is known as Hoffa’s fat pad. 2 It is covered with fibrous tissue and synovial membrane so that it is not only intraarticular but also extrasynovial, and its function is not well understood. Its nutritional potential has been suggested 3 and has mechanical function in distributing the SF and absorbing loads. It is useful in aiding the vascular supply to meniscal grafts. 4 Hoffa’s fat produces mesenchymal stem cells (MSCs) and enzymes. The most usual source from which MSC are obtained is the bone marrow, the iliac crest being the most efficient area, 5 but MSC derived from adipose tissue, such as Hoffa’s fat pad, are more abundant (5% of nucleate cells in adipose tissue, compared with less than 0.1% in bone marrow) and easier to obtain with less local morbidity, expand more rapidly and more prolifically, and conserve their structure over a larger number of stages. 6
In addition to adipocytes Hoffa’s fat pad, also contains macrophages, lymphocytes, and granulocytes, but like all adipose tissue it is a good source of MSCs. Hoffa’s fat has a dual function in the knee joint 7 : On one hand it reduces the concentration of nitric oxide, the release of glycosaminoglycans, and the expression of matrix metalloproteinase–1 (MMP1) in the cartilage, but it also contains MSC and macrophages. Macrophages of the fatty tissue8,9 differentiate into anti-inflammatory macrophages 10 and pro-inflammatory macrophages, 11 which inhibit chondrogenesis. These macrophages are responsible for producing fibrocartilage instead of hyaline cartilage, 12 a phenomenon that can be observed after a trauma, in cases of microfracture, at the early stages of OA and also in the joints of obese people. Hoffa’s fat expresses greater quantities of inflammatory genes than other adipose tissues, and also releases antichondrogenic factors (interleukin-6 [IL-6], IL 1, tumor necrosis factor-α [TNFα], and adipokines like leptin, resistin, and adiponectin), which are not produced in other subcutaneous tissues. 11 Adipokine concentrations in the SF of OA patients are usually elevated in comparison with healthy controls 13 and the tissues which show a higher concentration of these factors are Hoffa’s fat pad and the synovial membrane.
Our hypothesis is that SF contains elements, not all of which are understood, which act as messengers and alter the “homeostasis” of the knee and the metabolism of all the cellular components of the joint, including the MSC of Hoffa’s fat pad, thus making them another piece in the puzzle as far as OA of the knee is concerned. Hoffa’s fat pad may be a MSCs source to be differentiated to chrondrocytes, which are a therapeutic strategy for chondral and osteochondral knee injuries. For this, the aim of our study is to analyze gene expression of osteoprotegerin (OPG), PTHr1 and transforming growth factor-β (TGF-β), fibroblast growth factor-2 (FGF2), TGF-β and MMP3 and MMP13 factors in chondrocytes derived from MSCs of Hoffa’s fat pad. MMP3 not only degrades the noncollagenous protein matrix but also denatures the collagen itself, while MMP13 directly degrades the intact collagen type II. MMP13 is 10 times more active in the digestion of collagen type II than other collagenases, 14 in young patients with an ACL lesion and in patients with severe knee OA, in which Hoffa’s fat pad and fat from the thigh, far away from the knee joint, as control was extracted.
Materials and Methods
A nonrandomized and nonblinded prospective study using 3 groups was designed. Our study was a level III evidence for individual cross-sectional studies with consistently applied reference standard. Twelve donors were included in this study, with an average age of 25 years (range 20-28 years) in ACL injury patients, an average age of 68 years (range 55-81 years) in osteoarthritis patients. We took Hoffa’s pad samples from 6 patients diagnosed with severe OA, before surgery to insert a total knee replacement (TKR) and samples from another 6 young patients with an ACL rupture without OA. In all the patients, the adipose tissue from the proximal and lateral area of the thigh was also obtained and samples of SF were also extracted ( Fig. 1 ). All donors gave informed consent in accordance with the appropriate clinical protocol. The clinical research ethics committee of our institution approved the study.
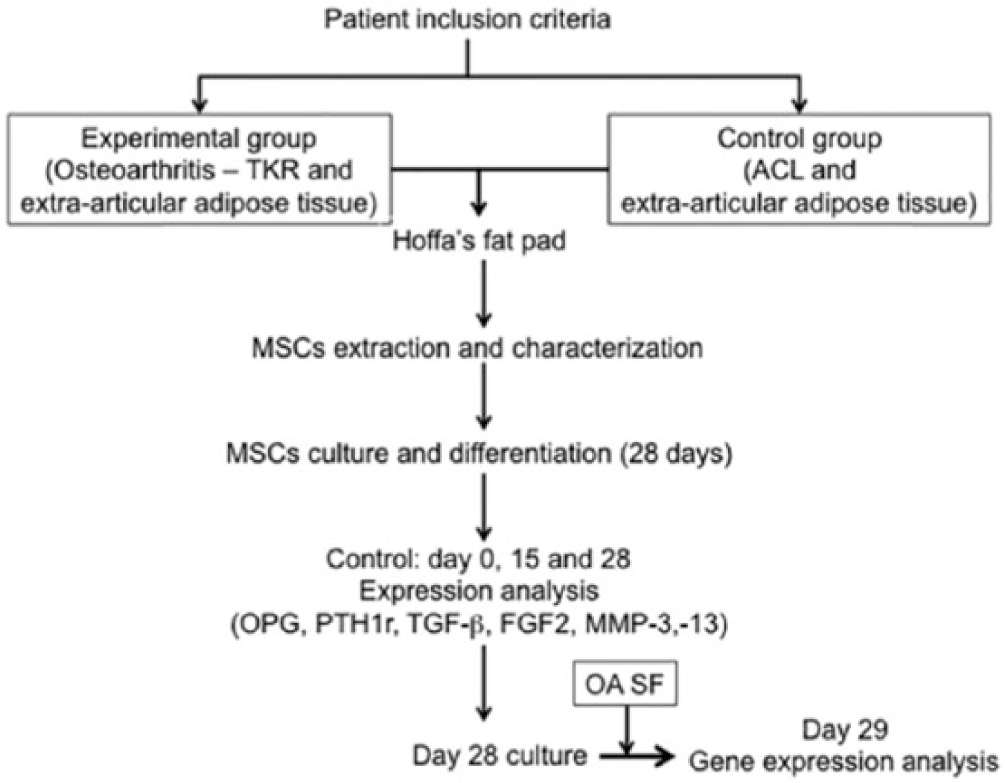
Flow diagram.
The criteria for inclusion in the group of OA patients were that they had received a TKR, with OA of grade 3 or 4 on the Kellgren-Lawrence scale, with a diagnosis of primary OA with no further associated pathologies (such as inflammatory, immunological, or rheumatic conditions). To perform TKR, we used the medial parapatellar approach, taking SF before the operation by puncture with an intramuscular needle and syringe via the medial retinaculum. During the surgery, we extracted Hoffa’s fat pad. In these patients, we took advantage of the incision to extract fat from the proximal anterolateral aspect of the thigh. The control group consisted of patients with no diagnosis of OA, who were being operated on because of injury to the ACL. In none of them was any X-ray sign of joint degeneration detected (grade 0 on the Kellgren-Lawrence scale). The surgery was arthroscopic, and we obtained SF by puncture using an intramuscular needle and syringe via the anteroexternal portal prior to surgery. A portion of Hoffa’s fat pad was extracted through the arthroscopic portal. All the patients were adults, had undergone no operations on the knee in question, were free of inflammatory disease, and took no corticoids or systemic immuno-modulatory treatment, nor did they have toxic habits (smoking, alcoholism, etc.). Their body mass index was less than 35 kg/m2 and they had signed consent authorizing us to extract and process samples from their knees.
The SF used in the culture medium came from 1 patient with severe OA. The SF were put in sterile containers with saline serum, and transported quickly under refrigeration conditions to the laboratory.
Isolation, Primary Culture, and Functional Study of Mesenchymal Stem Cells
Adipose tissue was digested in 0.075% collagenase type I (Invitrogen, Life Technologies, New York, NY) solution for 30 minutes at 37°C, as previously described.
15
Next, cells were plated in plastic substrate (Corning, New York, NY) for 24 hours in DMAX (DMEM (1X)+ Glutamax (Dulbecco’s Modified Eagle Medium (+4.5 g/L
Flow Cytometry
For flow cytometry analysis, MSCs were resuspended in phosphate buffered saline (PBS) at a density of 1 × 106 cells/mL, fixed with 2% paraformaldehyde (PFA) and incubated with fluorochrome (FITC or PE)-conjugated mouse anti-CD90, CD29, CD34, CD45, CD44, CD144, CD31, CD106 and monoclonal antibodies (Biolegend, San Diego, CA) for 30 minutes at room temperature. The cells were washed 3 times with PBS and were analyzed using Accuri’s C6 Flow Cytometer System at 488 nm and 15 mW. Forward scatter (FSC), side scatter (SSC), and specific fluorescent label (LGFL) at 488 nm and 540 nm, were obtained automatically for every cell. Data were digitally collected over a 6-decade dynamic range (16 million channels of digital data). The fluorescence logarithmic amplification and analyses were done using BD Accuri C6 Analysis Software. ASCs from 5 different patients were studied.
Chondrogenic Differentiation Assays
Specific cell lineage differentiation media were used to evaluate the chondrogenic differentiation potential of ASCs. To do this, a 2 to 4 culture passages of MSCs were plated in plastic substrate (Corning, NY) for 24 hours in DMEM and cultured for 2 days. Next, culture medium was exchanged by differentiation media (1% ITS, 0.9 nM sodium piruvate [Gibco Life Technologies], 50 ug/mL ascorbic acid [Sigma], 10−7 M dexamethasone, 40 µg/ml
RNA Isolation and Real-Time Quantitative Polymerase Chain Reaction
The gene expression was analyzed by real-time quantitative polymerase chain reaction (RT-qPCR). We extracted the RNA from all the experimental conditions explained above using Trizol reagent (Sigma-Aldrich; St. Louis, MO). RNA retrotranscription was carried out with 0.5 to 1 µg of total RNA performed with the high-capacity cDNA reverse transcription kit (Applied Biosystems, Foster City, CA). RT-PCR was done with an Assay-by-Design system using a TaqMan MGB probe (Applied Biosystems, Foster City, CA) for OPG, PTH1r, TGFβ, MMP3, MMP13 and FGF2 Premix Ex-Taq (Takara, Otsu, Japan) in an Applied 7900HT Fast Real Time PCR System (Applied Biosystem) Results were calculated for each sample using the cycle threshold (Ct) value, and normalized against 18S.
Statistical Analysis
Results were expressed as mean ± SD. Differences between experimental and control conditions were analyzed by using the 2-tailed t test or Mann-Whitney test when appropriate, and P < 0.05 was considered significant. Descriptive statistic was performed and to compare quantitative variables with factors or groups, Student’s t test for independent samples and analysis of variance (ANOVA) was applied. In all cases, the homoscedasticity will be evaluated with Levene test and normality with the Kolmogorov-Smirnov test, along with the graphs to evaluate normality conditions. A Mann Whitney nonparametric ANOVA test robust Brown-Forsythe and Welch is used when normality or homoscedasticity are refused.
Results
Hoffa fat samples were characterized for their morphology. The MSCs were plated for 24 hours and adherent cells were primary cultured for 7 days until confluent stage ( Fig. 2 ). They were next passaged and cultured for another 7 days. Phenotypic analysis of MSCs by flow cytometry revealed that 99.33% were CD90+/CD34– and 97,45% CD29+/CD45– ( Fig. 3 ). In addition, 91.6% were CD44+, 92.4% CD106–, 99.8% CD31–, 99.4% CD144–, and 99.5% KDR– (data not shown). Specific cell lineage differentiation media were next used to evaluate the adipogenic and osteogenic differentiation potential of MSCs ( Fig. 2 ). Oil Red histochemical staining revealed the presence of lipid vacuoles on the 14th day in the majority (88%) of primary cultured MSCs. Alizarin Red histochemical staining also revealed that the number of MSCs per microscopic field surrounded by red-stained extracellular matrix was 14 ± 7 cells, representing 87% of ASCs given osteogenic differentiation medium for 21 days ( Fig. 2 ).
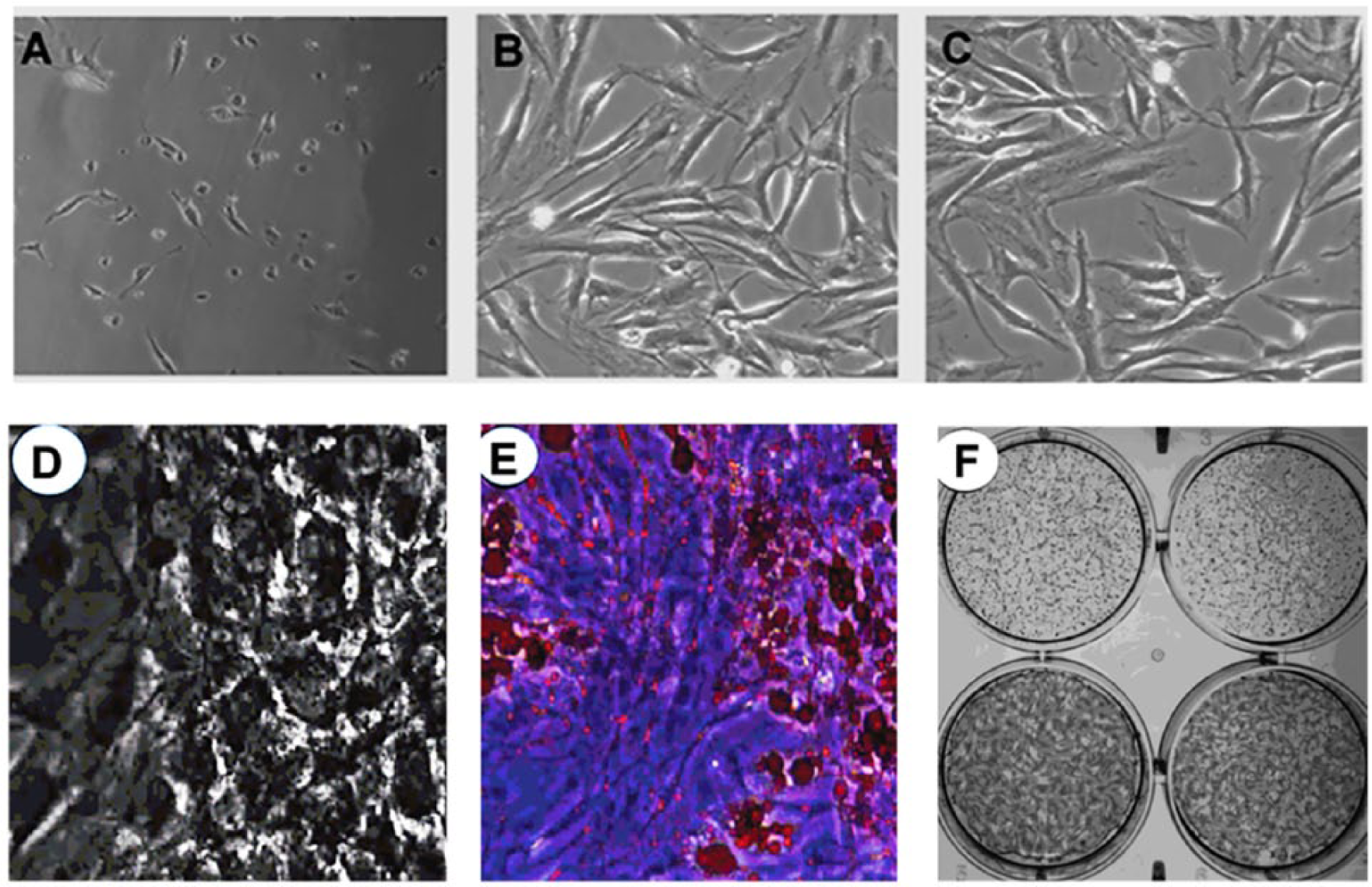
Contrast-phase microscopic image from fleshly isolated and primary cultured adipose stem cells (ASCs) on day 1 (
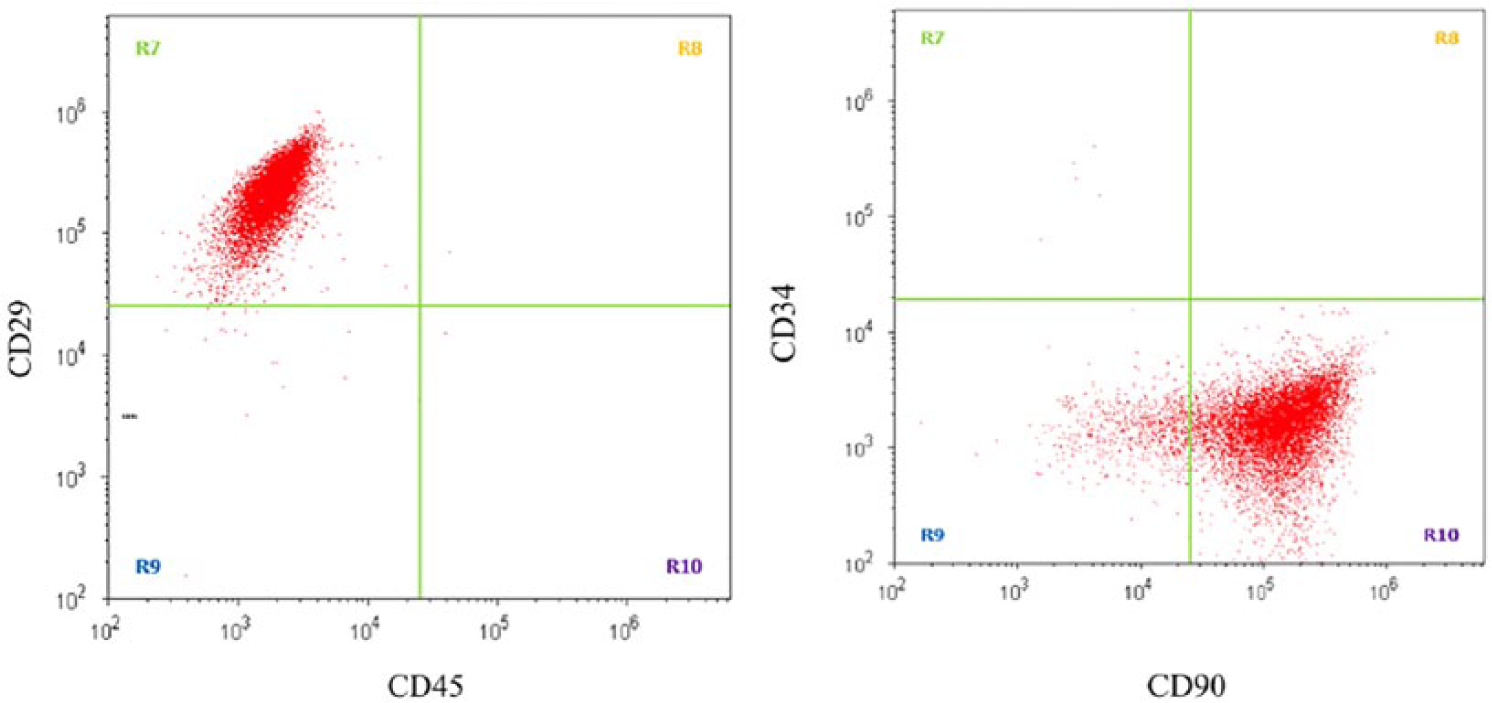
Flow cytometric analysis at day 7 on the surface expression of CD29-CD45 (
The genetic analysis demonstrated a significant (P < 0.05) increase of levels in the OPG, PTH1R, FGF2, MMP3, and MMP13 expression in ACL group at 0, 15, and 28 days for chondrocyte differentiation ( Fig. 4 ). However, the expression levels in the OA group not showed a significant increase in OPG, PTH1R, FGF2, TGFβ, and MMP3 ( Fig. 4 ).
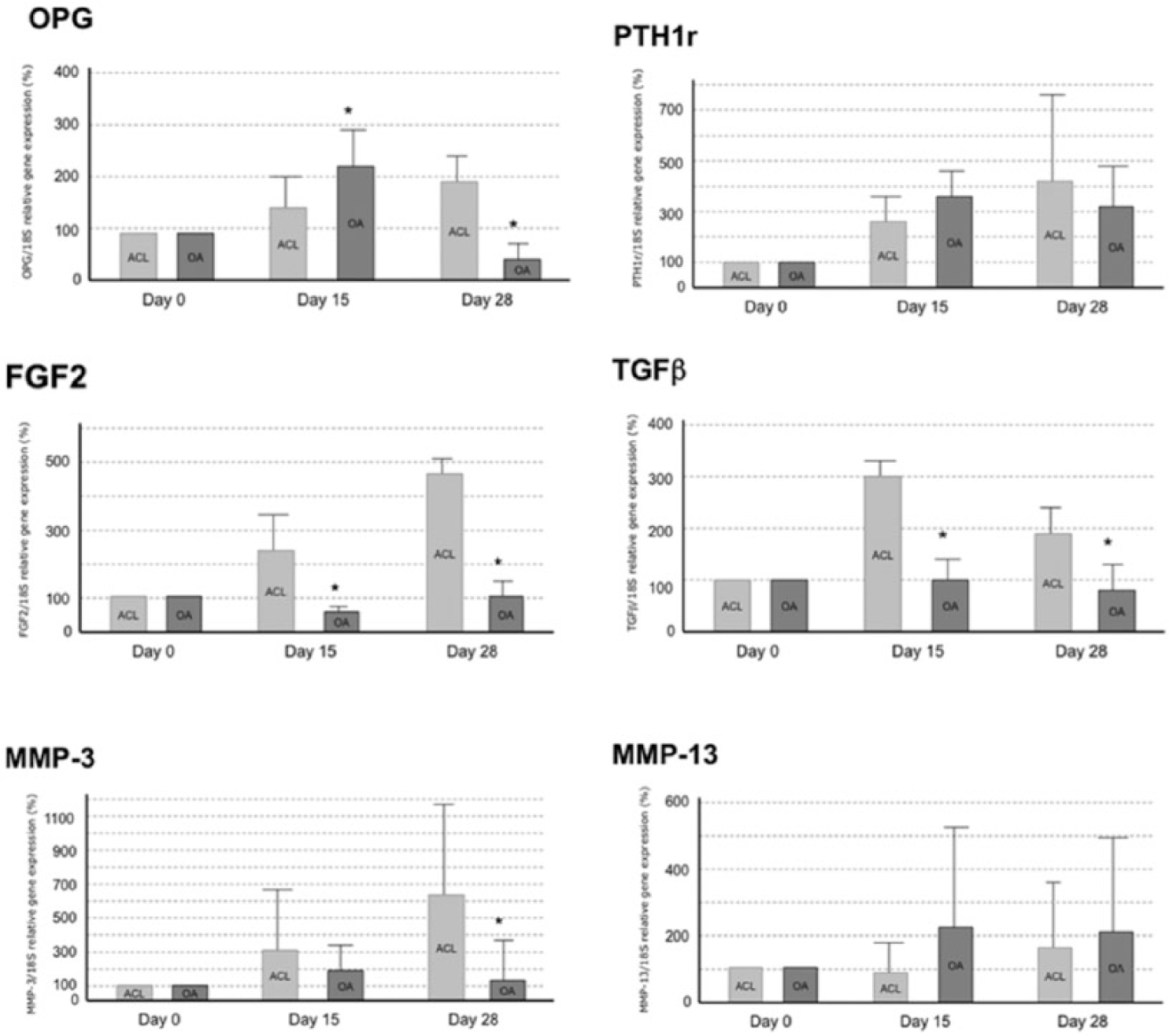
Gene expression of OPG (
The genetic analysis also demonstrated a significant (P < 0.05) increase in the OPG, PTH1R, TGFβ, and MMP13 expression in ACL group after SF treatment. SF treatment did not induce any change in these gene expressions in the OA group. Moreover, in the control fat group (subcutaneous fat) SF abrogated the expression of OPG, PTH1R, FGF2, TGFβ, and MMP13 expression but increased MMP3 expression ( Fig. 5 ).
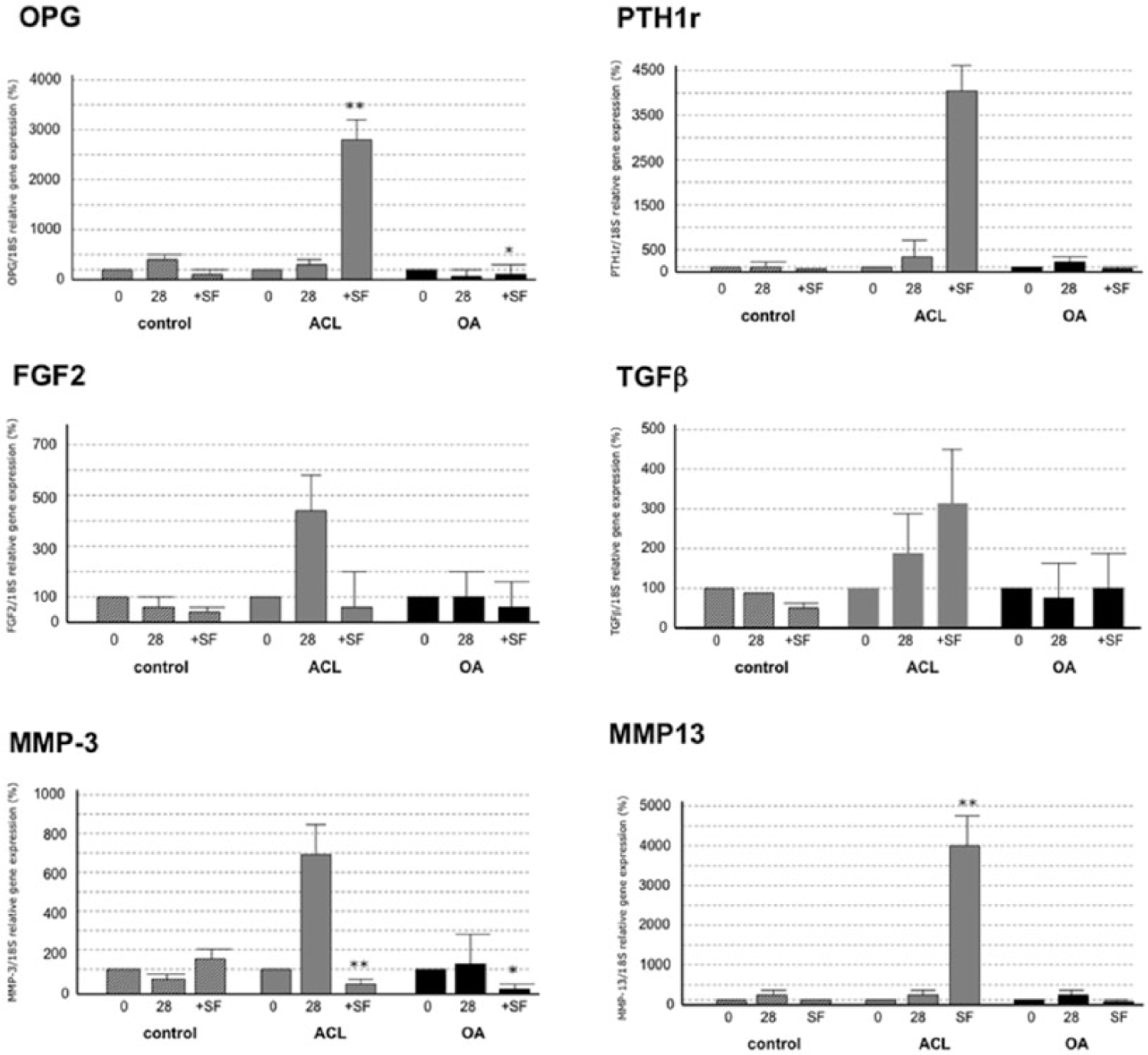
Gene expression of OPG (
Discussion
In articular trauma and OA, the joint release of anti-inflammatory cytokines is intensified.11,16,17 Moreover, the SF receives large numbers of cytokines (bFGF, VEGF, TNFα), IL6, growth factors and adipokines, which control MMP expression and pro-inflammatory cytokines, which degrade the extracellular chondral and synovial matrix. 18
It is still unclear whether Hoffa’s fat triggers the degenerative process in the synovial membrane, or whether it is the synovial membrane, which affects Hoffa’s fat, or whether the 2 processes occur simultaneously. Eymard et al. 19 analyzed specimens of Hoffa’s fat, subcutaneous cellular adipose tissue, and synovial membrane from OA patients and observed little response to extracellular adipose tissue, whereas Hoffa’s fat overexpressed degenerative factors (IL6 and IL8, PGE2, COX-2, MMP1, -3, -9, -13), which led them to suggest that Hoffa’s fat induces the synovial membrane into the process of degeneration. Their results are consistent with our study, in which we found an increase or decrease in the concentration of the different biomarkers, but no modifications of the fat obtained outside the joint. Furthermore, Hoffa’s tissue has greater inflammatory activity in ruptures of the ACL. 20 Cuellar et al. 21 studied the profile of pro-inflammatory cytokines in the SF of patients with acute knee pain secondary to meniscal injury, and observed that the expression of pro-inflammatory cytokines was greater in the symptomatic than the asymptomatic knees. Higuchi et al. 22 also showed that the concentration of IL6 remained high in the SF of knees with injury to the ACL 50 weeks after the lesion occurred. Martinez de Albornoz and Forriol 1 analyzed SF from knees with meniscus tears, ACL, and isolated chondral injury, and noted that the SF environment in the ACL injury was mainly anabolic and inflammatory, with increased levels of IL1, IL6, significant levels of TGFβ, IL10, and significantly decreased levels of TNFα. There was mainly a catabolic environment in chondral and meniscal disease, with a significant increase in TNFα and a significant decrease in TGFβ.
In the present study, we measured the expression of anabolic factors from bone (OPG, PTH1r, TGFβ) and cartilage (TGFβ and FGF2), and 2 catabolic factors (MMP3 and MMP13). FGF2 acts as a protective agent against aggressions 23 and is overexpressed in SF in animal knees subjected to mechanical overload or with chondral injuries. 24 For its part, TGFβ not only shares anabolic functions but also plays a part in apoptosis 25 and helps repair chondral and meniscal injuries.26,27 In OA cartilage, it is overexpressed in the most superficial cells. Ye et al. 28 observed that MSC from Hoffa’s fat cultured in the presence of TGFβ and BMP6 differentiated to chondrocytes, taking on a 3-dimensional shape.
MMP3 degrades collagen of types II, III, IV, IX, and X, as well as proteoglycans, fibronectin, laminin, and elastin. It can also activate other MMPs like -1, -7, and -9, to remodel the conjunctive tissue. 28 In the development of OA, MMP3 and TNFα are responsible for the traditional association between hyperlipidemia, obesity, and OA. 15 On the other hand, MMP13 is overexpressed in pathological situations such as carcinomas, rheumatoid arthritis, and OA.29,30 The 2 MMPs analyzed behaved differently. The concentration of MMP3 did not increase in patients from the control group. However, in the study group it rose gradually over time, and also when SF was added. It is possible that the presence of TIMPs in the control group was responsible for this behavior, while its absence in the study group failed to inhibit MMP3. In the case of MMP13, what happens is similar to the situation with osteogenic factors: a gradual increase in the control group, which soars with the addition of SF. In the study group, the concentration of MMP-13 diminished when SF was added.
MMP3, -10, and -11 not only degrade the noncollagenous protein matrix, but also denature the collagen itself, while MMP1 and -13 directly degrade the intact collagen type II. MMP13 is 10 times more active in the digestion of collagen type II than other collagenases. 14 Haller et al. 14 noted how, in the SF from joints that had suffered a trauma, the MMP3 level was raised, as was that of MMP1, -9, -10, and -12, while the same was not the case with MMP13, as this MMP initially allows other inflammatory cytokines and MMPs to act, thus affecting chronic processes.31,32
Our study is obviously limited in terms of the small number of samples used. Moreover, we were unable to analyze the action of the non-OA fluid, as its volume in healthy patients is very small and extraction proves difficult. The 2 groups in this study diverged widely in terms of age and sex. We must also point to a common limitation of all clinical studies on OA, which is that the diagnosis is performed by radiographic imaging, since this is the only validated noninvasive method, and the assessment scales lack sufficient sensitivity and specificity to characterize differences in order for homogeneous study groups to be reliably established. Finally, the study was not blinded.
Our results show that OA of the knee, in its more severe phases, also affects the MSCs of Hoffa’s fat, which is a new actor in the OA degenerative process and which can contribute to the origin, onset, and progression of the disease. Chondrocytes derived from MSC in Hoffa’s fat express anabolic factors related to cartilage (FGF2 and TGFβ) and bone (PTHr1 and OPG) as well as catabolic factors (MMP3 and MMP13) differently in the 2 groups. In all cases, the control group showed a tendency for expression to be higher on day 28 of culture, while we found no variations in the cultures of OA chondrocytes. Only MMP3 followed the opposite pattern, showing no variation in the control group, while its expression had increased on day 28 in the culture of OA chondrocytes.
Footnotes
Acknowledgments and Funding
The authors thank Irene Buendía and Sonia Moraleja for technical support. The author(s) received no financial support for the research, authorship, and/or publication of this article.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical Approval
Ethical approval for this study was obtained from the clinical research ethics committee of our institution (Comité Ético de Investigación Clínica – Area 1, Madrid, Spain, CI: 317/11).
Informed Consent
All donors gave informed consent in accordance with the appropriate clinical protocol.