
Abstract
Objective
To characterize the immunolocalization of clusterin in the repair cartilage of patients having undergone autologous chondrocyte implantation (ACI) and evaluate correlation to clinical outcome.
Design
Full-depth core biopsies of repair tissue were obtained from 38 patients who had undergone ACI at an average of 18 ± 13 months previously (range 8-67 months). The biopsies were snap frozen, cryosectioned, and clusterin production immunolocalized using a specific monoclonal clusterin antibody and compared with normal and osteoarthritic cartilage. Clinical outcome was assessed from patients preoperatively, at the time of biopsy, and annually postoperatively.
Results
Intensity of immunostaining for clusterin decreased with age in healthy cartilage tissue. Clusterin was detected to a variable degree in 37 of the 38 ACI cartilage biopsies, in single and clustered chondrocytes, in the pericellular capsule and the cartilage extracellular matrix, as well as the osteocytes and osteoid within the bone. Chondrocytes in hyaline repair tissue were significantly more immunopositive than those in fibrocartilage repair tissue. Clinical outcome improved significantly post-ACI, but did not correlate with the presence of clusterin in the repair tissue.
Conclusions
These results demonstrate the presence of clusterin in actively repairing human cartilage and indicate a different distribution of clusterin in this tissue compared to normal cartilage. Variability in clusterin staining in the repair tissue could indicate different states of chondrogenic differentiation. The clinical significance of clusterin within repair tissue is difficult to assess, although the ideal functioning repair tissue morphology should resemble that of healthy adult cartilage.
Introduction
Clusterin, a secreted heterodimeric glycoprotein, has previously been detected both by immunolocalization and in situ hybridization within the superficial layer of articular cartilage,1-3 in addition to many other tissue–fluid boundaries in the body. Although its specific function remains unclear, clusterin appears to be involved in many biological processes ranging from protection against stress and apoptosis, to lipid transportation.4,5 Expression of clusterin in superficial chondrocytes of articular cartilage has been suggested to regulate chondrocyte apoptosis in osteoarthritic tissue. 3 There is in vitro evidence that the state of differentiation of chondrocytes may influence the production of clusterin. Equine chondrocytes isolated from the superficial layer of cartilage lost production of clusterin following population expansion in culture but this was resumed as the cells were redifferentiated in 3-dimensional alginate cultures.2,6 Additionally, it has been suggested that the production of clusterin has the potential to be used as a marker for superficial chondrocytes because of their high expression of clusterin compared with deep zone chondrocytes, in both native tissue1-3 and tissue-engineered constructs.2,6
Because of the limited self-repair capability of hyaline cartilage, autologous chondrocyte implantation (ACI) has become an established method for the treatment of symptomatic full-depth chondral and osteochondral defects. 7 In preparation for treating the patients, their chondrocytes are isolated and culture expanded in vitro during which time they dedifferentiate. 8 Following their return to the patient and implantation beneath a periosteal or collagen patch, it is expected that the cells redifferentiate into a chondrocyte phenotype. The quality of repair tissue formed post-ACI is likely to dictate the clinical outcome for the patient, but the process by which repair tissue is formed is little understood. By investigating the localization of clusterin in healthy, diseased, and repairing cartilage, we may begin to understand the repair processes and determine if clusterin is important to the development of the layered structure of cartilage that gives it its specific properties for its required function.
Patients undergoing ACI in our center are offered an arthroscopic follow-up at approximately 12 to 18 months posttreatment with ACI, during which time a biopsy of the repair cartilage is obtained; sometimes, however, patients do present in clinic outside these times. We have analyzed these biopsies for the presence and distribution of clusterin, compared it with that in “normal” articular cartilage and assessed if there was any correlation to clinical outcome. We have also examined the distribution of collagen types I and II in repair tissue and compared it with that of clusterin.
Methods
Patients and ACI Procedure
Repair tissue was obtained from 38 fully informed and consented patients, who underwent ACI (9 on the lateral femoral condyle, 23 on the medial femoral condyle, 2 on the trochlea, 1 on the patella, and 3 on the talus) as part of an ethically approved study. ACI was performed as described by Brittberg et al. (1994) 9 using a two-stage procedure, where the first stage required arthroscopic removal of a small piece of macroscopically healthy cartilage, usually from either the intercondylar notch or trochlea. Chondrocytes were isolated from the harvested cartilage in the GMP (Good Manufacturing Practice)–approved John Charnley Laboratory and cultured in autologous serum. 10 After an average of 21 days in culture, the autologous chondrocytes were implanted beneath a periosteal or collagen (Chondro-Gide, Geistlich Pharma AG, Wolhusen, Switzerland) membrane, which was stitched and glued with fibrin over the defect.
Postoperative Tissue Biopsies
Core biopsies of repair tissue were obtained arthroscopically from the above 38 patients using a juvenile bone marrow biopsy needle, as close as possible to the centre of the treated defect to include the cartilage repair tissue and where possible, the subchondral bone. Lysholm scores, a measure of knee function, 11 were obtained pre- and postoperatively at the time of biopsy and annually thereafter for the 35 knee patients. The score ranges from 0 (poor outcome) to 100 (best outcome). Ankle scores were used in place of Lysholm scores as appropriate.
The biopsies (1.8-mm diameter and full depth) were snap-frozen in liquid nitrogen, embedded in optimal cutting temperature compound (Tissue-Tek, Zoeterwoude, The Netherlands) then cryosectioned to produce 7-µm thick sections, which were collected onto poly-
Histology
Frozen sections were defrosted and stained with either hemotoxylin and eosin to evaluate general morphology of the tissue or safranin O/fast green and/or toluidine blue, to indicate glycosaminoglycan content as per standard protocols. 12 The normal cartilage was then assessed for signs of degeneration using the Mankin score 13 (total score 0-14). A higher score represents a greater degree of degenerative change, with a score of 0 to 1 considered acceptable for normal healthy tissue.
Sections were viewed under polarized light microscopy to assess collagen organization within the tissue to distinguish between hyaline cartilage and fibrocartilage. The quality of the repair tissue was assessed under normal light microscopy and scored using both the International Cartilage Repair Society–II (ICRS-II) visual analogue scale 14 (each of 14 parameters scored 0-10) and the OsScore 15 (total score 0-10). A higher score represents better quality histology for both scoring systems.
Immunohistochemistry
Clusterin production was immunolocalized using a monoclonal antibody raised against human clusterin (1:200, G7), as described previously.2,16 In brief, cryosections were formalin fixed for 10 minutes before blocking endogenous peroxidase activity with 0.3% hydrogen peroxide in methanol for a further 10 minutes. Additional blocking was performed with 5% bovine serum albumin prior to an overnight incubation with the anti-clusterin antibody at 4°C. Sections were then incubated with the secondary goat anti-mouse biotinylated antibody (Vectastain Elite ABC kit, Vector Laboratories, Peterborough, UK).
For collagen types I and II, sections were incubated with hyaluronidase for 2 hours to aid exposure of antigens, prior to fixing in formalin for 10 minutes. Sections were then incubated for an hour in the presence of either anti-collagen type I (1:500, clone I-8H5, MP Biomedicals, Cambridge, UK) or type II (1:10, clone CIIC1, Developmental Studies Hybridoma Bank, Iowa City, IA) before incubating with the secondary goat anti-mouse biotinylated antibody (Vectastain Elite ABC kit). Endogenous peroxidase activity was then blocked using 0.3% hydrogen peroxide in methanol.
For detection of the proliferation marker Ki67, all sections of repair tissue with cell clusters present were briefly fixed in ice cold acetone prior to blocking endogenous peroxide activity with 0.3% hydrogen peroxide in methanol for 20 minutes. An anti-human Ki67 nuclear antigen antibody (1:100, clone MM1, Novacastra, Peterborough, UK) was applied to the slides for an hour prior to a secondary goat anti-mouse biotinylated antibody (Vectastain Elite ABC kit).
For all immunoprocedures, steps were performed at room temperature (unless otherwise stated) and sections were washed 3 times with PBS between steps. All labeling was enhanced with streptavidin-peroxidase (Vectastain Elite ABC kit, Vector Laboratories, Peterborough, UK) and visualized with diaminobenzadine before dehydrating and mounting. Adjacent sections were stained with an isotype-matched murine IgG1 (Dako, Cambridge, UK) in place of the primary antibody as a normal control. For Ki67 immunostaining, sections of breast carcinoma were used as a positive control.
Analysis
To analyze the localization and distribution of clusterin, the biopsies were assessed in a semiquantitative manner as two parts: cartilage and bone ( Fig. 1 ). This was performed by one (blinded) observer. The complete section of cartilage was assessed in 3 regions: the surface/superficial zone (upper 25% of the cartilage), the mid zone (central 50% of the cartilage), and the deep zone (lower 25% of the cartilage). The intensity of staining was recorded (0 = none, 1 = weak, 2 = medium, 3 = strong) for the extracellular matrix, single chondrocytes, and pericellular capsules for each of the 3 regions of cartilage. In addition, for the articulating surface (upper 5-10 µm) was assessed as this was the most striking location of clusterin in normal tissue.1 -3 Cell clusters (groups of 3 or more chondrocytes) in each region and their intensity of staining were also noted. The bone was assessed for the localization of clusterin in the osteoid and osteocytes.
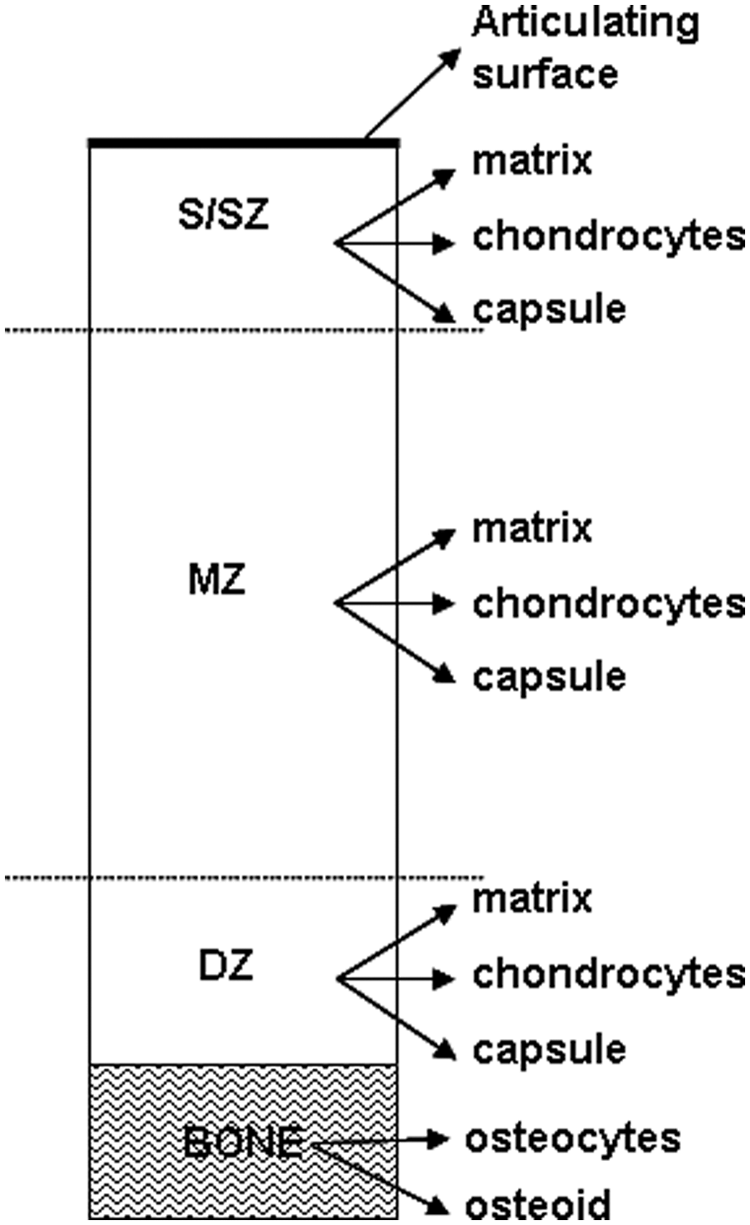
Schematic diagram of a core biopsy defining the regions and parameters scored for clusterin. The cartilage was assessed in the following zones: surface/superficial zone (S/SZ), mid zone (MZ), deep zone (DZ) and the bone as a separate entity.
Intensity scores for the extracellular matrix from the 3 cartilage regions and the upper 5 to 10 µm were totaled to yield an overall extracellular matrix score. Likewise, intensity scores for the single chondrocytes and pericellular capsules from each 3 cartilage regions were totaled to yield an overall chondrocyte score. A total cartilage clusterin score was calculated by summating the total extracellular score and the total chondrocyte score. Clustered chondrocyte scores for each region were totaled to yield the clustered chondrocyte score. For the bone, the osteoid and osteocyte intensity scores were totaled to yield the bone score.
For collagen types I and II distribution, positive extracellular matrix staining was recorded as a percentage of the total area of the cartilage. Extracellular matrix immunostaining for clusterin was also calculated in the same way to allow comparison with collagen types I and II staining.
Statistical Analysis
Statistical differences between clusterin scores for the various parameters and clinical outcomes were tested for via a Mann–Whitney U test and correlations were tested for with a Spearman’s rank correlation using a software program (Analyse-it Software Ltd, Leeds, UK). Statistical significance was considered when P < 0.05. Values are quoted as means ± standard deviations.
Results
The average age of the patients at the time of ACI was 36.8 ± 9.5 years (range 17-58 years) with a mean treated defect size of 4.3 ± 3.1 cm2 (range 1-12 cm2). Core biopsies of repair tissue were obtained arthroscopically from these patients at 8 to 67 months postoperatively (average 18 ± 13 months), depending on when patients presented in clinic for routine or other follow-up appointments. Of the 38 biopsies included in this study, 33 included the underlying subchondral bone. The morphology of the repair tissue was predominantly hyaline cartilage in 6 biopsies; 3 biopsies were a mixture of both hyaline and fibrocartilage and 29 were mainly fibrocartilage. The normal samples used for comparison were all of a hyaline morphology and all except the 11-year-old patella included the subchondral bone. All control comparative sections had a Mankin score of 0 to 2, indicating very little or no degenerative change.
Immunolocalization of Clusterin in Healthy Cartilage
In all of the “normal” control samples, there was a strong band of positive immunolocalization for clusterin observed along the very upper 5 to 10 µm of the articulating surface ( Fig. 2 ), the intensity of which reduced with age. In the 11- and 21-year-olds, the extracellular matrix below this was intensely positive for clusterin in the surface/superficial zone to a depth of ~40 µm. In the remaining control samples, however, no staining was detected elsewhere in the extracellular matrix ( Fig. 3A ). In all but one of the control samples there was staining of the chondrocytes and peri-cellular capsules in the surface and mid zones, but not in the deep zone. The intensity of the staining was strongest in the cartilage from the 11-year-old.
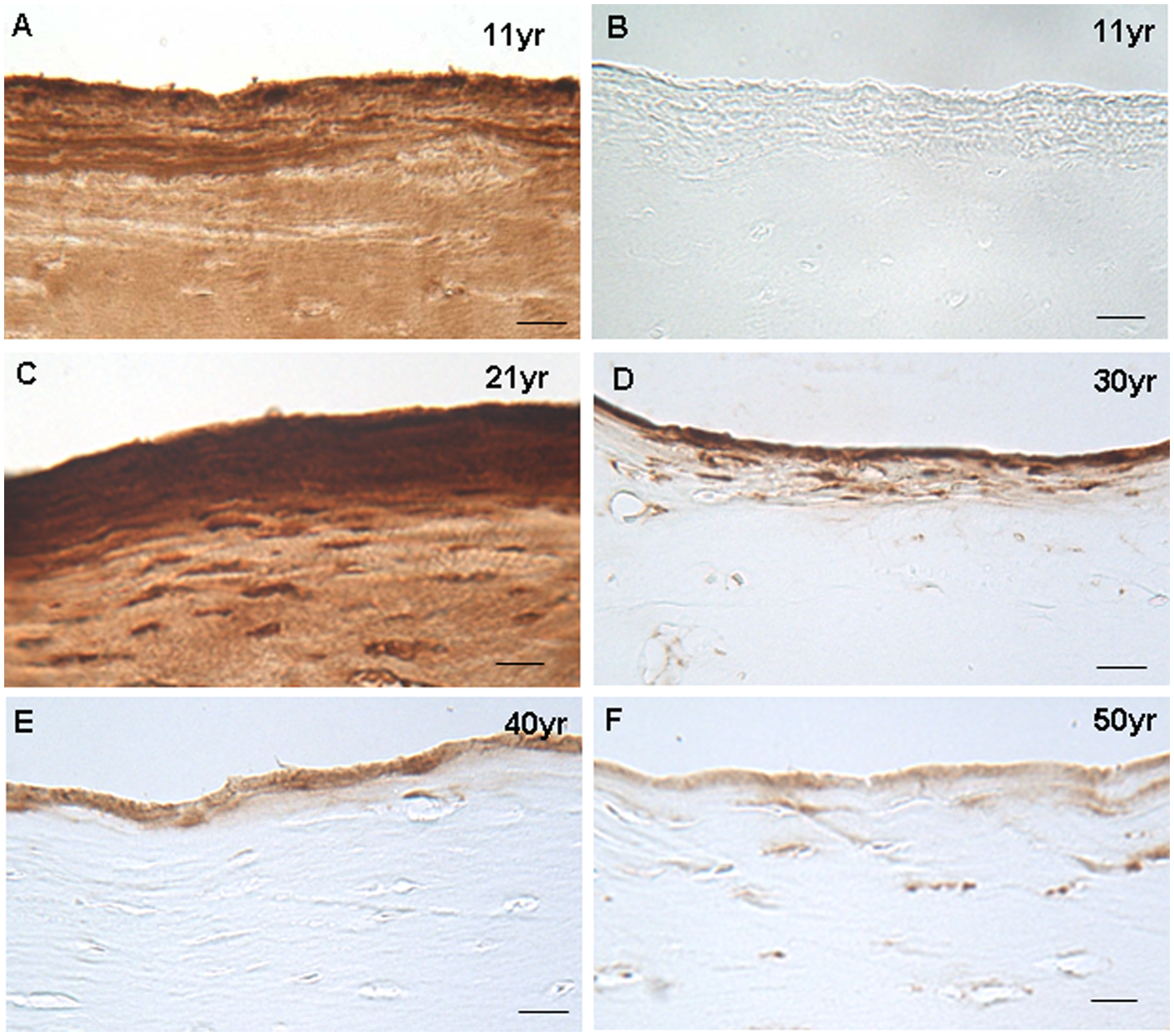
Immunolocalization of clusterin in the upper 5- to 10-µm of the surface zone in a sample from the patella of an 11-year-old (
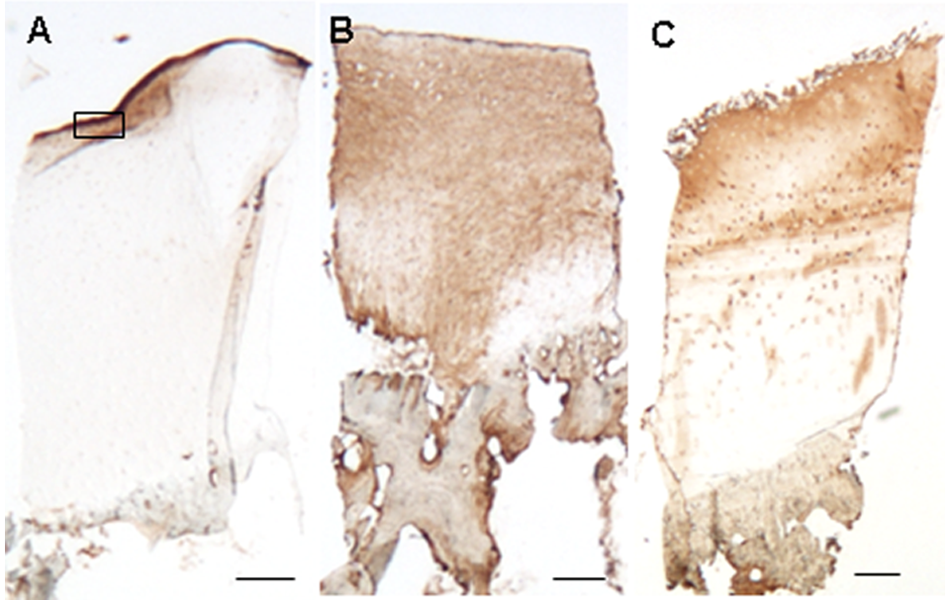
Lower power micrographs showing the immunolocalization of clusterin in the extracellular matrix of full-depth cartilage cores from a 21-year postmortem sample (
Although chondrocyte clusters were observed in the mid and deep zone of the 40- and 50-year-old PM samples, only those in the mid zone of the 50-year-old sample exhibited positive cellular staining for clusterin. In the bone there was clusterin immunopositivity in the osteoid of the 21-, 30-, and 40-year-old PM samples and in the osteocytes in only the 40- and 50-year-old PM samples.
Immunolocalization of Clusterin, Ki67, and Collagen Types I and II in Repair Cartilage
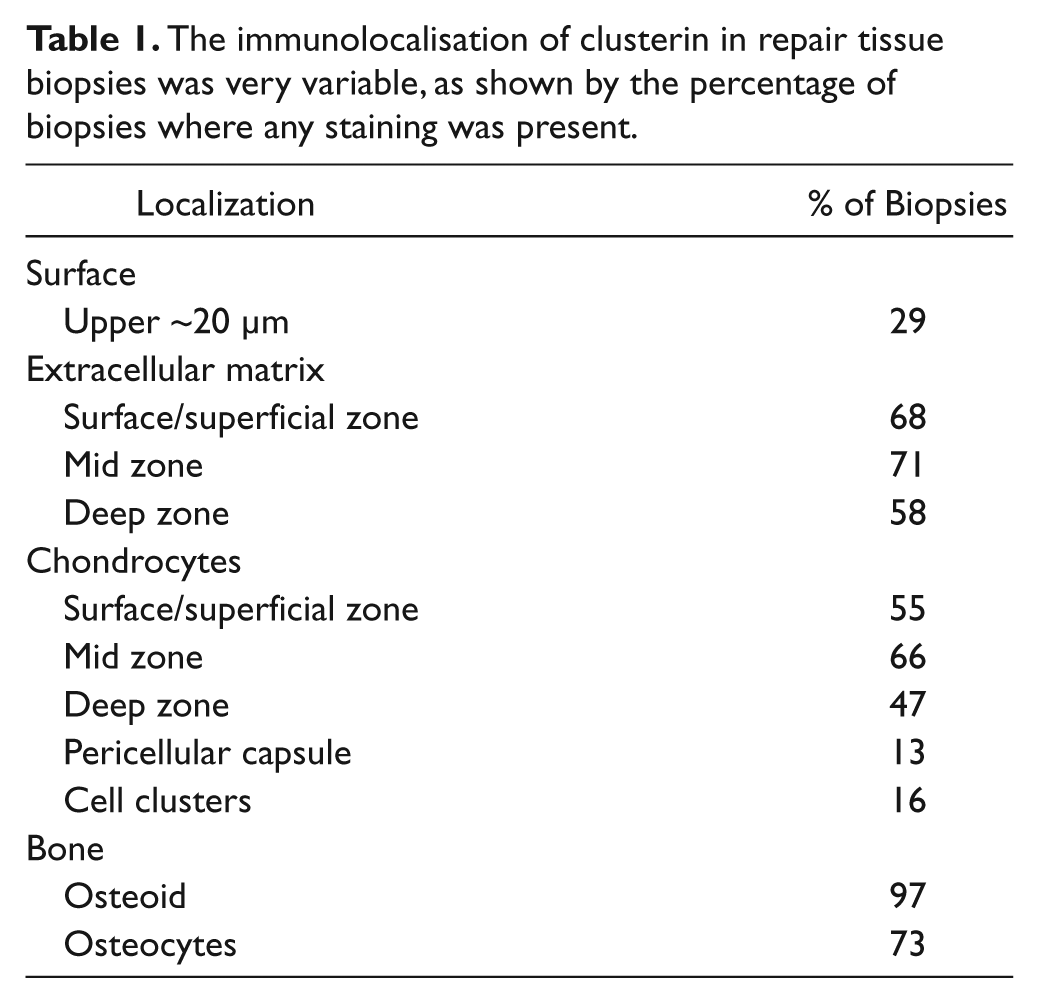
Immunolocalization of clusterin was detected in 37 of the 38 biopsies and was found to be very variable (see Table 1 ). Positive extracellular matrix staining was detected in 31 biopsies, often throughout the full depth of the cartilage ( Fig. 3B ) but usually with the different zones having a different intensity of staining and with considerable variation between samples ( Fig. 3C ). Detection of clusterin in the very upper 5 to 10 µm was observed in only 11 of the ACI biopsies ( Fig. 4A ) and did not correlate with the morphology of the surface when scored with either the ICRS-II or OsScore scoring systems. Immunolocalization for clusterin in the chondrocytes was detected in 32 biopsies, with the majority of positive cells appearing in the mid zone. The localization of extracellular matrix staining did not correlate with the localization of cellular staining, but the extracellular clusterin score and the chondrocyte score were significantly positively correlated (P < 0.001). The presence of positive extracellular matrix staining did not correlate with the morphology of the repair tissue but biopsies consisting of only hyaline cartilage had a significantly higher clusterin chondrocyte score than those containing fibrocartilage (P < 0.01, Fig. 5 ).
The immunolocalisation of clusterin in repair tissue biopsies was very variable, as shown by the percentage of biopsies where any staining was present.
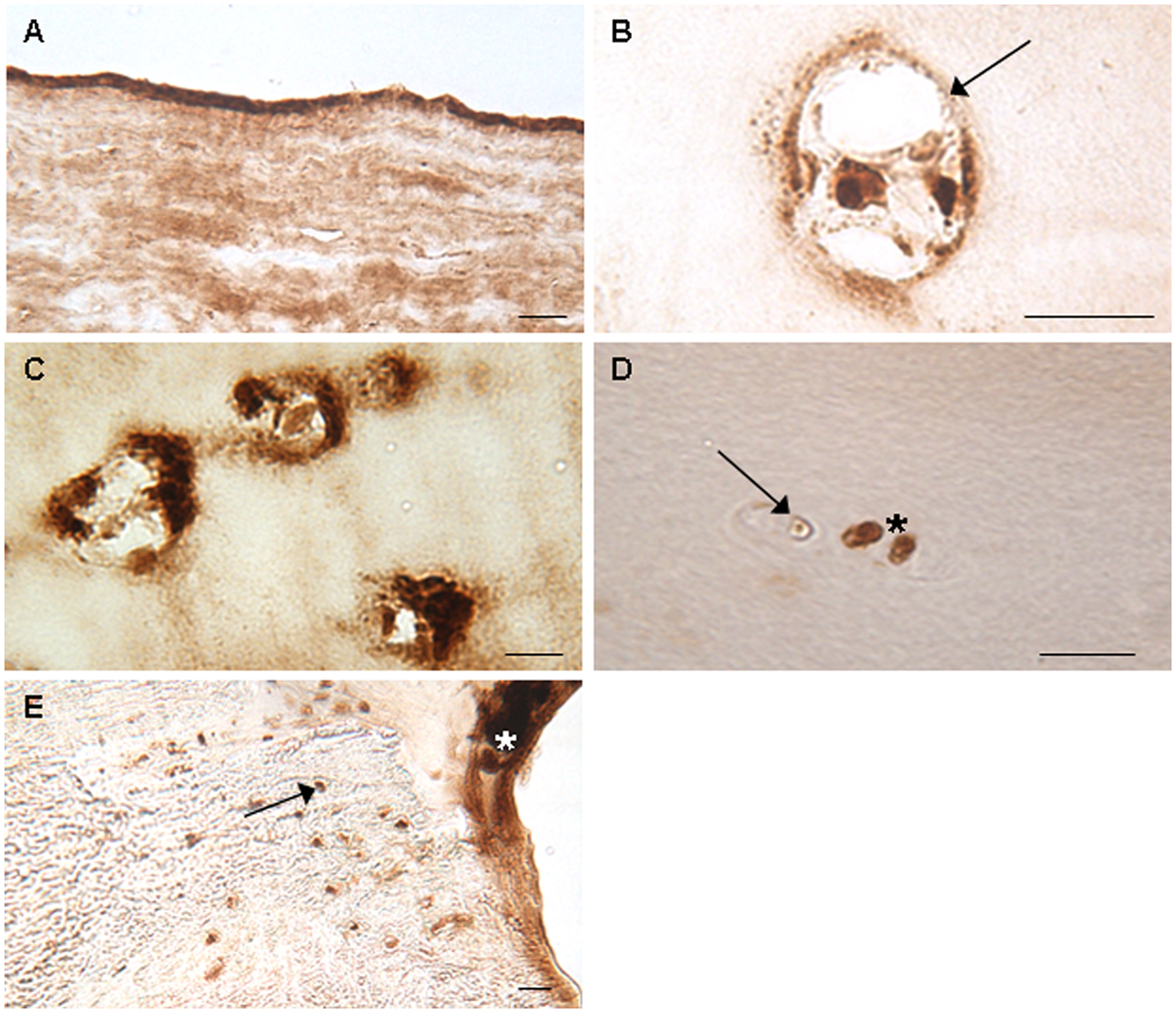
Immunolocalization of clusterin in repair cartilage biopsies in the upper 5- to 10-µm of the surface zone (
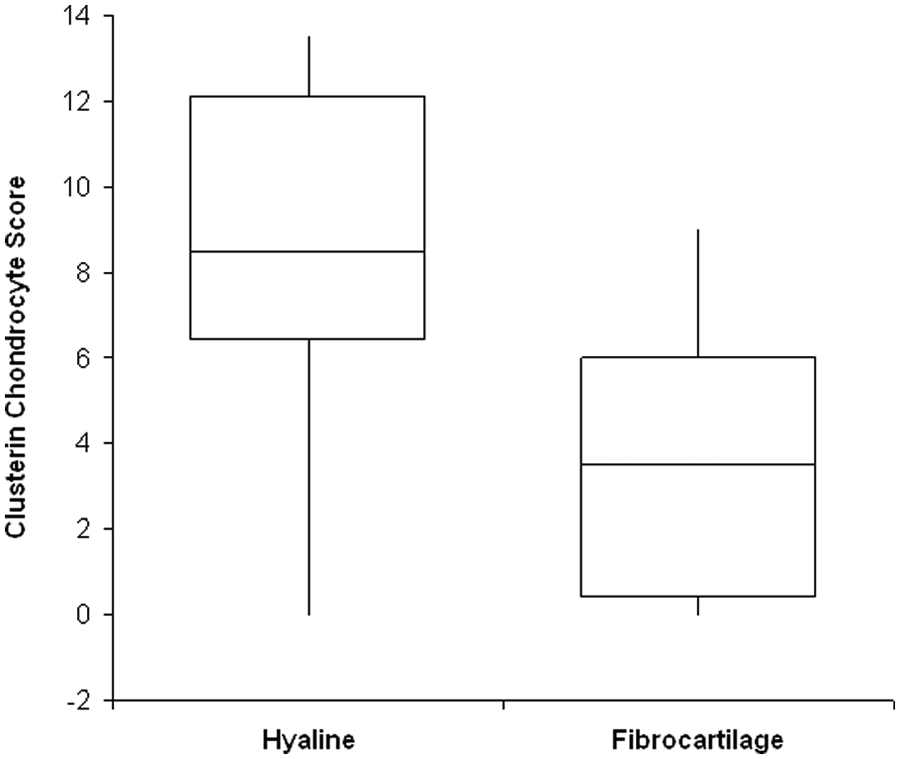
Box and whisker plot comparing clusterin chondrocyte score with tissue morphology. Autologous chondrocyte implantation biopsies consisting of only hyaline cartilage had a significantly higher clusterin chondrocyte score than those biopsies containing fibrocartilage (P < 0.01).
Pericellular capsule staining for clusterin ( Fig. 4B ) was present in only 5 biopsies, also with differing zonal intensity of staining. The presence of this appeared to correlate with tissue morphology with all 5 biopsies where it was seen being of hyaline morphology. The only other biopsy of hyaline morphology within this study exhibited positive immunostaining for clusterin in the bone only.
Of the 11 repair biopsies in which cell clusters were observed, 6 exhibited either moderate or strong positive staining in the clusters ( Fig. 4C ), although not all cells within the clusters would necessarily stain positive for clusterin ( Fig. 4D ). Even within the same biopsy, there was variation in the intensity of staining for clusterin within the clusters. The clustered-cell score for clusterin correlated significantly with the total cartilage clusterin score, the extracellular matrix score, (singular) chondrocyte score, bone score, time since ACI and percentage area of extracellular matrix clusterin staining. Although Ki67 staining was obvious in the positive controls (breast carcinoma) but not in the negative controls, no immunolocalization for Ki67 could be observed in either single or clustered chondrocytes or anywhere else in the biopsies.
Of the 33 biopsies with subchondral bone attached, immunolocalization of clusterin was observed in the osteoid and osteocytes in 32 and 24 biopsies, respectively ( Fig. 4E ). The bone score was found to correlate significantly with the total cartilage score (P < 0.001) but not the extracellular matrix score. The total cartilage clusterin score, the chondrocyte score, and the bone score all significantly positively correlated with time since ACI (P < 0.05) but not age of the patient at ACI or defect size. The extracellular matrix score was not found to correlate significantly with any of the parameters tested. Neither clusterin production nor localization was found to correlate with any other scoring parameter from either the ICRS-II or the OsScore scoring system.
Clusterin production and localization was compared between the repair tissue obtained from the medial femoral condyle (23 biopsies) and the lateral femoral condyle (9 biopsies). There was no significant difference in the clusterin scores for the extracellular matrix, chondrocytes, or bone between the 2 locations. Interestingly, of the 11 biopsies in which immunolocalization for clusterin was observed in the very upper 5 to 10 µm, 10 were taken from the medial femoral condyle and the remaining one from the lateral femoral condyle. Likewise, of the 5 biopsies in which pericellular staining was observed, 2 were from the medial femoral condyle and 3 from the lateral femoral condyle. Also, of the 11 biopsies observed containing chondrocyte clusters, 10 were condyle biopsies. Comparisons for clusterin production and localization in the other treated defect locations could not be analyzed because of low numbers.
Immunolocalization of collagen types I and II were also variable throughout the repair biopsies, ranging from 0% to 100% of positive extracellular matrix staining. Neither collagen type I nor collagen type II exhibited a significant co-distribution with staining for clusterin ( Fig. 6 ), any of the clusterin staining scores, or tissue morphology.
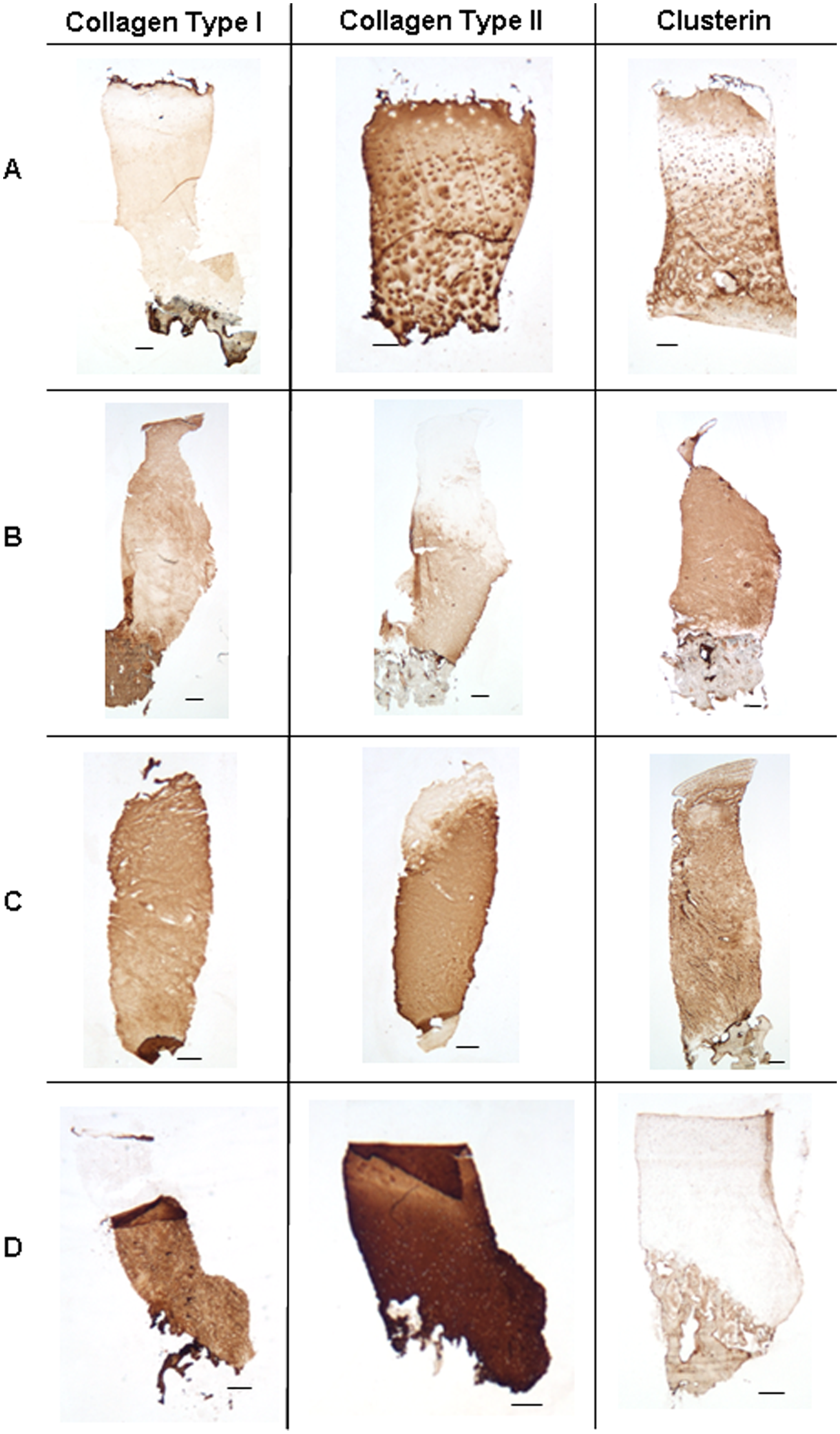
Varying co-distributions between positive extracellular matrix staining for clusterin and collagen types I and II; co-distribution between collagen type II and clusterin (
Clinical Outcome
For those 35 patients having received ACI on the knee, there was an overall significant improvement in Lysholm score from preoperative (mean 43 ± 17, range 17-74) to the time of biopsy (mean 58 ± 22, range 13-95, P < 0.01). However, there was no significant correlation between the Lysholm score at the time of biopsy with the total cartilage clusterin score or any of the OsScore or ICRS-II scoring parameters. For those 3 patients having received ACI on the ankle, 2 patients reported an improved clinical outcome at the time of biopsy whereas for 1 patient the ankle score was reduced.
Discussion
In this study, we have analyzed the localization and distribution of clusterin in the repair tissue of patients having previously received ACI treatment for chondral defects and compared it with healthy cartilage. Although the localization of clusterin in articular cartilage has been demonstrated previously, we are the first to show its localization and distribution in actively repairing human cartilage. We have also demonstrated, by using healthy cartilage obtained from individuals of a wide age range (11-50 years) that the localization and intensity of immunostaining for clusterin in normal cartilage decreases with age. Positive immunostaining for clusterin along the upper 5 to 10 µm on the surface was very prominent in samples from both the 11- and 21-year-olds, with decreasing intensity in the remaining PM samples indicating that clusterin may be required during growth and development of cartilage. Detection of such a band of staining in the ACI biopsies occurred in less than a third of samples, perhaps indicating different degrees of repair and regeneration toward native tissue. A high degree of clusterin has previously been detected in tissue engineered constructs consisting of equine chondrocytes 2 suggesting a possible role for clusterin in the remodeling and regeneration of cartilaginous tissue.
Despite clusterin having been first discovered in sheep testes almost 30 years ago, 17 elucidation of its specific function remains unclear. Clusterin is also known as apolipoprotein J, testosterone-repressed prostate message-2, cytolysis inhibitor, serum protein 40,40, glycoprotein 80, NA1/NA2, clone pADHC-9, and serum glycoprotein 2. 4 This array of different names for clusterin, combined with its almost ubiquitous production throughout the body, could mean either a multitude of different functions for clusterin or disguised commonality of function throughout various tissue types.
Localization of clusterin staining noted at the articulating surface of normal controls and some repair tissue samples could in fact be the lamina splendens, although previous studies have suggested that cell therapies for cartilage defects do not stimulate the repair of such feature.18,19 Once believed to be an artifact, 20 the lamina splendens is a distinct thin acellular layer on the articular surface of intact cartilage in which collagen fibrils run parallel to the surface. 21 It is believed to have key roles in the cartilage’s response to mechanical load, 22 reduction of proteoglycan leakage from the tissue and is often the first region of cartilage to degrade in osteoarthritis (OA). 23 Clusterin localization in the proposed lamina splendens bears a striking similarity to the localization of lubricin, 24 a surface zone proteoglycan associated with surface lubrication of the articular joint. 25 Although clusterin is commonly found at fluid–tissue boundaries as a cytoprotective agent, 26 the age-related decrease in staining of clusterin at the articulating surface of healthy controls observed in this study could be contributory to the development of OA in older individuals.
Although the distribution of clusterin in the superficial zone of the healthy cartilage samples used in this study was similar to previous studies,2,5 we also found immunostaining for clusterin in the chondrocytes and pericellular capsules in the mid zone. In this study, no staining for clusterin was observed in the deep zone of the healthy control samples, in contrast to a previous study where mid zone and deep zone chondrocytes were found to have similar levels of clusterin. 1 Clusterin expression in articular cartilage is indeed increased in early OA and decreased in late OA when compared with healthy controls. 1 In line with this, increased levels of clusterin have also been detected in both the serum and synovial fluid of patients with primary OA. 27 Fandridis et al. 27 therefore suggested that measuring clusterin levels could be used as a biomarker and hence as a diagnostic/prognostic tool in the treatment of OA. However, results presented here demonstrate a lack of correlation between clusterin production, repair tissue histology, and clinical outcome, consistent with previous results. 28
Our study has demonstrated striking differences between the localization of clusterin in actively repairing cartilage compared with normal cartilage, with much higher levels of clusterin in the former. Previous studies have demonstrated an inverse relationship between the severity of OA and the expression of clusterin.1,27 Our results suggest a similar high level of clusterin produced in actively repairing tissue to that previously reported to appear in early OA. Very early stages of OA development involve an initial reparative phase of the tissue. 29 It is possible therefore that the increased levels of clusterin observed early in the disease could be attributed to the tissue’s early attempt at repair, particularly as clusterin is suggested to have a cytoprotective role.4,5 On the other hand, it is of course possible that the increased clusterin observed in early OA is simply indicative of early degenerative change. However, this does not explain the elevated clusterin observed in both repairing cartilage, as observed in this study, or tissue-engineered constructs2,6 in vitro with no signs of degeneration. The role of clusterin in articular cartilage remains unknown, but it is possible that the increase in clusterin in early OA is as a result of the chondrocytes attempting to protect and repair the tissue in the early stages of the disease. 1
Although no significant correlation was observed between the tissue morphology of the repair biopsies and overall staining for clusterin, those biopsies consisting of solely hyaline cartilage had significantly more positive staining in chondrocytes than those biopsies that contained fibrocartilage. This suggests that chondrocyte clusterin production may be contributing to the repair of the cartilage toward a more native phenotype. It would be interesting to obtain further biopsies from such patients to see if the elevated chondrocyte clusterin production reduces with time to become comparable to native healthy tissue. Although the biopsies used in this study ranged widely from the time they were taken postoperatively (between 8 and 67 months), no samples were excluded from the analysis. Even though the total cartilage clusterin score, chondrocyte score, and the bone score were all significantly positively correlated with time since ACI treatment, there are not enough numbers to determine any short- or long-term effects of clusterin. This, however, would be an interesting avenue to investigate in the future to determine how early in the repair process clusterin production is increased, how long the increased production is maintained and if there is a correlation between time and clusterin production. It would also be interesting to compare clusterin production and localization in the repair cartilage formed between ACI and other surgical techniques such as microfracture, mosaicplasty, and matrix-guided ACI.
Biopsies obtained for use in this study came from different locations of treated defects, the majority being from either the medial (23) and lateral (9) femoral condyles. Although the staining patterns of clusterin between all of the different locations were not examined in this study because of insufficient sample numbers, it is something that can be analyzed in the future as more biopsies become available. We were, however, able to compare the staining patterns of clusterin between the medial and femoral condyles. Although there was no significant difference in the clusterin scores between each condyle, clusterin immunolocalization in the upper 5 to 10 µm of the surface of the repair tissue could reflect the mechanical loading specific to this location.
Despite clusterin being found in a variety of different tissues, a common feature is its production in diseased states and tissue injury, particularly those where the tissue is under oxidative stress such as during OA, 30 renal disease, 31 Alzheimer’s disease, 32 diabetes, 33 cancer, 34 and atherosclerosis. 35 The clusterin gene promoter contains both heat shock transcription factor-1 and activator protein-1 response elements 36 thus making the clusterin gene sensitive to changes in reactive oxygen species. 37 It is therefore possible that the increased levels of clusterin observed in OA could have been induced by oxidative stress, perhaps as a defensive mechanism as has previously been suggested. 1
Similarly to clusterin, transforming growth factor-β (TGF-β), a reparative mediator in articular cartilage, is upregulated in early OA 38 and downregulated in late OA. 39 In addition, TGF-β is known to indirectly activate the production of clusterin, 40 so it is possible that TGF-β and clusterin act synergistically in an attempt to repair and maintain the cartilage early in the osteoarthritic degenerative process. Moreover, TGF-β also upregulates the expression of lubricin. 41 Levels of TGF-β were not analyzed in the repair biopsies included in this study. However, it would be interesting to investigate the correlation and potential co-localization between both TGF-β and clusterin production during repair of articular cartilage, considering the link between TGF-β and the similar localization of staining for both clusterin and lubricin.
Although cell clusters are commonly found in osteoarthritic cartilage, they are not considered destructive. 42 The significance of cell clusters in repairing tissue such as those we examined in this study, however, remains unclear. It is possible that they may be formed as part of the regenerative remodeling healing process as they are observed in immature and developing cartilage in both humans and animals43,44 even though their formation alone is believed to be insufficient for repair. 45 Many clusters observed in the patients in this study stained positively for clusterin, although some clusters did not show positive staining for clusterin even within the same biopsy. Clusterin is known to play a role in cell proliferation 46 and chondrocyte clusters have been reported to form via chondrocyte proliferation. 47 Many chondrocyte clusters observed in the repair tissue biopsies included in this study were found to be positive for clusterin. Despite this, all biopsies were in fact negative for Ki67, a nuclear antigen expressed in proliferative cells. This is in agreement with a previous study where there was no correlation observed between clusterin and Ki67 in tissue-engineered constructs. 2 Malda et al. 2 hypothesized that clusterin was therefore produced in the latter stages of cluster development as the cells reached a less proliferative and more terminally differentiated phenotype. It is possible that the same applies within these biopsies.
The results presented here also demonstrate a range of staining distributions for both collagen types I and II with no correlation to either clusterin staining or the morphology of the repair tissue. Collagen type I is generally either absent from healthy articular hyaline cartilage or is present in very small amounts whereas it is present in large amounts in fibrocartilage. 48 In contrast, collagen type II is the main collagen component in healthy articular hyaline cartilage.48,49 A previous study has demonstrated, however, that the presence of either collagen type I or II is not indicative of the quality or morphology of the repair tissue formed. 50 The various distributions of staining for collagen types I and II throughout the tissue and the lack of correlation to either clusterin staining or repair tissue morphology, may indicate different roles for clusterin and collagens in the repair tissue. In the same way that collagens and proteoglycan content are used to assess cartilage formation, clusterin may also serve as a useful addition to clarify the nature of the repair tissue.
Taken together, the results of the present study demonstrate a very different distribution of clusterin in repairing cartilage from that observed in healthy adult control cartilage. Although the pattern was very variable in the repair tissues, in some cases it resembled that seen in the youngest control samples studied. This could suggest that the repair cartilage is perhaps in a state of remodeling akin to that of developing cartilage. 49 This variability perhaps illustrates the patient-to-patient variation, but it could also indicate that the cells and the repair tissue are in different states of dedifferentiation and redifferentiation toward a true adult chondrogenic phenotype. Additionally, it has been demonstrated that zone-dependant expression of clusterin varies with culture duration and oxygen tension, 51 and care should be taken when evaluating so-called zone-specific markers in tissue-engineered cartilage. Different isoforms of clusterin have been found in other disease states such as Alzheimer’s; if alternative isoforms are found at a later date to occur in OA it may be possible to differentiate between these isoforms and so distinguish between the clusterin produced in healthy versus diseased tissue.
Currently, the clinical significance of clusterin within repair tissue is difficult to assess, but its presence is unlikely to have detrimental effect. Indeed, it is to be expected that the ideal functioning repair tissue should ultimately have a morphology resembling that of healthy adult cartilage. It was anticipated that the temporospatial production of a protein such as clusterin in repair tissue forming post-ACI treatment would provide information on the biological processes involved. However, as collagen production in repair tissue has previously been demonstrated to not be indicative of its quality or morphology, 49 it is possible that other markers, such as clusterin, may also have expression levels different to what would be expected in native tissue. Hence, we need to further our understanding of the processes involved during cartilage repair and how to drive it toward regenerating true articular cartilage. It may be that clusterin is a significant factor in this process.
Footnotes
Authors’ Note
The work reported in this article was undertaken at the Robert Jones and Agnes Hunt Orthopaedic Hospital NHS Trust.
Acknowledgments and Funding
We are grateful to the Arthritis Research UK for supporting this work (Grants 19429 and 18480), to the Dutch Arthritis Foundation for supporting JM, to Dr. B.F. Murphy (Austin Health, Melbourne, Australia) for providing the G7 clusterin antibody and to The John Charnley Laboratory, RJAH Orthopaedic Hospital, UK, for the expansion and preparation of harvested chondrocytes used in the ACI procedures. The anti-collagen type II antibody (clone CIIC1) developed by Rikard Holmdahl and Kristofer Rubin was obtained from the Developmental Studies Hybridoma Bank, developed under the auspices of the NICHD and maintained by The University of Iowa, Department of Biology, Iowa City, IA 52242.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical Approval
This study was approved by our institutional review boards.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.