
Abstract
Objective
Meniscus tissue is composed of highly aligned type I collagen embedded with cartilaginous matrix. This histological feature endows mechanical properties, such as tensile strength along the direction of the collagen alignment and endurance to compressive load induced by weight bearing. The main objective of this study was to compare the fibrocartilage construction capability of different cell sources in the presence of mechanical stimuli.
Design
Synovial multipotent stem cells (SvMSCs) and meniscal chondrocytes (MCs) from immature and mature rabbits were maintained under similar conditions for comparative evaluation of growth characteristics and senescence tendency. The differentiation potential of cell sources, including fibrocartilage generation, were comparatively evaluated. To determine the capability of fibrocartilage generation, cultured cell sheets were rolled up to produce cable-form tissue and subjected to chondrogenic induction in the presence or absence of static tension.
Results
Although SvMSCs showed superior cell growth characteristics during in vitro cell expansion, senescence-associated β-galactosidase expression was consistently higher, compared with MCs. MCs showed glycosaminoglycan (GAG)-rich matrix formation during default in vitro chondrogenesis. While application of static tension significantly reduced GAG production, MCs continued to show robust tissue growth. SvMSCs showed inferior chondrogenic differentiation and diminished tissue growth in the presence of static tension.
Conclusions
While SvMSCs produced fibrous tissue during default in vitro chondrogenesis, their fibrocartilage generation potential in the presence of static tension was significantly lower, compared with MCs. Our results support evaluation of cellular response to tensile stimulus as a decisive factor in determining the ideal cell source for fibrocartilage reconstruction.
Introduction
Meniscus is a crescent-shaped cartilage tissues located between the femur and tibia in the lateral and medial side of the patellar joint. The meniscus exhibits differential histological microstructures in the inner and outer sides of the crescent shape associated with tissue functionality. The outer side of meniscus displays typical fibrocartilage morphology with highly aligned type I collagen throughout the tissue and blood vessels penetrating the outermost layer. The inner side of the meniscus partly includes hyaline cartilage characterized by a glycosaminoglycan (GAG)-rich region.1-3 Highly aligned collagen-rich matrix can bear high tensile loads and resist shape deformation caused by loads laterally delivered from the weightbearing inner part of meniscus.2-4 The GAG-rich region in the inner meniscus performs a similar function as hyaline cartilage and shares ~50% of the weight load with articular cartilage. 4 Meniscus tissues collected from large animals include tie fiber, an additional hierarchical structure that potentiates tensile modulus of tissue by radially traversing the tissue from the outer to inner side. 3 Fibrocartilage and hyaline cartilage share common histological features, such as low cellularity and an avascular nature. Therefore, following damage, regeneration rarely occurs.1-3,5 Meniscal injuries, such as meniscal tear, are treated via meniscectomy and require orthopedic surgery in ~50% cases. 3 Accumulating meniscus tissue damage can result in compromised function due to structural deformation, in turn, causing higher weight load to articular cartilage in the joint and, eventually, increased risk of early osteoarthritis development.2-4,6,7 In view of the importance of meniscus in prevention of early-onset osteoarthritis, several approaches have been developed to repair damaged meniscus tissue. Despite advances in tissue engineering approaches to formulate the unique microstructure of meniscus, the mechanical properties of in vitro reconstructed meniscus structures remain suboptimal compared with natural tissue, suggesting the need for mechanical stimulation to facilitate maturation and remodeling in vitro. Therefore, for reconstruction of meniscus, evaluation of the matrix generation capability of the cell source is essential to optimize response to mechanical stimulation. Results from a recent study showed that tensile stimulus induces highly arrayed collagen fiber alignment or increased collagen expression.8-10 However, collagen alignment by tensile stimulus was associated with reduced GAG-rich cartilaginous matrix production. 8 Fibrocartilage differentiation potential involves unique cell functionality of balanced cartilaginous matrix production, even in the presence of tensile stimulus. Furthermore, for materialization of transplantable meniscus tissue, functionality of the cell source is critical in terms of differentiation potential of both fibrocartilage and hyaline cartilage.
Synovial MSCs (SvMSCs) are postnatal multipotent stem cells (MSCs) that can potentially differentiate into chondrocytes, adipocytes, and myocytes. SvMSCs present a promising cell source for meniscus reconstruction, considering the source, synovium, is a type I collagen-rich fibrous tissue, and display significantly superior chondrogenic differentiation potential compared with bone marro–derived stem cells (BMSCs).11-16
In contrast, meniscal chondrocytes (MCs) are not MSC-like cells with lack of multilineage differentiation potential. Although SvMSCs and MCs show relatively low chondrogenic differentiation potential relative to articular cartilage chondrocytes, high type I collagen expression in these cell types supports their potential utility as a promising cell source for meniscus reconstruction.11,17 Several previous reports have shown higher chondrogenic potential of meniscal fibrochondrocytes (MCs) than SvMSCs.11,18,19
In earlier studies, MCs were cultured in polyglycolic acid (PGA)–hyaluronan 20 or GelMa scaffold 21 for reconstruction of tissue-engineered meniscus. Transplantation of PGA mesh scaffold seeded with rabbit MCs into rabbit joints promoted regeneration of the tissue with proteoglycan and collagen expression. 22
As for the available donor tissue for MC cell isolation, meniscus tissue fragment could be obtained during meniscectomy, involving removal of the meniscus tear to relieve pain and prevent further damage. In case of extensive meniscus damage, restoration of its function is essential to prevent early onset of degenerative joint disease. 23 The tissues obtained from meniscectomy may effectively serve as autologous donors for cell banking in preparation for meniscus reconstruction.
In this study, 2 promising cell sources, SvMSCs and MCs, were subjected to comparative evaluation in terms of fibrocartilage matrix production capability in response to tensile stimulus as well as basic cell characteristics, including in vitro expansion profiles and differentiation potentials. The comparative data suggest that MCs are the most ideal cell source for meniscus reconstruction.
Materials and Methods
Materials
All chemicals were purchased from Sigma-Aldrich (St. Louis, MO, USA) unless otherwise indicated.
Primary Culture of BMSCs
Primary cell cultures were generated from rabbit tissue after review and approval of the study (Approval Number: MCTTIACUC ASP 17-002/Approval Date: September 6, 2017) Institutional Animal Care and Use Committee (IACUC) of Biosolution Inc. Research Institute (Seoul, South Korea). BMSCs were isolated from femur and tibia of New Zealand white rabbits as described previously.24,25 For comparison of age-related characteristics, 4 rabbits in each age group (1 and 8 months old and weighing between 2 and 4.8 to 5 kg, classified as immature and mature, respectively), were subjected to cell yield examinations (n = 4). Isolated cells were plated at a density of 1 × 107 cells per 100 mm cell culture dish (Corning Life Sciences, Corning, NY, USA) and maintained in standard growth medium composed of α-minimum essential medium (α-MEM; Life Technologies, Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 5% fetal bovine serum (FBS; JRScientific Inc., Woodland, CA, USA), 1% penicillin/streptomycin (Welgene, Gyeongsangbuk-do, South Korea), 10−8 M dexamethasone, 2 mM
Primary Culture of SvMSCs and MCs
Synovial and meniscus tissues were collected from the patellofemoral joints of rabbits from each age group (immature and mature, n = 4). Each cell type was marked and traced for comparison with BMSCs from matching individuals. Isolated synovial tissues were washed more than 3 times with phosphate buffered saline (PBS) containing 2% penicillin/streptomycin and the attached blood vessels removed before digestion with 1% collagenase I (Worthington Biochemical, Lakewood, NJ, USA) for 4 hours in a humidified CO2 incubator at 37°C and 5% CO2. Isolated meniscus tissue samples were washed more than 3 times with PBS containing 2% penicillin/streptomycin (Welgene) and the attached connective tissues removed before cutting into tissue fragments 1-2 mm in size. Meniscus tissue fragments were digested with 1% collagenase I for 12 hours in a 37°C CO2 incubator. The isolated cell suspension with dissolved tissue was filtered through a cell strainer (Corning) with an additional volume of α-MEM. During this step, cell numbers were calculated to compare cell yield per mg tissue or total tissue (Supplementary Figure S1). After centrifugation at 1,500 rpm for 5 minutes, cell pellets were suspended in standard growth medium and plated at a density of 1 × 106 cells per 100 mm culture dish. The medium was changed every other day and maintained in a similar manner to BMSC cultures.
Cell Senescence Assay
Senescence-associated β-galactosidase (SA-β-gal) expressed by each cell type during passage progression between P4 and P8 was assessed using a specific β-Gal Assay Kit (Millipore, Billerica, MA, USA) according to the manufacturer’s instructions. Briefly, cells at each passage were fixed at 95% confluence and treated with the SA-β-gal substrate via overnight incubation in a 37°C incubator in a sealed state. After development of blue color confirmed under phase-contrast microscopy, the reagent was removed and rinsed twice with PBS. The number of β-gal-positive cells was compared between SvMSCs and MCs originating from a matching mature rabbit to reduce the errors caused by differential basal levels in each individual rabbit.
In Vitro Osteogenic Differentiation Potential
SvMSCs and MCs originating from the same mature rabbit were seeded into 6-well plates at a cell density of 2 × 105 cells/well at passage 3 and incubated overnight in standard growth medium. After overnight culture, the medium was switched to osteogenic-induction medium composed of α-MEM supplemented with 20% FBS, 10−8 M dexamethasone, 0.2 mM ascorbic acid, and 10 mM β-glycerophosphate. The medium was changed every 2 days for 4 weeks of in vitro osteogenic induction culture. Cultured cells were washed with PBS and fixed with 4% paraformaldehyde (PFA) in PBS for 2 hours at room temperature. To visualize deposition of calcium-rich matrix, culture plates were stained with filtered 2% Alizarin Red S in DW for 20 minutes. After staining, unbound dye was washed off with running tap water.
In Vitro Adipogenic Differentiation Potential
For evaluation of adipogenic differentiation potential, SvMSCs and MCs originating from the same mature rabbit were seeded into 6-well culture plates at a density of 2 × 105 cells/well at passage 3 and incubated overnight in standard growth medium. After overnight culture, the medium was changed to adipogenic induction medium composed of high-glucose Dulbecco’s modified Eagle medium (DMEM; Lonza, Basel, Switzerland) supplemented with 10% FBS, 1% penicillin/streptomycin, 10 µg/mL insulin, 10−7 M dexamethasone, 0.2 mM indomethacin, and 0.5 mM 3-isobutyl-1-methylxanthine. Following 2 days of culture in adipogenic induction medium, cells were incubated in adipogenic nutrition medium composed of DMEM supplemented with 10% FBS, 1% penicillin/streptomycin, 10 µg/mL insulin, and 10−7 M dexamethasone. The 2 adipogenic media were alternately replaced every 2 days. After 4 weeks of in vitro adipogenic induction, cultured cells were washed with PBS and fixed in 4% PFA for 2 hours at room temperature. Lipid droplets were visualized by oil red O staining.
In Vitro Chondrogenesis Assay for Hyaline Cartilage Construction
SvMSCs and MCs originating from the same mature rabbit (passage 3) were counted and added to a 15 mL conical tube (2.5 × 105 cells per tube). After seeding, cells were centrifuged at 1,500 rpm for 5 minutes at 4°C and incubated overnight in 1 mL growth medium, carefully avoiding turbulence. After overnight culture, the medium was changed to chondrogenic induction medium composed of high-glucose DMEM (Life Technologies) with 1% penicillin/streptomycin and supplemented with 10−7 M dexamethasone, 0.35 mM proline, 1× ITS-3, 0.3 mM ascorbic acid and 10 ng/mL TGF-β3 (PeproTech, Seoul, South Korea). After 4 weeks of in vitro chondrogenic induction, pellets were washed with PBS and fixed in 4% PFA for 2 hours at room temperature. Fixed pellets were paraffin-embedded and sectioned with a microtome to 5 µm thickness. GAG-rich cartilaginous matrix was visualized via safranin O staining.
In Vitro Chondrogenesis Assay under Tensile Stimulus Application
To evaluate production of cartilaginous matrix under tensile stimulus application, sheet-cultured cells were rolled up to form a cable shape and static tension applied during chondrogenesis using a specific static tension chamber. Briefly, rabbit SvMSCs and MCs originating from the same mature individual (passage 3) were seeded at a density of 1 × 107 cells/dish on type I collagen-coated 125 × 125 mm square-shaped dishes (SPL Life Science, Pocheon-si, Gyeonggi-do, South Korea) in growth medium. After reaching confluence, cells were maintained via daily medium changes without passage for 10 to 14 days. Confluent cells were carefully lifted at the edge of the square dish by scraping slowly to form a cable shape. Cell cables were mounted on an acrylic static tension chamber. The initial length of cell cables was 30 mm and 10% static tension was applied. The chondrogenic induction medium was changed twice a week during culture. After 4 weeks of in vitro chondrogenic induction, tissue samples were washed with PBS and fixed in 4% PFA for 2 hours at room temperature. Fixed tissues were paraffin-embedded and sectioned with a microtome to 5 µm thickness. Deparaffinized sections were subjected to hematoxylin and eosin (H&E), safranin O, and Masson trichrome staining.
DNA Quantitative Analysis
The DNA quantification assay was performed using a specific kit (Takara Bio Inc., Kusatsu, Shiga, Japan) for quantification of live cells per tissue following the manufacturer’s protocol. Briefly, tissues were incubated overnight at 37°C with proteinase K and RNase A. Dissolved tissues were reacted with Hoechst 33258 dye and DNA fluorescence measured on a Tecan ELISA reader (Spark Multimode Microplate Reader, Tecan Trading AG, Switzerland) with excitation at 346 nm and emission at 460 nm.
Quantitative Analysis of Cartilaginous Matrix Components
Quantification of Collagenous Matrix
Quantification of collagen generated during in vitro chondrogenesis under tensile stimulus application was performed using the Sircol Assay kit (Biocolor Life Science Assays, Carrickfergus, Northern Ireland, UK) according to the manufacturer’s protocol. Briefly, equally divided cell cable tissues were digested using Pepsin solution digest-all (Life Technologies by Thermo Fisher Scientific) overnight at 37°C. Following digestion, dissolved tissue solutions were mixed with Sircol dye and centrifuged at 12,000 rpm 10 minutes. Pellets were dissolved in alkali solution and absorbance at 555 nm analyzed with a spectrophotometer (Tecan Trading AG).
Quantification of Glycosaminoglycans
Quantification of GAGs in tissues subjected to in vitro chondrogenesis under tensile stimulus conditions was performed using a Blyscan Assay kit (Biocolor Life Science Assays) according to the manufacturer’s protocol. Briefly, equally divided cell cables were digested with 0.1 M papain solution. Dissolved GAG molecules in digested tissue solution were bound to 1,9-dimethylemthylene blue and absorbance at 656 nm analyzed with a spectrophotometer. A proportion of the papain-digested tissue solution was used for DNA fluorescence quantification assay based on Hoechst 33258 binding.
Statistical Analysis
Numerical data plotted on graphs were presented as mean ± standard deviation. Differences between 2 groups were compared using a 2-tailed paired t test. Statistical significance was accepted at P < 0.05. All experiments were repeated at least 3 times as specified in the figure legends.
Results
Comparison of Colony-Forming Unit–Fibroblasts Proportions in Primary SvMSCs and MCs
The frequencies of colony-forming unit–fibroblasts (CFU-Fs) in primary cultured SvMSCs and MCs were compared with those in BMSCs at passages 0 and 1. At passage 0, CFU-F numbers of BMSCs, SvMSCs, and MCs (1 × 103 cells in each group) were 0, 125.6, and 212.8 in immature animals and 1, 74.3, and 79 in mature animals, respectively. At passage 1, CFU-F numbers in BMSCs, SvMSCs, and MCs were 39, 132.2, and 77.8 in immature animals and 70.6, 178.7, and 123.8 per 1 × 103 cells in mature animals, respectively. SvMSCs and MCs from mature animals showed a higher proportion of CFU-Fs, compared with cells isolated from immature animals at passage 1 (Supplementary Figure S2). The colonies generated by SvMSCs and MCs isolated from mature animals were significantly bigger (2.41- and 13.94-fold difference, respectively) than those generated by cells from immature animals (Supplementary Figure S3). Although there was individual difference in terms of SvMSCs’ colony size, the tendency of SvMSCs’ producing relatively bigger colonies than MCs was consistently repeated in each animal (Supplementary Table S1). The colony size differences could be interpreted as the state of cellular activity, since the colony size is determined by cell population doubling number. 26 Our results indicate that SvMSCs and MCs from mature individuals are more active compared with the corresponding cell populations from immature individuals.
Comparison of In Vitro Cell Expansion Rate between SvMSCs and MCs in Immature and Mature Animals
SvMSCs and MCs were subcultured at 95% confluence and the cell number at each passage calculated to compare cumulative cell growth between the two sources. As the gold standard, BMSCs from matching animals were compared starting at passage 1, since BMSCs at passage 0 cannot be distinguished from mononuclear cells of hematopoietic lineages. SvMSCs displayed a higher in vitro expansion rate relative to MCs in both immature and mature animals. Comparative analyses between age groups showed higher expansion rates of both SvMSCs and MCs from mature animals relative to the corresponding cell sources from immature animals (Supplementary Figure S2). This in vitro expansion tendency was consistent with CFU-F size data (Supplementary Figure S1). Our results indicate that both cell types from mature individuals show excellent growth profiles and may be utilized for reconstruction of engineered tissues. Since the necessity for fibrocartilage reconstruction is higher in elderly individuals, further examinations proceeded with cells obtained from mature animals.
Comparison of Cell Senescence Tendencies of SvMSCs and MCs
Cell senescence characteristics according to passage progression were confirmed based on expression of SA-β-gal. Levels of SA-β-gal in SvMSCs and MCs showed a gradual increase until passages 6 and 7, respectively. At passage 7, SvMSCs showed a marked increase in SA-β-gal that continued to increase at passage 8, clearly indicative of senescence. Although SA-β-gal expression of MCs was significantly increased at passage 7, the number of SA-β-gal(+) MCs remained lower than that of SA-β-gal(+) SvMSCs at all passage time points ( Fig. 1 ), suggesting enhanced cell activity and stable functionality of MCs compared to SvMSCs.
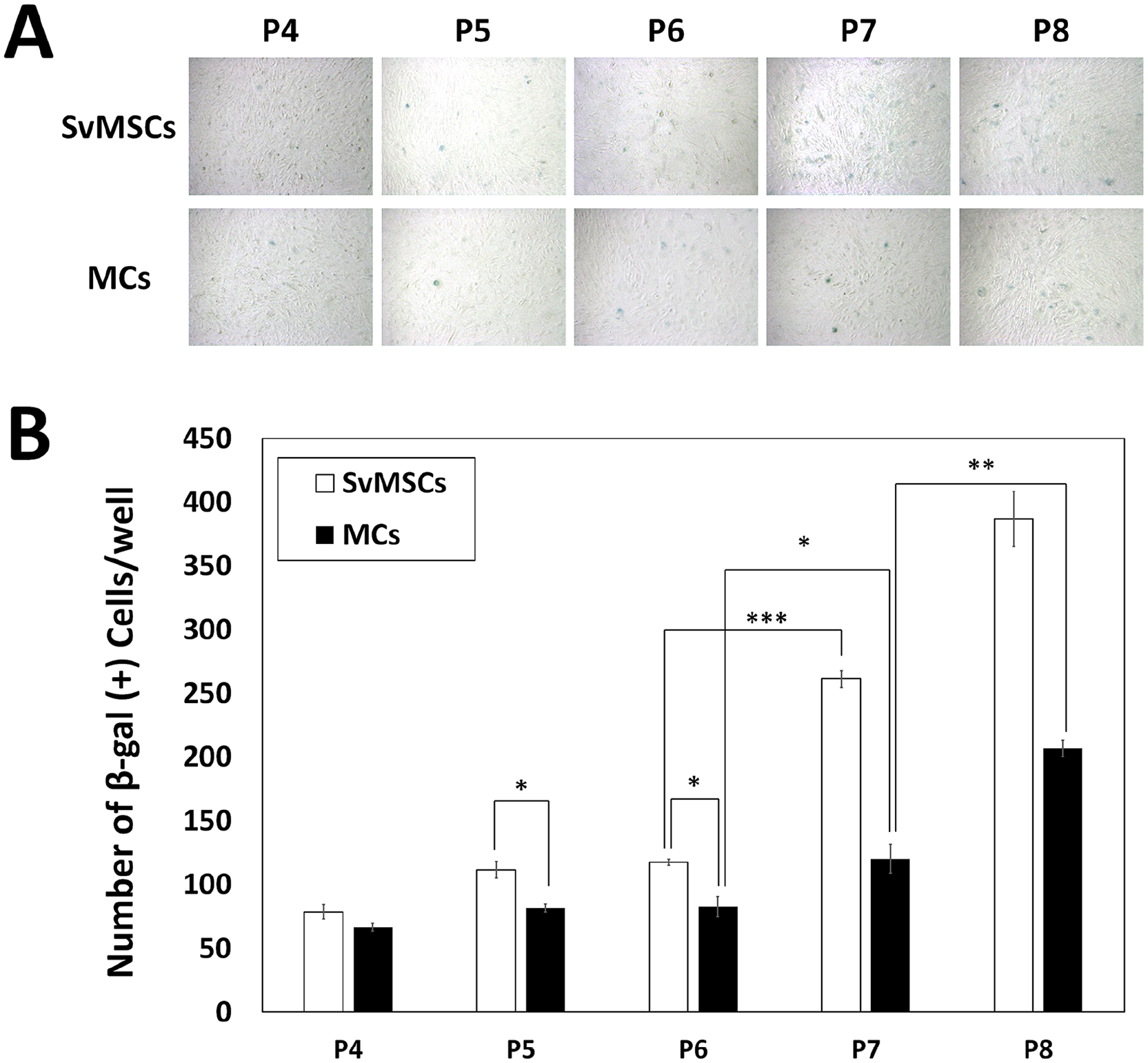
Cell senescence assay via senescence-associated β-galactosidase (SA-β-gal) staining over the course of passage progression in synovial mesenchymal stem cells (SvMSCs) and meniscal chondrocytes (MCs). The number of SA-β-gal(+) cells per well was compared at each passage (n = 3, *P < 0.05, **P < 0.01, ***P < 0.001).
Comparative Evaluation on In Vitro Differentiation Potential of SvMSCs and MCs
SvMSCs are classified as MSCs and show similarity to BMSCs in terms of in vitro expansion ability and multilineage differentiation potential in many previous reports.12-15 To compare the functional characteristics of SvMSCs and MCs, in vitro differentiation potential into osteogenic and adipogenic lineages was investigated. Following osteogenic induction, both cell types barely showed alizarin red–positive calcium-rich matrix formation ( Fig. 2A ). Quantification of the alizarin red–positive area via image analysis disclosed no significant differences between SvMSCs and MCs. As for in vitro adipogenesis, SvMSCs produced 2.689-fold greater Oil Red O-positive adipocytes, compared with MCs, as determined via image analysis ( Fig. 2B ).
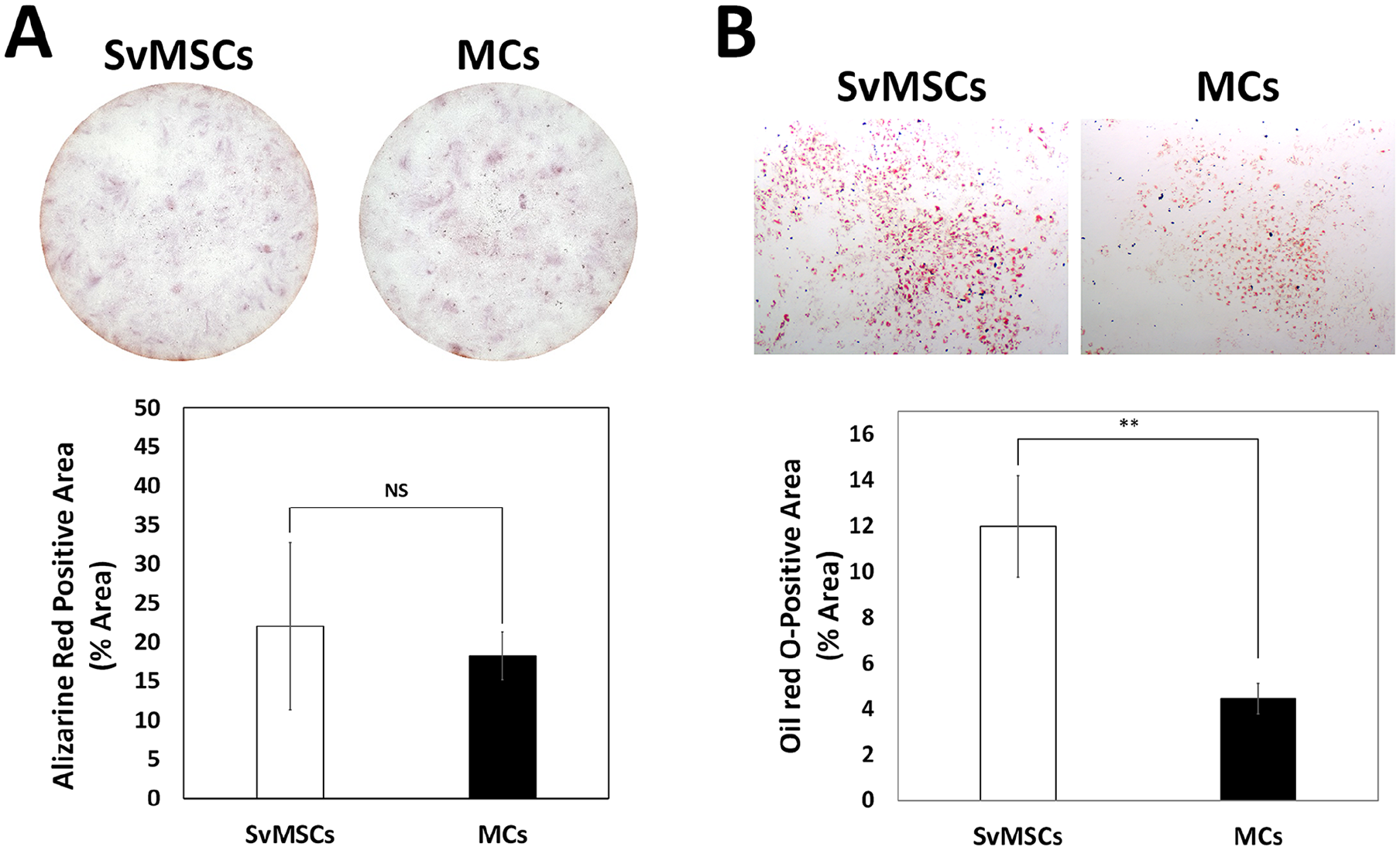
Comparison of in vitro differentiation potential of synovial mesenchymal stem cells (SvMSCs) and meniscal chondrocytes (MCs). (
Comparative Evaluation of Default Chondrogenic Potential in the Absence of Mechanical Stimulus
SvMSCs is reported to show superior chondrogenic differentiation potential compared with other MSCs from various tissue sources.6,13-15 Here, the chondrogenic differentiation potential of SvMSCs and MCs was comparatively evaluated using the pellet culture method. While the pellet size in both cell types increased during the differentiation period, MCs produced bigger pellets relative to SvMSCs, which were 1.28-, 1.31-, and 1.30-fold bigger at 1, 2, and 3 weeks of chondrogenic differentiation, respectively ( Fig. 3A and B ). Histological staining showed that MCs started to produce GAG-rich cartilaginous matrix at 2 weeks of chondrogenic induction while SvMSCs failed to form a GAG-rich matrix at all time points ( Fig. 3C and D ). Based on trichrome staining, matrix produced by both cell types were devoid of fibrous matrix up to 2 weeks of chondrogenic differentiation. At 3 weeks of chondrogenic induction, SvMSCs started to show trichrome-positive fibrous tissue formation while MCs lacked the trichrome-positive area ( Fig. 3E ). At 4 weeks of chondrogenic induction, SvMSC pellets obviously formed fibrous tissue while MC pellets showed greater staining intensity of safranin O–positive GAG-rich matrix. Our findings indicate that MCs maintain superior differentiation potential toward hyaline cartilage while SvMSCs resulted in fibrous tissue formation over 4 weeks of chondrogenic induction.
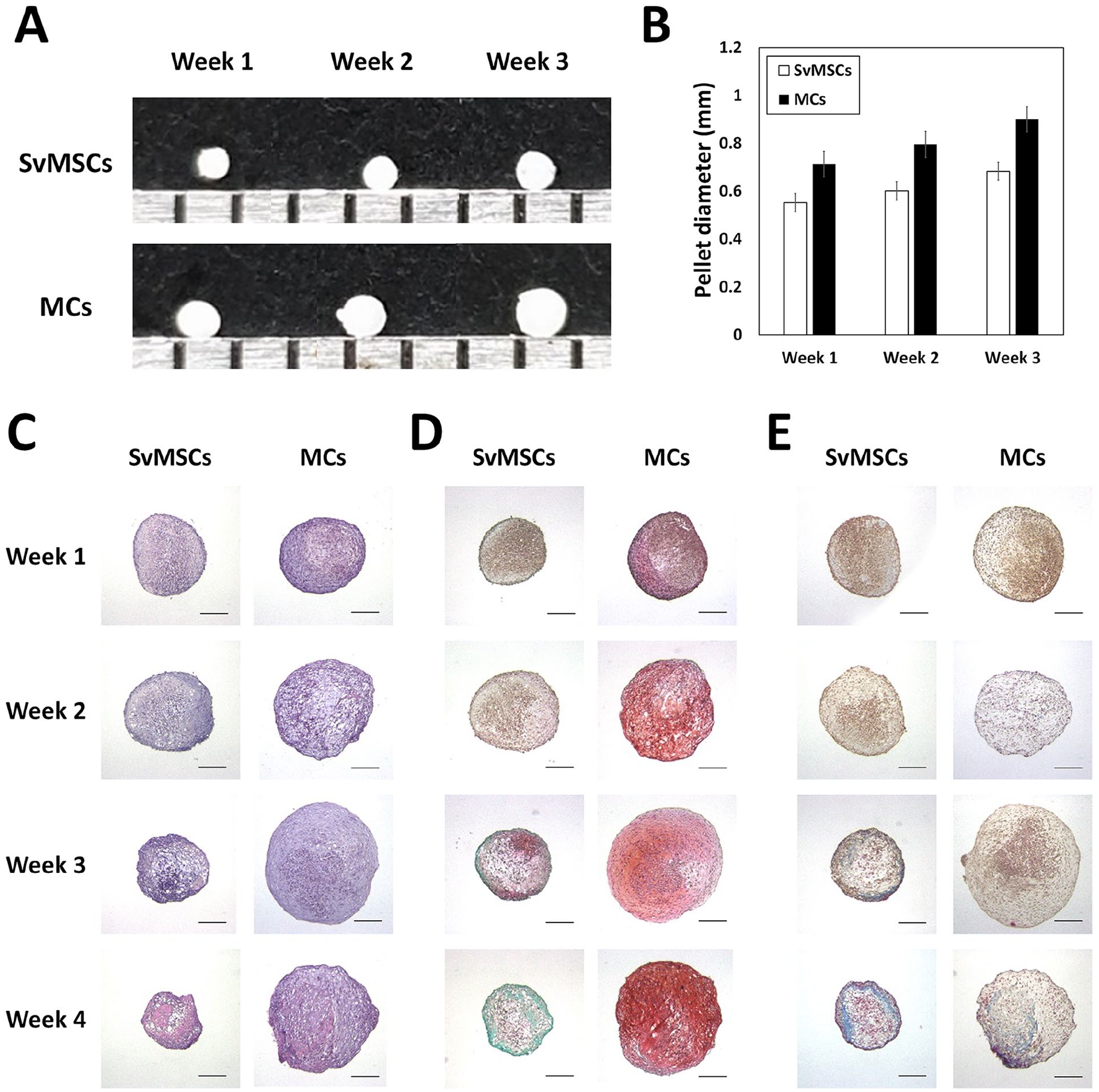
Comparison of in vitro chondrogenic differentiation potential of synovial mesenchymal stem cells (SvMSCs) and meniscal chondrocytes (MCs). The chondrogenic pellets produced by SvMSCs and MCs were compared in terms of size (
Comparative Evaluation of Fibrocartilage Generation Potential with Application of Tensile Stimulation
The microstructure of fibrocartilage involves uniaxial tissue alignment, which can be effectively induced via application of mechanical stimulus. However, mechanotransduction induced by tensile stimulus is reported to inhibit chondrogenic induction.27,28
To compare the cartilaginous matrix production capability of the different cell sources during tissue alignment induction by mechanical stimulation, sheet-cultured SvMSCs and MCs in growth media were rolled up to generate cable-form tissue (Supplementary Figure S6) and subjected to chondrogenic induction with tensile mechanical stimulation by 10% extension of the original length using a static tension chamber (Supplementary Figure S3). The thickness of tissue formed by MCs increased during the chondrogenic culture period while the tissue formed by SvMSCs showed no significant differences in thickness ( Fig. 4A and B ). In contrast to pellets of cultures subjected to chondrogenic induction without tensile stimulus ( Fig. 3 ), cable-form tissues generated with SvMSCs and MCs subjected to 10% tensile stimulus did not show high-intensity safranin O(+) GAG-rich cartilaginous matrix formation ( Fig. 4C ). On the other hand, the blue color in the trichrome-stained section was indicative of fibrous matrix ( Fig. 4D ). Although SvMSCs formed trichrome(+) fibrous tissue via default chondrogenesis ( Fig. 3E ), intense staining was not observed in tissues subjected to chondrogenesis with tensile stimulation ( Fig. 4D ). Notably, MCs generated more intensive trichrome(+) fibrous tissue under tensile stimulation, compared with SvMSCs ( Fig. 4D ). For numerical comparison, equally divided tissue fragments were subjected to quantitative analysis for DNA, collagen, and GAG. Cell numbers predicted via DNA quantification showed no significant changes during chondrogenic induction ( Fig. 5A ). Quantification of collagen content revealed a significant increase in collagen production by tissues formed by MCs during in vitro chondrogenic induction with tensile stimulation while tissues formed by SvMSCs showed no differences in collagen content at the indicated time points ( Fig. 5B ). In terms of GAG content, tissues formed by SvMSCs showed a slight decrease at the time points examined while tissues formed by MCs displayed a gradual increase in GAG amounts during in vitro chondrogenesis with tensile stimulation ( Fig. 5C ). With increasing GAG quantities over the examined time course in MC-derived tissues, tissue thickness was increased ( Fig. 4A ), which may have resulted in the density of matrix molecules per unit volume of MC-derived tissues remaining almost constant over the course of the experiment. This finding may explain why increased GAG production was not observed as more intensive safranin O staining in histological analysis ( Fig. 5C ).
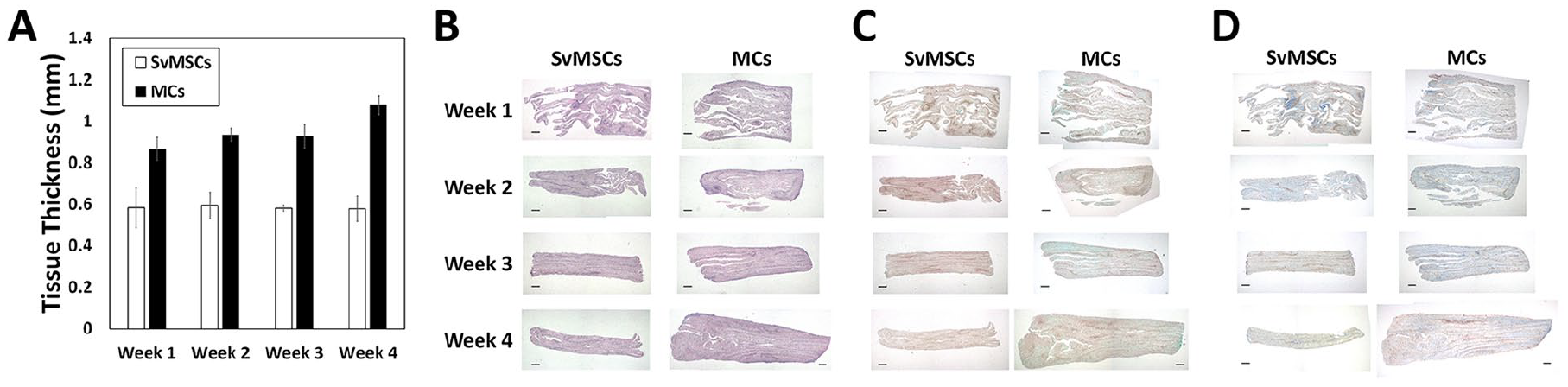
Comparison of the chondrogenic capabilities of synovial mesenchymal stem cells (SvMSCs) and meniscal chondrocytes (MCs) on application of tensile stimulation. Changes in tissue thickness were measured at time points each week (
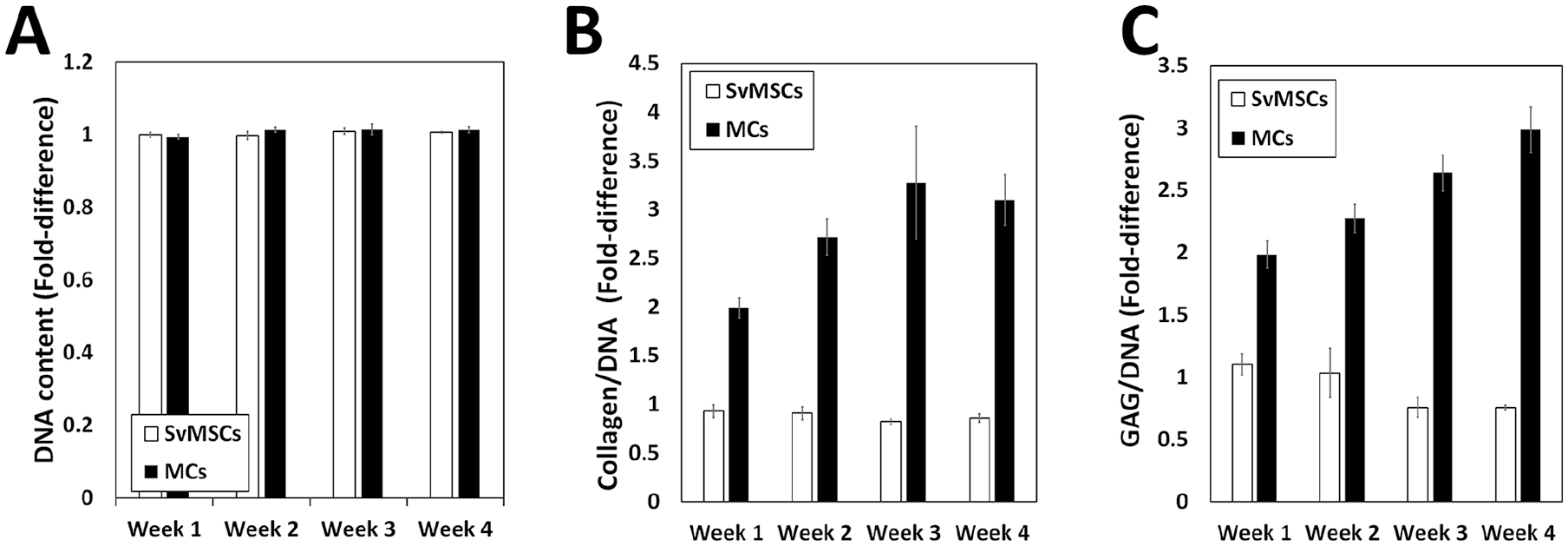
Analysis of changes in tissue components at time-points at each week in tissues subjected to fibrocartilage differentiation with tensile stimulation. Tissue fragments were divided into groups and subjected to Hoechst-based DNA quantification (
Discussion
Because of the technical challenges in reconstructing tissue-engineered meniscus, replacement with an allogeneic meniscus from cadaver donors is considered the best clinical practice for patients with severely damaged meniscus with no signs of osteoarthritis. 4 Several trials have been performed to reconstruct meniscus tissue with various cell sources and tissues, among which the most potent tissue construction strategies include mechanical training. 29 Application of tensile stimulus can promote fibrous tissue formation by inducing tissue alignment and type I collagen production. However, tensile stimulus activates mechanotransduction and induces differential responses according to cell type, such as an inhibitory effect on chondrogenesis in chondrocytes27,31 and stimulation of osteogenesis in MSCs.28,30 Therefore, cell responses to mechanical stimulation should be fully considered before utilization for meniscus reconstruction.
SvMSCs have been shown to possess MSC-like characteristics.11-16 In the current investigation study, both SvMSCs and MCs did not display alizarin red S staining after in vitro osteogenesis as observed in a previous report 17 and SvMSCs showed slightly higher adipogenic differentiation potential. In terms of chondrogenic differentiation, MCs were consistently superior to SvMSCs in producing cartilaginous matrix. Consistent with the finding that cell sources with low chondrogenic potential show fibrous tissue formation during in vitro chondrogenesis even without tensile stimulation,32,33 SvMSCs produced highly fibrous tissues with extremely low GAG formation during induction of hyaline cartilage differentiation ( Fig. 3D ). Despite the highly fibrous nature of SvMSCs during hyaline cartilage differentiation, these cells failed to show robust fibrous tissue growth during fibrocartilage differentiation in chondrogenic medium with application of 10% static tension ( Fig. 4B-D ).
These results indicate that evaluation of source cell functionality to achieve maximal mechanical load-bearing capability is essential for optimization of functional tissue engineering. 34 For efficient construction of tissues with mechanical functions, determination of the tissue formation characteristics of source cells in response to mechanical stimuli is critical, not only for the tissue construction process but also to replicate their response to the continuous mechanical stress imposed by the local environment following transplantation.
To elucidate the characteristics of in vitro–expanded cells, we examined expression of type I and type II collagen as markers for dedifferentiation and chondrogenic cells, respectively (Supplementary Figure S5). Over the course of passage progression, mature SvMSCs showed increased type I collagen expression and mature MCs showed a significant increase in the number of type I collagen–expressing cells along with decreased intensity of type II collagen expression. Interestingly, SvMSCs showed increased type II collagen intensity in the immature cell group and increased number of type II collagen–expressing cells in the mature cell group, consistent with a previous report. 35 Although SvMSCs displayed increased type II collagen expression at later passages, the chondrogenic potential of these cells was insufficient to produce a GAG-rich matrix ( Fig. 3 ).
In general, cells isolated from younger animals grow faster and show higher activity compared with those isolated from older animals. In our study, cells isolated from immature animals produced more colonies at primary culture (P0) in the CFU-F assay. However, once established, cells isolated from mature animals produced greater numbers and sizes of colonies than cells isolated from immature animals at passage 1 (Supplementary Figures S2 and S3). These characteristics may account for the higher number of cells from mature animals in terms of accumulative cell growth. Cell functionality can be enhanced if mechanical stress exerts beneficial effects by stimulating activities, such as survival, proliferation, and matrix production. In fact, mechanical stress experienced during the postnatal development process activates chondrocytes to acquire functionality resulting in tissue maturation and property changes. 36
Although no direct comparison of the responses of SvMSCs and MCs to mechanical stimuli has been reported in the literature, comparison of chondrogenic potentials of the 2 cell types revealed consistency with previous reports. Pellets of human meniscal fibrochondrocyte cultures showed better cartilaginous matrix formation based on GAG production than synovium-derived MSCs. 18 Human synovium-derived MSCs are additionally phenotypically unstable with low expression of type II collagen and Sox-9.11,19
In the current study, matrix production characteristics in response to static tension were examined in a simple acrylic chamber designed for small-scale tissue cultivation. To generate functional tissue-engineered meniscus, culture conditions need to be optimized. First, the mechanical dose should allow sufficient cartilaginous matrix formation while maintaining tissue alignment to limit the chondroinhibitory effect of mechanotransduction.27,28,37 Second, differential application of combined mechanical stimuli may be required to materialize the tissue alignment and replicate the unique tissue characteristics of meniscus. 29 Third, scaling up cultivation of tissue-engineered cartilage devoid of the necrotic inner core caused by undernutrition remains a considerable challenge. Therefore, several additional strategies require consideration to generate transplantable functional meniscus tissue using the tissue engineering approach.
To overcome the challenges in meniscus reconstruction, utilization of the optimal cell source is critical to reproduce target tissue properties. In our experiments, SvMSCs failed to show tissue growth during in vitro chondrogenesis while MCs showed robust tissue growth with GAG-rich cartilaginous matrix formation, even on application of tensile stimulus. Our collective findings support the utility of MCs as a suitable cell source for meniscus reconstruction.
Supplemental Material
R2_-_Supplementary_information – Supplemental material for Comparative Evaluation of Synovial Multipotent Stem Cells and Meniscal Chondrocytes for Capability of Fibrocartilage Reconstruction
Supplemental material, R2_-_Supplementary_information for Comparative Evaluation of Synovial Multipotent Stem Cells and Meniscal Chondrocytes for Capability of Fibrocartilage Reconstruction by Jisoo Lee, Seoyoung Jang, JunPyo Kwon, Tong In Oh and EunAh Lee in CARTILAGE
Footnotes
Supplemental Material
Author Contributions
JL: Contribution to experiment, analysis, drafting of the article. SJ: Contribution to experiment, analysis, drafting of the article. JPK: Contribution to experiment. TIO: Contribution to concept and design, interpretation of the data, drafting of the article, final approval of the article. EL: Contribution to concept and design, experiment, analysis, interpretation of the data, drafting of the article, final approval of the article.
Acknowledgments and Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Basic Science Research Program through the National Foundation of Korea (NRF) funded by the Ministry of Education (2018R1D1A1A02047934), Basic Science Research Program through the National Foundation of Korea (NRF) funded by the Ministry of Science and ICT (2018R1D1A1B07047508), and a grant of the Ministry of Health and Welfare of Korea (HI18C2435).
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical Approval
This study was approved by the Institutional Animal Care and Use Committee (IACUC) of Biosolution Inc. Research Institute (Seoul, South Korea) (Approval Number: MCTTIACUC ASP 17-002/Approval Date: September 6, 2017).
Animal Welfare
The present study followed international, national, and/or institutional guidelines for humane animal treatment and complied with relevant legislation.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.