
Abstract
Background
Mandibular condyle cartilage (MCC) has a unique structure among articular cartilages; however, little is known about its nanoscale collagen network architecture, hampering design of regeneration therapies and rigorous evaluation of regeneration experiment outcomes in preclinical research. Helium ion microscopy is a novel technology with a long depth of field that is uniquely suited to imaging open 3D collagen networks at multiple scales without obscuring conductive coatings.
Objective
The objective of this research was to image, at the micro- and nanoscales, the depth-dependent MCC collagen network architecture.
Design
MCC was collected from New Zealand white rabbits. Images of MCC zones were acquired using helium ion, transmission electron, and light microscopy. Network fibril and canal diameters were measured.
Results
For the first time, the MCC was visualized as a 3D collagen fibril structure at the nanoscale, the length scale of network assembly. Fibril diameters ranged from 7 to 110 nm and varied by zone. The articular surface was composed of a fine mesh that was woven through thin layers of larger fibrils. The fibrous zone was composed of approximately orthogonal lamellae of aligned fibrils. Fibrocyte processes surrounded collagen bundles forming extracellular compartments. The proliferative, mature, and hypertrophic zones were composed of a branched network that was progressively remodeled to accommodate chondrocyte hypertrophy. Osteoid fibrils were woven around osteoblast cytoplasmic processes to create numerous canals similar in size to canaliculi of mature bone.
Conclusion
This multiscale investigation advances our foundational understanding of the complex, layered 3D architecture of the MCC collagen network.
Keywords
Introduction
The temporomandibular joint (TMJ) is the most active joint in the body, 1 completing as many as 2,000 cycles per day 2 ; however, characterization of the structure of TMJ tissues has received little attention relative to that of other joints. The TMJ is unique because its articular cartilage is classified as a fibrocartilage, 3 not a hyaline cartilage like that of other articulating joints. It is also a secondary cartilage that arises from the periosteum, not from a mesenchymal condensation as hyaline cartilage does.4,5 Degeneration of the TMJ articular cartilage results in pain that interferes with eating, speaking, and yawning. 6 TMJ disorders are the second most common musculoskeletal condition after chronic low back pain with an estimated annual treatment cost of $4 billion. 7 They may be precipitated by trauma to the head or neck, 8 functional or parafunctional overload or articular disc derangement, 9 or hormonal influences. 10
There has been an increasing emphasis on regenerating damaged mandibular condyle cartilage (MCC).11-13 The goal of cartilage regeneration therapies is to produce a durable tissue for long-term repair. However, there is little foundational knowledge on the TMJ articular cartilage collagen network nanostructure, the very structure researchers aim to regenerate. The structure of articular cartilages, particularly that of the collagen network, is highly anisotropic, and it determines articular cartilage mechanical function.14-17 Designing a regeneration therapy and evaluating the outcome of preclinical trials requires a detailed understanding of the structure and mechanical function of the native tissue. The collagen network is a fundamental structural component of articular cartilage, and failure to mimic the structural features of the native network or to develop robust integration of the regenerated and native networks will result in degeneration of the repair. Without the knowledge base and tools necessary to evaluate network nanostructure, researchers are limited to lengthy animal studies to evaluate the durability of repair tissue produced by candidate regeneration therapies, increasing research cost and slowing the rate of progress. This critical gap also limits our insight into TMJ articular cartilage function in health and the structural changes that occur during development and due to injury or disease.
Imaging a collagen network is technically challenging because it is an open 3D structure with features of interest at the nanoscale. Collagen is also a nonconducting material resulting in sample charging and image artifacts during high-resolution scanning electron microscopy (SEM). Conductive coatings applied to enable high resolution SEM obscure nanoscale network features. Helium ion microscopy (HIM), a recently introduced technology, uses a positively charged helium ion beam that enables sample charge neutralization for nanoscale imaging of uncoated fibril surfaces and a 5× longer depth of field to visualize open 3D structures.18,19 The objective of this research was to visualize, at the microscale and nanoscale, the depth-dependent collagen network architecture of the MCC in the rabbit, an established preclinical model.11,20,21 Cartilage at the late juvenile stage was imaged, a point at which the stratified surface layers have developed and the subchondral plate will soon be forming. HIM, in conjunction with transmission electron (TEM) and light microscopy, was used to evaluate the MCC collagen network structure.
This multiscale visualization revealed that the fibrocartilage layer of the MCC has an orthogonal lamellar structure with fibrocyte morphology typical of that found in tendons and the cornea. The collagen network through the proliferative, mature, and hypertrophic layers was extensively remodeled to accommodate chondrocyte growth. In the developing subchondral bone, the osteoid collagen network deposited on the calcified cartilage scaffold was woven about numerous osteoblast cell processes, creating a layer punctuated with canals. This study reports for the first time the nanostructural features of healthy TMJ MCC without obscuring conductive coatings, which is essential knowledge for the evaluation of candidate regeneration therapies and the basic understanding of the development and function of the MCC in health and disease.
Methods
Cartilage Tissue
Left and right mandibular condyles were collected from 5 female New Zealand white rabbits at the late juvenile stage, age approximately 3.5 months. The rabbits were humanely euthanized under a protocol approved by the Medical University of South Carolina Institutional Care and Use Committee.
Helium Ion Microscopy
Tissue Preparation
The collagen network architecture of MCC was revealed by enzymatic digestion to remove both proteoglycans and cells. Mandibular condyles were cut into approximately 2 mm slices in the coronal plane (i.e., medial-to-lateral) ( Fig. 1A ). A previously reported enzymatic digestion protocol 18 was used with the addition of a broad spectrum protease inhibitor cocktail (10 mM ethylenediaminetetraacetic acid [EDTA], 1% [v/v] protease inhibitor cocktail [P8340, Sigma, St. Louis, MO]) and 0.02% (w/v) sodium azide. Digests were performed at 37°C with gentle agitation. Chondroitin and dermatan sulfate were removed with 0.6 U mL−1 chondroitinase ABC (Sigma-Aldrich, St. Louis, MO) in 0.05 m Tris–HCL and 0.06 M sodium acetate at pH 8.0 for 24 hours. Hyaluronic acid and keratan sulfate were removed with 20.4 U mL−1 hyaluronidase (Sigma) and 1.0 U mL−1 keratanase II (Seikagaku, Tokyo, Japan), respectively, in phosphate-buffered saline (PBS) with 10 mM sodium acetate at pH 6.0 for 24 hours. DNA was removed with 1,082 Kunitz U mL−1 DNase I (Worthington Biodhemical Corp., Lakewood, NJ) in PBS with 1 mM CaCl2 and 6 mM MgCl2 at pH 7.7 for 1 hour. The DNase buffer did not contain EDTA. The samples were rinsed in PBS for 12 hours, in deionized water, without protease inhibitors or sodium azide, for 3 hours, dehydrated in a graded ethanol series (50% to 100%), and dried at critical point (EMS 850, Electron Microscopy Sciences, Hatfield, PA). Note that samples prepared for HIM imaging were not aldehyde-fixed, and no conductive coatings were applied. Aldehyde fixation chemically crosslinks proteins to collagen fibrils, which obscures network morphology.
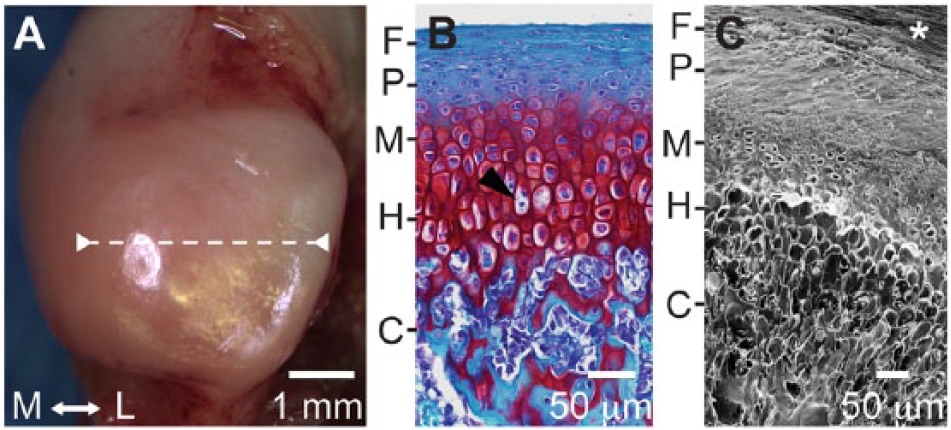
Gross and microscale morphology of rabbit mandibular condyle cartilage. (
Image Acquisition
Micro- and nanoscale features of the MCC collagen network were imaged using a HIM (Orion Plus, Carl Zeiss Microscopy, LLC, Peabody, MA). Images were acquired in secondary electron mode with an acceleration voltage of 34.9 kV, a beam current of 0.2 to 0.5 pA, and a dwell time of 0.1 to 0.9 µs. Signal was acquired with an in-chamber Everhart-Thornley detector. Sample charge was neutralized with an electron flood gun. Images were acquired from each zone to visualize morphological changes in the collagen network architecture. High-resolution images, with a 1 µm field of view and 0.49 nm/pixel, were acquired to visualize fibril morphologies and connectivity. Image brightness and contrast were adjusted.
Network Feature Measurement
Collagen fibril diameters were measured from a representative image for each unmineralized zone. The images were selected from a single condyle in which fibril diameters were clearly visible in all zones. The images were aligned along a path perpendicular to the articulating surface. A 12 × 12 sampling grid was overlaid on each image (ImageJ v1.44p).22,23 Diameter was measured at each grid point with a clearly visible fibril. A minimum of 100 fibril diameters was measured for each zone.
The size and spacing of osteoid canal openings were measured in a 10 µm field of view ( Fig. 2 ). An elliptical template was deformed to mask each opening (Adobe Illustrator vCS5, Adobe Systems, Inc., San Jose, CA). Feret’s minimum and maximum (caliper) distances were measured from the binary mask image (ImageJ). Spacing was characterized using nearest neighbor distances. Neighbors were identified using the ImageJ nearest neighbor macro, 24 which is based on a Voroni tessellation of the mask image ( Fig. 2 ). Tessellation divided the image into polygonal cells, each containing a single opening. The neighbors of a selected opening were those whose polygons shared an edge with the corresponding selected polygon. The set of nearest neighbors was identified for each opening, and the centroid-to-centroid distance between an opening and its neighbors was measured.
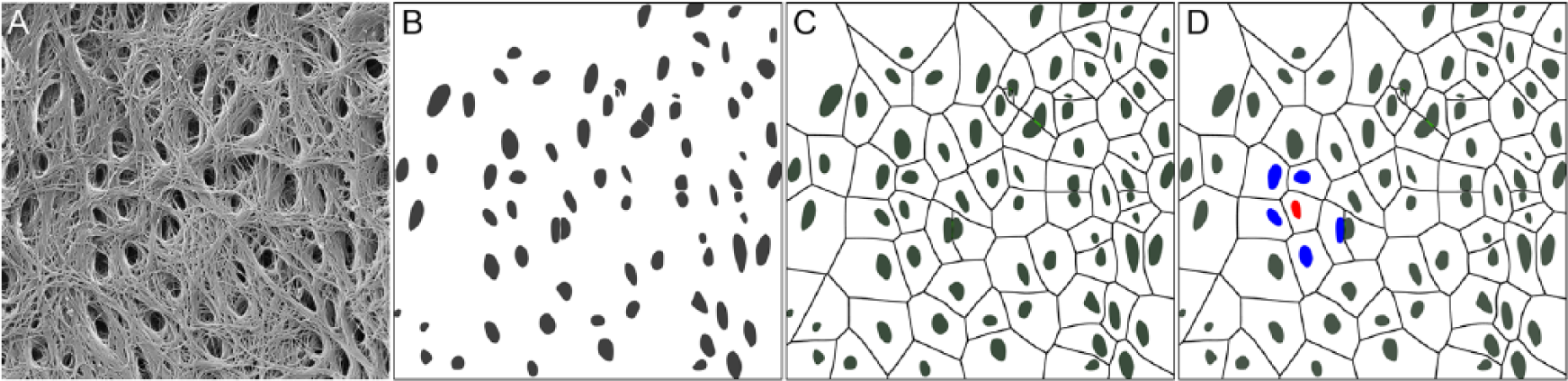
Osteoid canal nearest neighbor identification. (
Transmission Electron Microscopy
Mandibular condyles were fixed in 2.5% (v/v) glutaraldehyde and 2% (w/v) paraformaldehyde in 0.1 M sodium phosphate buffer for 24 hours. The articular cartilage, with an attached layer of subchondral bone, was cut from the condyle with a disposable microtome blade. A new blade was used for each sample to avoid tearing and crushing artifacts. The cartilage was then postfixed in OsO4 for 1 hour and embedded in Epon 812 resin. Ultrathin sections, 70 nm, were stained with saturated uranyl acetate and Reynold’s lead citrate for 10 minutes each. The solutions prior to dehydration included 0.6% (w/v) ruthenium hexaammine trichloride (Strem Chemicals, Newburyport, MA) to preserve cell morphology. 25 The fibrous zone and osteoid were imaged with an acceleration voltage of 80.0 kV (JEM-1210, JEOL, Peabody, MA) and an AMT digital camera (Advanced Microscopy Techniques Corp., Danvers, MA). Image brightness and contrast were adjusted.
Cartilage Histology
Mandibular condyles were fixed in 10% neutral buffered formalin for 48 hours. Selected condyles were decalcified in formic acid (Immunocal, Decal Chemical Corp., Tallman, NY) and paraffin embedded. Sulfated glycosaminoglycans were demonstrated by safranin O with a fast green counter stain. The non-decalcified condyles were embedded in methylmethacrylate. Mineralization was demonstrated by McNeal’s tetrachrome with a von Kossa counter stain. Osteoid was identified by Movat’s pentachrome stain. Images were acquired with a light microscope (Olympus America, Inc., Center Valley, PA) equipped with a digital color camera, motorized stage, and VisioPharm image acquisition software (v4.6, Hoersholm, Denmark).
Statistical Analyses
Collagen fibril diameters among structural zones were evaluated by Welch’s ANOVA (v9.3, SAS Institute, Inc., Cary, NC). Residual normality was evaluated graphically using quantile-quantile plots for the normal distribution. The assumption of normality was satisfied by a Box-Cox power transformation with a λ of -0.2. Homogeneity of variance was evaluated with Levene’s test. The Games-Howell test was used for multiple post hoc comparisons to control family-wise error. Significance was set at P < 0.05.
Results
The rabbit mandibular condyles were free of injury and signs of degeneration ( Fig. 1A and B ). The safranin O fast green stain revealed the structural zones characteristic of MCC ( Fig. 1B ). Glycosaminoglycan staining was absent in the fibrous zone, abundant in the mature and hypertrophic zones, and it persisted in the calcified cartilage scaffolds. The enzymatic digestion and critical point drying protocol produced samples without fibril buckling or breakage. The appearance of the HIM images was strikingly different than that of the optical histology images ( Fig. 1B and C ). The long depth of field generated 3D-like images of the extracellular matrix with empty cell lacunae.
The collagen fibril diameter distributions from the unmineralized cartilage zones and the osteoid were positively skewed, with a larger proportion of smaller diameter fibrils. Mean fibril diameter varied among the zones (F4,245.7 = 110.29, P < 0.001) ranging from 14.63 nm in the mature to 32.58 nm in the fibrous zone ( Table 1 ). Fibrils from the surface and fibrous zones were both significantly larger than those from the proliferative and mature zones (P < 0.001). Fibrils in the newly deposited osteoid layer were significantly smaller than those in the fibrous zone (P < 0.001) and significantly larger than those in the mature and proliferative zones (P < 0.001).
Collagen Fibril Diameters: P Values for Least-Squared Means Comparisons Among Structural Zones a .
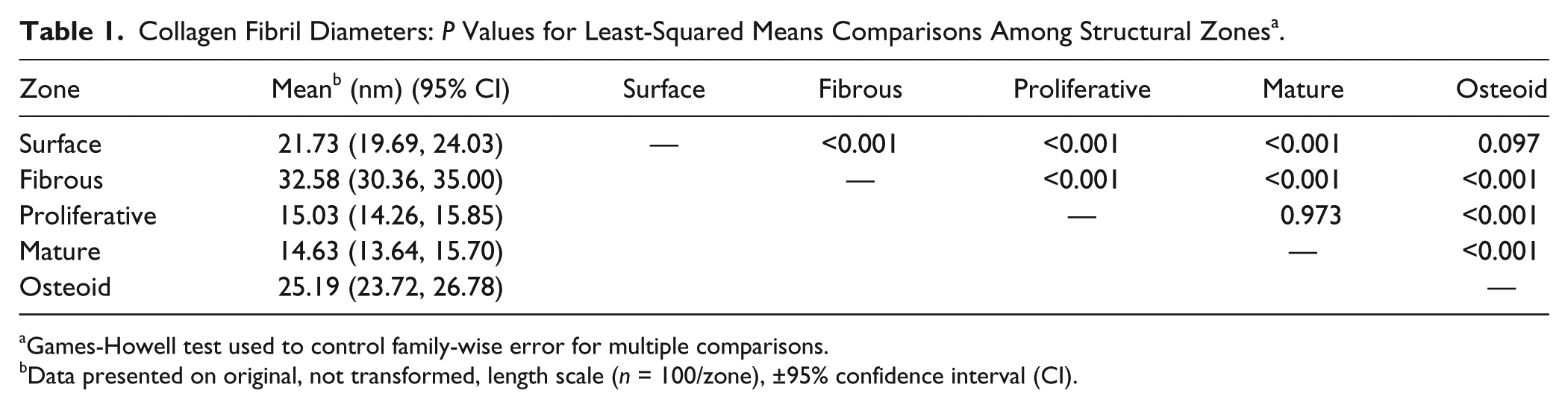
Games-Howell test used to control family-wise error for multiple comparisons.
Data presented on original, not transformed, length scale (n = 100/zone), ±95% confidence interval (CI).
The articular surface was covered by a fine network of fibrils, as small as 7 nm in diameter that overlaid and were interspersed within a network of fibrils as large as 110 nm in diameter ( Fig. 3A ). The larger fibrils were approximately aligned with the anterior-posterior direction, the primary direction of motion. The underlying layer of fibrils was approximately orthogonal to the top layer and was aligned in the medial-lateral direction. The outer 1 μm at the cartilage surface formed a distinct surface zone that was composed of alternating orthogonal layers, each a few collagen fibrils thick ( Fig. 3C2 ).
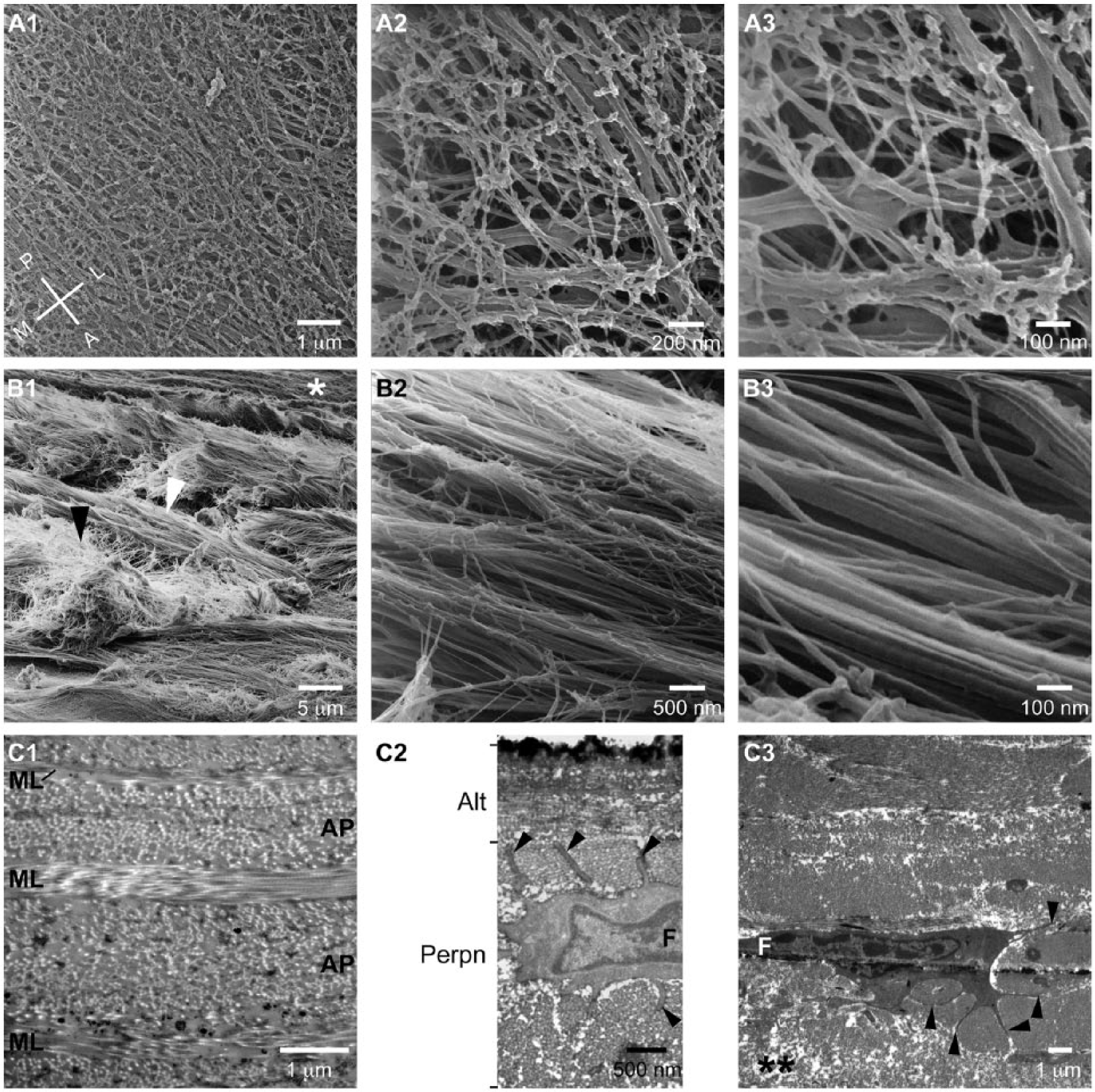
Mandibular cartilage articular surface and fibrous zone. (
The fibrous zone was composed of dense bundles of fibrils arranged in approximately orthogonal layers ( Fig. 3B1 and C1 ). The anterior-posterior layers were thicker than the medial-lateral layers ( Fig. 3C1 ). Individual bundles were composed of closely packed collagen fibrils arranged in parallel ( Fig. 3B2 and B3 ). The cells had a fibroblast morphology with thin cytoplasmic processes that encircled adjacent collagen bundles ( Fig. 3C2 and C3 ).
The proliferative zone network was an open multidirectional branched structure ( Fig. 4 ). The transition between the aligned bundles of the fibrous zone and the branched network of the proliferative zone was abrupt ( Figs. 3C3 and 4A ), and it was a frequent location of tearing ( Fig. 5D1 ). The cells were rounded ( Fig. 1B ) in contrast to the elongated fibroblasts in the fibrous zone.
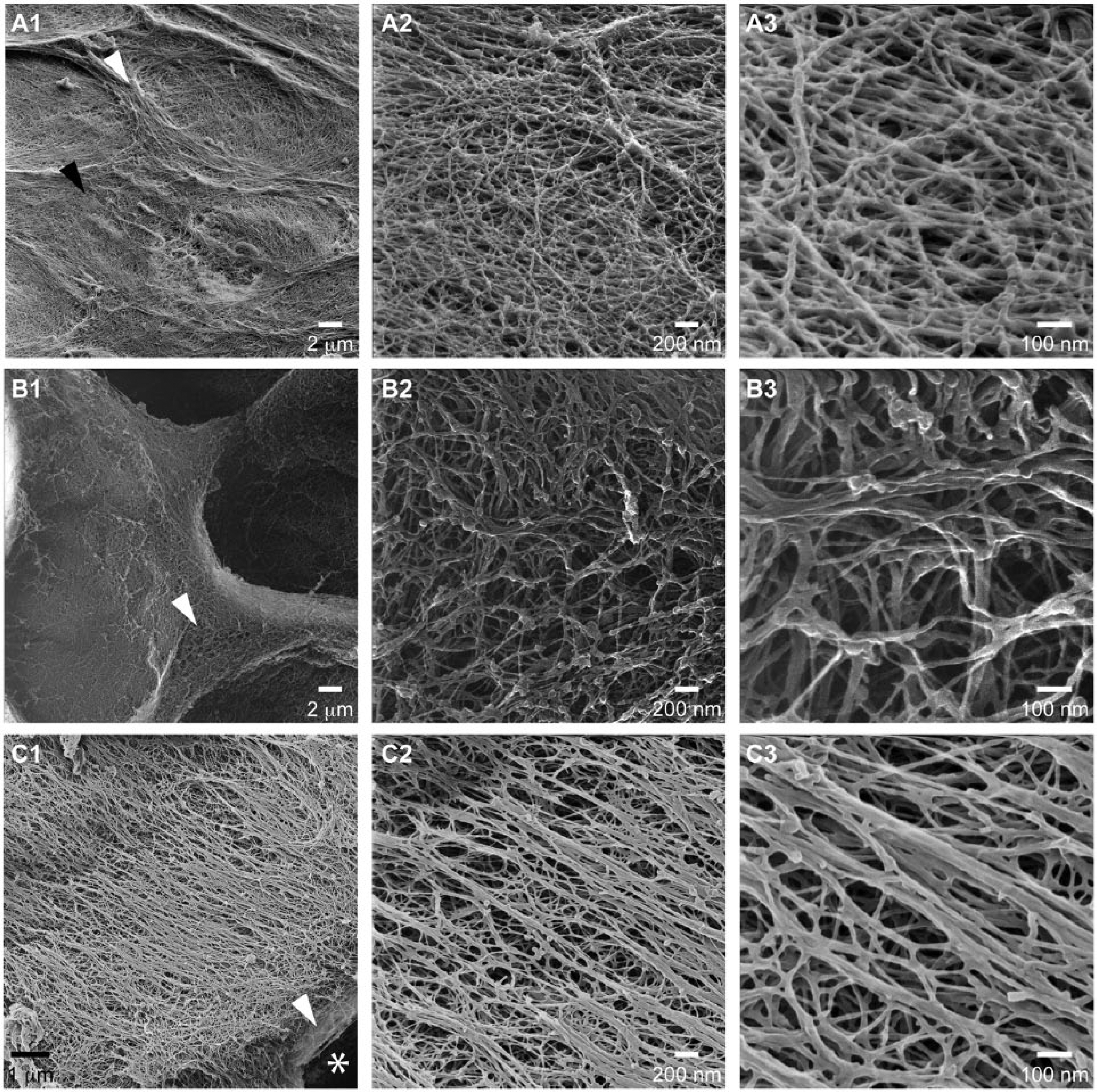
Mandibular cartilage proliferative and mature zones. (
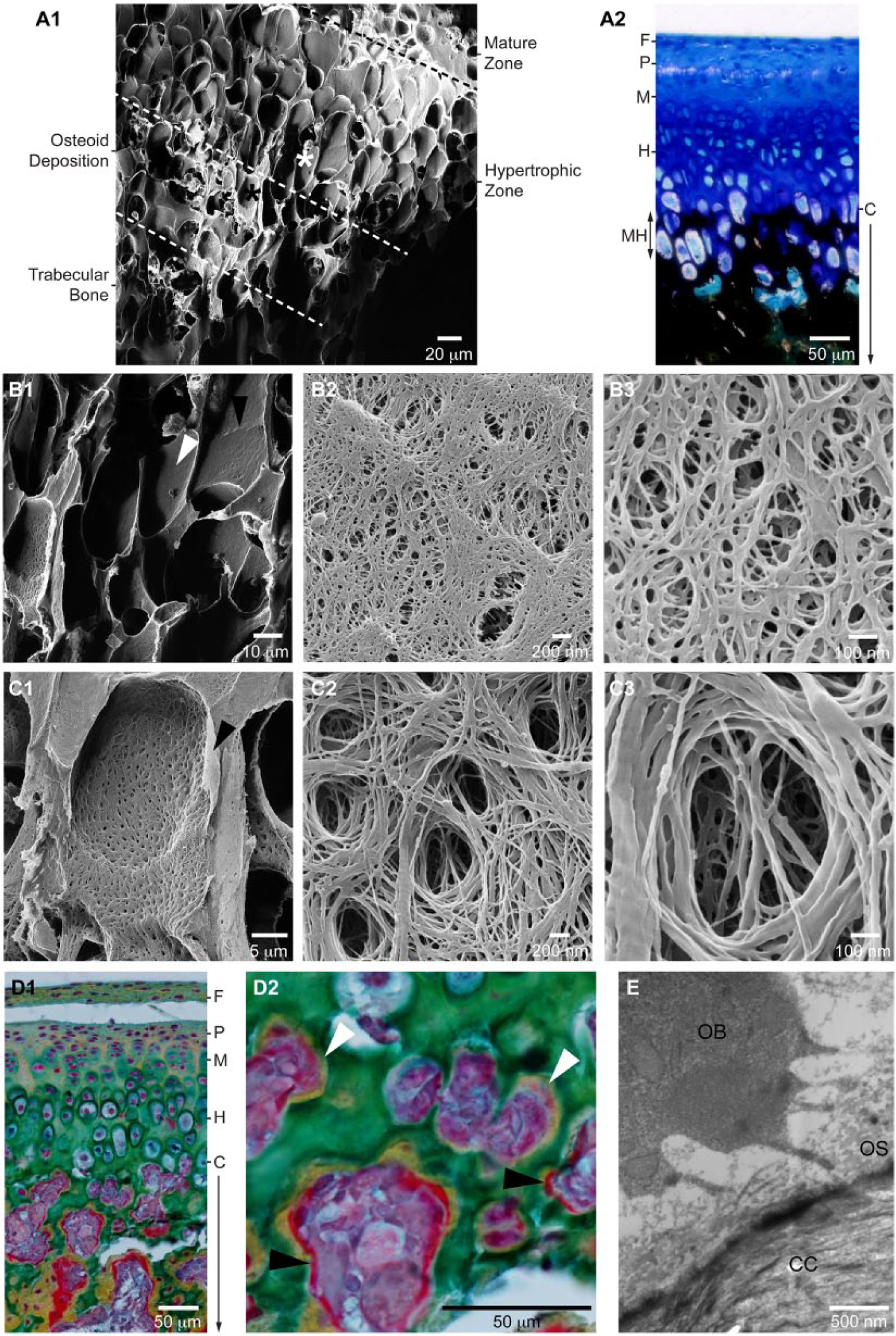
Mandibular cartilage hypertrophic zone and osteoid deposition. (
In the mature zone, the loose network of the proliferative zone was compressed into septae in the intraterritorial space of cell clusters ( Fig. 4 ). The fibril network in the interterritorial space was branched, but displayed a dominant directional alignment ( Fig. 4C ).
The hypertrophic zone was characterized by a dramatic growth in chondrocyte size in the direction of condyle growth ( Fig. 1B ). Remodeling of the collagen network resulted in a high void volume and thin compressed septae between the cell lacunae, mean thickness 7.48 μm (SD 2.58, n = 25) ( Fig. 5A1 and B1 ). Positive von Kossa staining demonstrated that the matrix of the lower hypertrophic zone was calcified ( Fig. 5A2 ). There was evidence that septae between some cell lacunae had been eroded ( Figs. 5B1 and 2B ). The collagen fibrils in the calcified lacunae had a flattened and stacked appearance rather than the round cross section typical of fibrils in the non-mineralized zones ( Fig. 5B2 , B3 ).
Trabecular bone formation was initiated by osteoid deposition on the calcified cartilage scaffold ( Figs. 1B and 5D1 ). The osteoid was a punctate layer composed of collagen fibrils woven to create numerous canals ( Fig. 5C ) that contained osteoblast cytoplasmic processes ( Fig. 5E ). In Movat’s pentachrome stain, the osteoid layer progressed from a yellow stain, indicating collagen, to scarlet red as collagen and noncollagenous protein deposition advanced ( Fig. 5D2 ). The median nearest neighbor distance among the canal openings was 1.21 μm (min 0.21, max 2.88, n = 75). The median of the maximum Feret (caliper) diameter was 0.52 μm (min 0.21, max 1.01); the median of the minimum Feret diameter was 0.29 μm (min 0.10, max 0.44).
Discussion
For the first time, the complex, layered 3D architecture of the MCC collagen network was revealed from the microscale tissue level to the nanoscale collagen fibril level. The uncoated fibril network was clearly visualized by HIM as a 3D assembly of interconnected collagen fibrils, with striking differences in fibril morphologies and diameter distributions among the zones. The micro- and nanoscale features revealed by HIM were identified in standard histological and TEM images to link this new data to the existing knowledge base.
Previous investigations of collagen network architecture, including that of rat, 26 macaque, 27 and human 28 MCC have used SEM to image gold- and platinum-sputtered samples. The data presented here extend that knowledge base. With HIM, the 5× longer depth of field enabled visualization deep into the collagen network revealing 3D structural information ranging from the tissue level to morphologies of individual fibrils. Charge neutralization enabled visualization of bare collagen fibrils and their connections. Nanoscale fibril and connection features may be obscured by conductive coatings. The unique collagen network of osteoid was also revealed by HIM. Osteoid was not imaged in the SEM studies that included immature tissue.26,27
Detailed comparisons of MCC nanoscale collagen network features revealed by HIM and by SEM in previous studies26-28 were not possible because there were substantial differences in sample preparation, including aldehyde fixation and application of conductive coatings to the SEM samples and differences in the relative locations at which the MCC structure was surveyed. Coatings inflate collagen fiber diameter measurements by approximately twice the coating thickness. The platinum coating was estimated to be up to 3 nm thick 26 ; thickness estimates were not reported for the gold coatings.27,28 Conductive coatings in standard protocols for biological samples may be 20 to 30 nm thick. 29 Because conductive coatings are not required for HIM, this modality has a substantial advantage for nanoscale collagen network investigation. Furthermore, only fibril thickness ranges (min, max) were reported for the SEM studies,26,27 so comparisons of the diameter distribution within and among the MCC zones and studies could not be made. However, despite these differences, similarities in overall structure were observed across species and ages, and these similarities are noted herein.
The fibrous zone is the structure that most distinguishes MCC from hyaline cartilage. 3 The HIM and TEM images revealed that the fibrous zone was composed of distinct layers of collagen bundles that were approximately orthogonal. This arrangement was also demonstrated in MCC of 8 week rats, 26 immature and mature cynomolgus monkeys, 27 and adult humans. 28 In the present study, the thicker layers were aligned in the anterior-posterior direction, the primary direction of condyle motion and shear loading. This structure is reflected in mechanical testing results where greater stiffness was measured in the anterior-posterior direction under tensile and dynamic shear protocols. 30 The fibrous zone fibrocytes had numerous cytoplasmic processes, which may form extracellular compartments that guide collagen bundle formation in a process similar to that in tendons and the cornea.31-33
The superficial 1 µm of the MCC fibrous zone consisted of a distinct surface region in which the alternating collagen layers were only a few fibrils thick. This layer may be analogous to the superficial layer, or lamina splendens, of hyaline cartilage. 34 Multiscale visualization revealed that the surface architecture was composed of fibrils with 2 distinct morphologies: longitudinal fibrils that were interwoven with a reticular network of smaller diameter fibrils. The role of this interwoven network in MCC function has yet to be determined.
Growth in length of the mandibular condyle occurs in the proliferative, mature, and hypertrophic zones through the process of endochondral ossification. 4 Research has largely focused on the substantial changes in cell size that drive growth; however, corresponding changes to the collagen network are required to enable the dramatic directional cell expansion while maintaining condyle mechanical integrity. Here, the HIM multiscale images revealed the result of this dynamic matrix remodeling process. The branched network, which was also seen in 8 week rat MCC, 26 became compacted into thin septae between adjacent hypertrophic cells, and it was eroded to accommodate cell expansion and to remove septae between adjacent cells, producing a compacted calcified scaffold for trabecular bone formation.
The layer of collagen fibrils deposited on the calcified cartilage scaffold was confirmed as osteoid by TEM and Movat’s pentachrome stain. The HIM, TEM, and pentachrome stain each revealed unique aspects of this very early osteochondral junction. The osteoid collagen fibrils were woven around numerous osteoblast cytoplasmic processes, creating a surface punctuated with numerous elliptical openings that narrowed to smaller diameter canals, with dimensions similar to those reported for rabbit cortical bone canaliculi. 35 However, the canals appear to be more closely spaced than those in resin cast and acid etched samples, suggesting that only a subset of these canals persists to become canaliculi in mature bone. Additional research is needed to better understand the early development of the lacunar-canicular system.
The goal of articular cartilage regeneration is to produce a durable repair structure that enables long term joint function. A key challenge has been an inability to readily visualize the 3D collagen network at the scale of its constituents and their assembly. Regeneration evaluation has typically relied on (immuno)histochemistry to identify the presence of cartilage constituents. Durability has been assessed by costly long-term animal experiments. Collagen network integration between the regenerated and surrounding cartilage has been identified as necessary for regeneration success. 36 However, fundamental questions regarding the process of and benchmarks for clinically effective repair remain. Cartilage structure determines its mechanical function;14,37 however, the manner in which structure and function evolve during regeneration is unknown. The extent to which the regenerated structure must replicate the highly anisotropic native structure to provide a clinical benefit is also unknown. Multiscale HIM visualization, in concert with TEM and (immuno)histochemistry, will enable researchers to address these questions and to advance the foundational understanding of MCC structural and functional development during regeneration.
The MCC collagen network is composed of a variety of collagen types.16,38,39 A limitation of this study was that fibril morphologies and size ranges could not be associated with collagen types. Future research to identify collagen types will improve our understanding of network assembly and the changes that occur with injury and disease.
In conclusion, the HIM visualization methods revealed the stratified MCC surface collagen network structure and the developing cartilage-subchondral bone junction researchers aim to regenerate, and they provide a means to evaluate the outcome of regeneration efforts. These data provide a baseline of comparison to better understand and evaluate collagen network changes that occur during MCC regeneration, as well as development, injury, and degeneration. This 3D, multiscale perspective on the MCC collagen network may stimulate new hypotheses regarding its development and the remodeling required to achieve the stratified adult structure and serve as a catalyst for evaluating MCC nanostructure at multiple points throughout development. This work is a step toward filling the critical gap in our foundational knowledge of the unique MCC collagen network.
Footnotes
Acknowledgments and Funding
The author acknowledges Carl Zeiss Microscopy, LLC, for access to the Orion® Plus HIM, Chuong Hyunh and Claus Burkhardt for HIM image acquisition, Kylie Martin for histology, and Nancy Smythe for TEM sample processing. This work was supported by NIH NIDCR K99DE023123 and NIGMS P30GM103331 and the AO Foundation.
Declaration of Conflict of Interests
The author declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical Approval
Ethical approval for this study was obtained from the Medical University of South Carolina Institutional Animal Care and Use Committee.