
Abstract
The methyl-CpG binding proteins (MBPs) interpret the methylation of DNA and its components. The number of MBPs in the human body currently stands at 15, which are split into 3 branches, a reflection of the intricate mechanisms of gene regulation. Each branch utilizes a different mechanism for interacting with methylated DNA or its components. These interactions function to direct gene expression and maintain or alter DNA architecture. It is these functions that are commonly exploited in human disease. For this review, we will focus on each protein and any roles it may have in initiating, promoting, progressing, or inhibiting cancer. This will highlight common threads in the roles of these proteins, which will allow us to speculate on potentially productive directions for future research.
Keywords
Introduction
The field of epigenetics and cancer is broad and extensive. The extent of these studies is a reflection of how much eukaryotic biology is reliant on appropriate DNA methylation. A key aspect of the field involves the study of the proteins that directly interpret the methylation of DNA and its components. The methyl binding protein (MBP) family plays a pivotal role in this interpretation. Since the identification of the first MBP in 1989, MeCP2, 1 their numbers have increased, and their roles in gene regulation have been extensively studied as incorrect interpretation of methylation, or vice versa, the interpretation of incorrect methylation, plays a role in many genetic diseases. For this review, we will focus on the roles of the MBPs in cancer. Initially, we will provide a brief history of the MBPs and a summary of the individual proteins. We will then provide an update on the most recent cancer-related findings for the MBPs since previous reviews on this topic in 2008.2,3 Finally, we will speculate on future directions for understanding the roles of MBPs in inhibiting, initiating, progressing, and altering cancer phenotypes.
A Brief History of MBPs
In mammalian DNA, the MBPs were originally characterized by their interactions with methylated DNA, namely CpG dinucleotides. The CpG is a cytosine-guanosine (CG) dinucleotide DNA sequence, in which the cytosine undergoes chemical modification to contain a methyl group. This methylation of cytosine was first detected in 1925 in the tubercle bacillus. 4 It was another 23 years before it was observed in eukaryotes when it was detected in calf thymus DNA in 1948. 5 The importance of methylation became apparent in the 1970s following key findings such as the fact that the doublet CpG sequence occurs at approximately one fifth of the expected frequency within the genome6,7 but represents the main target of methylation.8,9 These findings suggested that the regulation of genes had to be due to the interaction of genomic DNA with the cytoplasmic enzymes that it encoded.10,11 By the end of the 1970s, it had been demonstrated that an important aspect of eukaryote gene regulation came from protein-methylated DNA interactions12,13 and that DNA methylation patterns are inheritable.14,15
Today, the nature of DNA methylation and many of its roles within eukaryotic cells have been elucidated. The enzymes that catalyze the methylation of a CpG dinucleotide have been identified and characterized into the DNA methyltransferase family. 16 These enzymes take a methyl group, donated from S-adenosyl-L-methionine, and cova-lently attach it to the fifth carbon position of cytosine to form a symmetrical CpG dinucleotide. In the mammalian genome, approximately 60% to 90% of CpGs are methylated. 17 The remaining unmethylated CpGs are clustered into what are termed CpG islands, which comprise around 1% of the genome.18,19 These CpG islands are located within the promoter, first exon, or 3′UTR regions of approximately 60% of the RNA polymerase II–transcribed genes in the vertebrate genome, consistent with their role in gene expression.16,20 Indeed, methylated CpGs (mCpGs) are now synonymous with a repressed chromatin environment and silencing of gene expression. They play an important role in expression of patterns of imprinted genes, X chromosome inactivation, suppression of transposable elements, developmental biology, and many disease mechanisms. 17
One of the key aspects of the relationship between methylation of DNA and the silencing of gene expression was established in the late 1980s and early 1990s, with the identification of proteins that could bind to mCpGs and recruit protein complexes. These complexes contained histone-modifying enzymes, which leads to heterochromatin formation and gene silencing. 21 The initial identification of a methyl-CpG binding protein1,22 (MeCP1), which was capable of binding at least 12 symmetrically methylated CpGs, was later shown to be a complex with other MBPs. It was the subsequent identification of the first single protein capable of binding a single methylated CpG, the 53-kDa protein MeCP2, 1 that paved the way for greater understanding of the role of methylation and gene regulation. In MeCP2, a core 70–amino acid methyl binding domain (MBD)23-25 was identified along with a transcriptional repressor domain (TRD).24,26 This MBD was subsequently used to identify other proteins, which initially made up the MBP family. However, the last decade has seen the addition of 2 further branches following the identification of proteins that utilize different structures to recognize and bind methylated DNA or its components. In 2001, the protein Kaiso defined a second family that uses 3 tandem zinc fingers for binding mCpGs. 27 The third family of MBPs was identified after the UHRF1 and UHRF228,29 proteins were shown to bind mCpG using their SET and RING finger–associated domain (SRA).
The MBP Superfamily and Cancer
As previously mentioned, the MBP family is divided into 3 branches: 1) MBD containing proteins, 2) methyl-CpG binding zinc fingers, and 3) the SRA domain containing proteins. 30 This section will cover the functions of the individual members of each family that are present in humans and highlight any roles in cancer (Table 1). Much of the data available on the roles of these proteins in cancer are related to their functions as transcriptional repressors or chromatin remodelers (Fig. 1). There is little evidence in the literature that deficiency of any of these proteins results in elevated cancer predisposition, a notion supported by a lack of any entries in the Catalogue of Somatic Mutations in Cancer (COSMIC) database 31 implicating their inactivation.
Types of Cancer in Which Methyl-CpG Binding Proteins (MBPs) Have Been Implicated to Play a Role

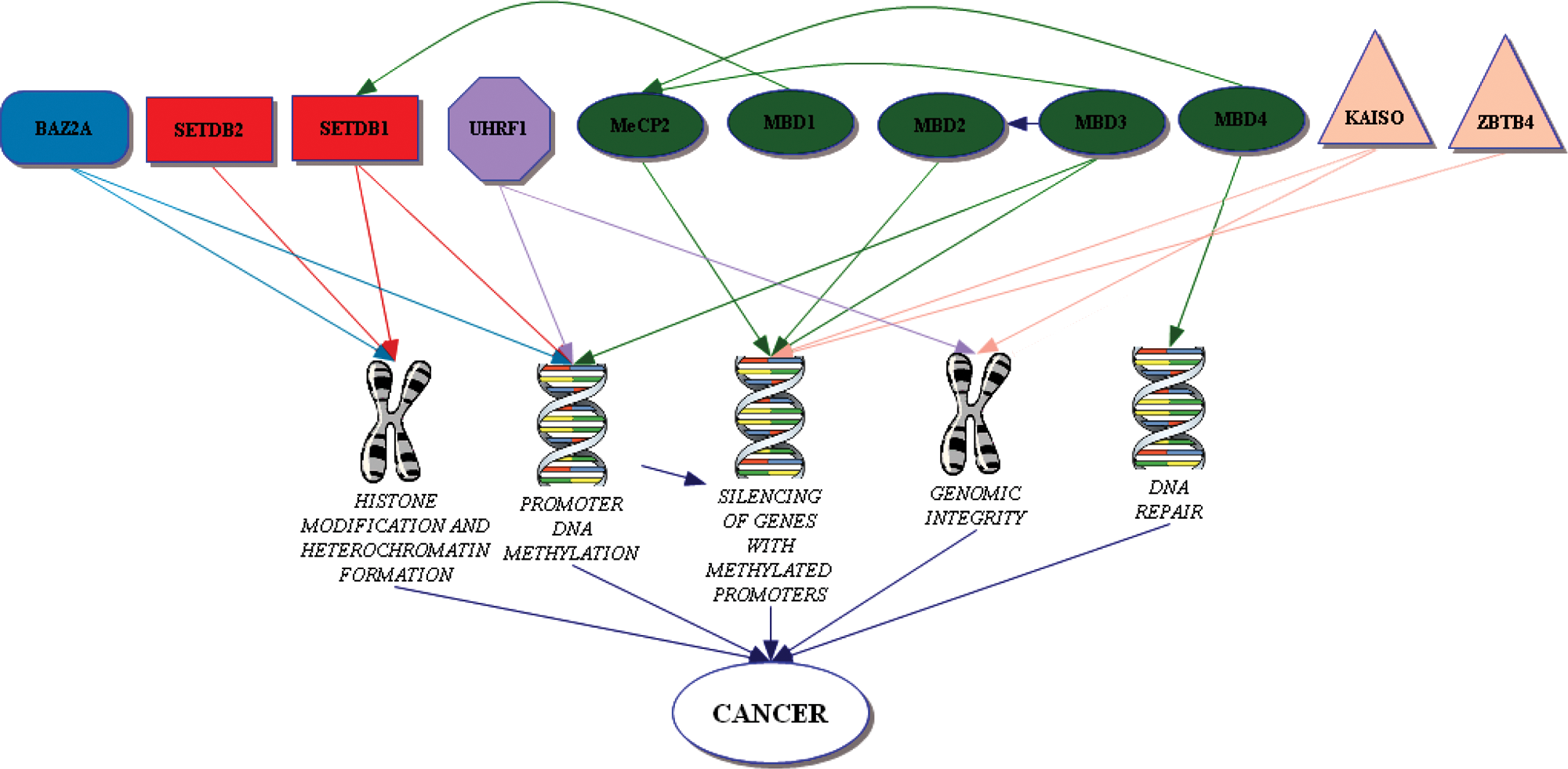
Mechanisms by which methyl-CpG binding proteins (MBPs) are implicated in cancer.
MBD-Containing Proteins
These proteins comprise the largest branch of the MBP family. Currently, the NCBI Conserved Domain Database (CDD)32-34 lists 11 human proteins containing the MBD derived from MeCP2 (CDD: cl00110 and cd00122). Based on the presence of other domains, these are further divided into 3 groups within the MBD superfamily according to the CDD 30 : the histone methyltransferases (HMT_MBD; CDD reference cd01395), the MeCP2_MBD proteins (CDD reference cd01396), and the histone acetyltransferases (HAT_MBD; CDD reference cd01397). Although all the members of this group contain the MBD, they do not all directly interact with methylated CpGs.
HMT_MBD
There are 2 members of this family: SETDB1 (also known as ESET) on chromosome 1 and SETDB2 (also known as CLLD8) on chromosome 13. As well as containing a MBD, they carry a PreSET and bifurcated SET domain, which mediates protein-protein interactions. SETDB1 was identified due to its interaction with the KAP-1 corepressor. 35 SETDB2 was identified due to its role as a potential gene involved in leukemogenesis. 36 They are both protein lysine methyltransferase enzymes that repress transcription through the formation of heterochromatin.35,37,38
As may be expected from their function, their roles in cancer are defined by the genes that they repress and interact with. Currently, the only evidence implicating SETDB2 in neoplasia is that it is 1 of the 14 genes encompassed by a 1-Mb deletion on chromosome 13q that is associated with disease progression in chronic lymphocytic leukemia. 39 There is increased evidence for a tumorigenesis role for SETDB1 as it has been shown to interact and mediate the functions of proteins, which have been demonstrated to have roles in cancer, for example, MCAF1, 40 KAP1, 38 serine/threonine kinase AKT, 41 and DNMT3A. 42
The SETDB1 interactions with MCAF1 and DNMT3A link promoter CpG hypermethylation to histone methylation, a crucial precursory event to heterochromatin formation. Its interaction with MCAF1 also links it to another transcriptional repressor, MBD1 (another MBP). 43 Through MBD1, it is directed to mCpGs, where the MBD1:SETDB1: MCAF1 complex converts the dimethyl H3-K9 to trimethyl H3-K9, resulting in heterochromatin formation and transcriptional repression.40,44-46 In cancer, this MCAF1:MBD1 complex is associated with the maintenance of telomerase activity, 47 but this function seems independent of SETDB1. The interaction with the DNMT3A protein is relevant as it is a de novo DNA methylase, which plays crucial roles in cancer pathogenesis via methylation of CpG islands.48,49 DNMT3A can also mediate gene repression independently of CpG methylation via heterochromatin formation.50,51 The SETDB1/DNMT3A interaction has been shown to mediate silencing of the p53BP2 and the RASSF1A gene in ovarian and breast cancer cell lines, respectively. 42 These genes have roles in apoptosis, DNA repair, and cell cycle arrest and are silenced in a variety of cancer types due to hypermethylation of their promoters.52-56
The link with heterochromatin formation is further supported by its interaction with KAP1, which also provides a link to global gene regulation as KAP1 is a universal cofactor involved in regulating the Kruppel-associated box domain zinc finger proteins (KRAB-ZFP), the largest group of transcriptional repressors in higher organisms. 57 Surprisingly, for such a large group, there is a paucity of information on the targets of KRAB-ZFP proteins. Along with the ability of KAP1 to mediate long-range repression through heterochromatin spreading, it is difficult to assess any specific roles for its interaction with SETDB1 in cancer. 38
It is perhaps SETDB1 interaction with Akt that is currently of greatest significance as the aberrant activation of the Akt/PI3K pathway is being targeted for treatment in a variety of cancers. 58 The role of SETDB1 on Akt, the main downstream effector molecule of the PI3K pathway, 59 is not well understood, although it has been demonstrated in vitro to enhance the ability of Akt to repress the FOXO3 transcription factor. 41 The FOXO transcription factors are increasingly considered to represent unique cellular targets directed against human cancer. As they have proapoptotic effects, they have an ability to lead to cell cycle arrest 60 and interact with β-catenin, the key effector molecule of the Wnt pathway, which is commonly deregulated in colorectal and many other cancers. 61
MeCP2-MBD
This represents the largest group of the MBD branch of the MBP superfamily. It comprises the original MeCP2 protein and 6 others termed MBD1 to MBD6.19,62,63 The roles of many of these proteins in cancer were reviewed in 2007 2 and 2008. 3 We will provide an update of the latest developments for the regulation of these proteins that have, or may have, roles in cancer.
MeCP2
The founder protein of the MBP superfamily MeCP2 is a global transcriptional repressor. It is a 50-kDa protein encoded by a gene on the X chromosome. It can bind to a single methylated CpG and recruit the Sin3A repressor complex to silence transcription via histone deacetylation. 26 As well as its role in Rett syndrome, a neurodevelopmental disorder, 64 it has also been shown to have a role in tumorigenesis2,3 as many cancers rely on genes becoming silenced via hypermethylation of their promoter. Recently, it has been shown to target genes in vivo in myeloma, 65 hematological malignancies, 66 and breast,67,68 colorectal, 69 lung, 70 liver, 71 and prostate 72 cancer. Despite its relevance to cancer, we still have little understanding of how the protein itself is regulated. However, due to the large amount of work on MeCP2 in the nervous system, we are now beginning to unravel these processes. These processes highlight potential avenues for exploration in cancer and therapy. For example, an ability to control the expression of the MeCP2 and HDAC1 to HDAC3 genes in C6 glioma cells using valproic acid, a widely used anticonvulsant and mood stabilizer, has recently been demonstrated. 73 Similarly, the miRNA miR-212 has been shown to suppress the translation of MeCP2 RNA in vitro in gastric carcinoma cells. 74 Significantly, in the 11 primary gastric carcinomas analyzed in this study, those with the smallest depth of invasion showed the highest expression level of the miRNA and the lowest levels of MeCP2. Finally, at a protein level, the first kinase to phosphorylate MeCP2 has been identified. Homeodomain-interacting protein kinase 2 (HIPK2) binds to MeCP2 in HEK293T and human fibroblast cells to phosphorylate the serine residue at position 80. 75 This phosphorylation event increases the apoptotic activities of MeCP2 via a mechanism that requires the MBD. As we understand further these regulatory processes, our ability to manipulate and target MeCP2 should lead to targets for therapy in cancer and Rett syndrome.
MBD1
The MBD1 protein is the largest member of the family. It has a complex expression profile as there are 13 isoforms of the gene expressed from chromosome 18. The main difference between the isoforms is the presence of 2 or 3 CXXC-type zinc fingers present in the protein. 76 The isoforms containing the first 2 CXXC domains preferentially repress methylated promoters, whereas those with the third CXXC domain are capable of DNA binding regardless of methylation status. 77 These isoforms containing the third zinc finger have been shown to repress the transcription of genes with unmethylated promoters in vitro. However, this ability to bind unmethylated DNA in humans seems to be largely dispensable.78,79 It is unlikely that these isoforms have a role in the mechanism of transcriptional repression as this is mediated via an interaction with proteins from the MCAF/AM family via the transcriptional repression domain.40,44 These MCAF/AM proteins also interact with another MBD-containing protein, SETDB1, to form the MBD1:MCAF1: SETDB1 complex, which we discussed previously.
In relation to cancer, polymorphisms in MBD1 have been shown to be significantly associated with lung cancer risk in a Chinese population, 80 both positively and negatively. The effect of these SNPs on the protein or its expression has yet to be investigated. In relation to its transcriptional roles, most of the genes silenced by MBD1 in cancer have been identified in vitro. These targets have been identified in acute promyelocytic leukemia 81 and pancreatic, 82 prostate, 83 and colon 84 cancer cell lines. MBD1 targets may also have a role in drug resistance and the immune system interactions of cancers as the MBD1 gene is present on an amplified region of chromosome 18, which is associated with the development of vinblastine resistance in human cervical carcinoma KB cells. 85 From an immune perspective, MBD1 silences the IRF8 gene in colon carcinoma cells, which is a central mediator in the IFNγ/STAT1 signaling pathway. 84 This silencing frequently occurs in human colorectal carcinoma in vivo, and IRF8 expression has been shown to induce apoptosis in tumor cells. 86 Given that the IFNγ/STAT1 pathway has a critical role in governing the establishment of innate immune responses, 87 disrupting it through such MBD1-mediated silencing clearly has implications for immune surveillance.
The capability of these MBD1 isoforms to play a role in cancer is likely to be effected by factors such as redundancy and target specificity. As MBD1 binds with greater efficiency to genes in which the promoter has the sequence TCMGCA or TGCMGCA, 78 and a comparison of cancer cell lines of different origins demonstrated that the promoters of many silenced genes were occupied by more than one MBD protein. 79
MBD2
The gene for MBD2 is approximately 4 Mb downstream of MBD1 on the “q” arm of chromosome 18. They have limited similarity to each other, whereas MBD2 shows 71.1% overall amino acid identity to MBD3. 62 MBD2 mediates the methylated DNA binding functions for 2 different transcriptional repressor complexes, MECP1 88 and Mi2/NuRD.89,90 Both these complexes use MBD2 to direct HDACs and chromatin remodelers to methylated promoters, where they effect transcriptional repression. Once again, this protein has been shown to silence genes in a variety of cancers: colorectal, 91 lung, 92 prostate,93,94 and renal. 95 The relevance of the silencing of these genes in cancer is apparent in colorectal cancer as mice deficient for MBD2 develop relatively normally, although they do show impaired maternal behavior and exhibit resistance to Wnt-driven cancer. 96 Hyperactivity of the Wnt pathway is responsible for the majority of colorectal cancer cases. These Mbd2-null mice also show an increased ability to retain a degree of control over the Wnt pathway when it is constitutively activated. 97 As the deficiency of MBD2 in mice appears relatively benign, it makes an attractive therapeutic target. Work is underway to further identify the derepressed genes in these mice, which results in the manifestation of this phenotype. One identified in vivo target for MBD2 silencing is IL-4, which plays a role in the immune system and IFNγ regulation.98,99 The deregulation of IL-4 upregulates IFNγ due to a negative feedback loop, 100 giving an increased Th1 immune response. 101 As IFNγ has been shown to subdue Wnt targets 102 and a Th1 response is a key element in tumor immune surveillance, 103 this clearly provides an attractive potential mechanism of perturbing tumorigenesis.
Despite its attractiveness as a therapeutic target in colorectal cancer, no drugs that specifically target the MBD2 gene or protein have been identified to date. However, the drug B1 (N-[2-[dimethylamino]ethy]l-2-aminothiazonaphthalimide) has been shown to alleviate MBD2 repression of the 14-3-3σ gene in promyelocytic leukemia cells and induce their apoptosis. Further characterization of this ability would be desirable. An alternative way of targeting MBD2 is offered by targeting the PRMT1 or PRMT5 protein arginine methyltransferases that have been shown to methylate the MBD2 arginine residues. This methylation inhibits the ability of MBD2 to associate with its repressive complexes and silence genes. 104
As attractive as MBD2 is as a therapeutic target, caution must be exercised in an application that will deregulate global gene transcription. By way of illustration, MBD2 is known to play a role in the silencing of the human telomerase reverse transcriptase promoter (hTERT) in somatic cells. Given that upregulation of this gene is a key event for the ability of many cancer cells to overcome replicative senescence associated with telomere shortening, 105 there are clear potential negative ramifications for inactivation of MBD2.
MBD3
The chromosome 19 gene, MBD3, has multiple spice variants, all of which are unable to bind methylated CpGs due to 2 alterations within the MBD. 106 Despite this, it still plays a crucial role in the silencing of methylated genes. It is an important part of the Mi2/NuRD repression complex, which can use MBD2 as its means of methylated DNA binding. The cancer-related genes repressed by this complex were discussed in the previous MBD2 section. In the absence of MBD2, the repression complex, hereafter termed MBD3/NuRD, uses other proteins for DNA binding. These non-MBD2–related roles are important in development, as Mbd3–/– mice die in utero, but it is its non-MBD2–related roles in cancer we will discuss here.
Roles in human cancer that are specific to MBD3 are not commonly observed. 107 There are MBD2-independent reports of a role for the MBD3/NuRD complex shown in both prostate cancer cells 108 and leukemias. In acute promyelocytic leukemia, it interacts with the fusion PML-RARa oncoprotein that is generated from the 15;17 translocation, 109 and in acute myeloid leukemia, it interacts with the Evi1 gene to inhibit histone deacetylation and activate genes. 110 In human lung cancer cells, MBD3/NuRD regulates p21WAF1/Cip1 and ErbB2 111 expression levels, and these can be altered by treatment with the HDAC inhibitor (HDACi) trichostatin A. Intriguingly, the HDACi released MBD3 from the p21WAF1/Cip1 promoter in cancer cells but recruited it in normal cells and vice versa with the ErbB2 promoter. This may reflect the fact that MBD3 can bind unmethylated DNA and mediate the observed histone acetylation even in the presence of HDACi.106,112,113 The p21WAF1/Cip1 observation is corroborated in HeLa cells, where depletion of MBD3 enhanced its transcription but repressed cyclin B1, Plk1, and survivin. 114 This resulted in an arrest at G2/M that led to cell death due to abnormal mitosis.
A different role has been shown in mouse lymphosarcoma cells in which Mbd3 interacts with Brg1 and Dnmt3a 115 to silence the methylated MT-I gene. It remains to be identified which complex mediates the silencing as the association of Mbd3/NuRD repressor with Dnmt3a can link it to methylated DNA. Alternatively, the SWI/SNF complex, of which Brg1 is a central component, could also be used as it associates with the Mecp2/Sin3a/HDAC complex.116,117 Its roles in the SWI/SNF complex are not well understood but may make MBD3 relevant to many more cancers than previously suspected. 118
The successful development of drugs to target this protein will again be dependent on tumor-specific targeting as MBD3 has recently been shown to play a significant role in maintaining intestinal homeostasis through the interaction with nonphosphorylated c-Jun.119,120 The N-terminal phosphorylation of c-Jun inhibits this interaction and results in upregulation of its target genes in intestinal homeostasis and tumorigenesis. In the absence of MBD3, the intestines are characterized by hyperproliferation and an increased susceptibility to colitis-induced tumorigenesis. Nevertheless, therapeutically targeting MBD3 may prove to have benefits in cancer treatment. Potentially, we could develop drugs that are specific for the role in silencing methylated genes. The proteins of 2 other genes, MBD3L1 and MBD3L2, which show substantial homology to MBD2 and MBD3 but lack a MBD, 121 may give us a clue to how to approach this as MBD3L2 can interact with the MBD2/MBD3/Mi2/NuRD complex in a way that displaces the complex from methylated DNA and reactivate transcription. 122
MBD4
MBD4 is primarily encoded from a single transcript on chromosome 3 62 and stands out from the other MBD proteins due to its interactions with the DNA repair machinery. 123 The protein uses its N-terminal MBD to bind methylated CpG and its C-terminal glycosylase (which is homologous to base excision repair proteins) to mediate repair of the hypermutable CpG, which is susceptible to mismatch formation following either the hydrolytic deamination of 5-methylcytosine to thymine or deamination of cytosine to uracil. Thus, MBD4 plays a key role in maintaining methylated DNA gene regulation and suppressing mutation at CpG sites.
These abilities, along with its interaction with the mismatch repair protein MLH1 (mutL homolog 1), potentially impact chemotherapy as MBD4 has been shown to be required for the maximal apoptotic effects of many of these chemotherapeutic drugs,124,125 a role that may in part be mediated through an interaction with Fas-associated death domain protein (FADD). 126 This indicates that MBD4 function is crucial for normal mutation suppression and optimal response to chemotherapy; however, there is also evidence that its absence can be beneficial, as it has been shown in vitro that the radiosensitizing drug iododeoxyuridine is more effective in the absence of MBD4 due to the normal activity of MBD4 in repairing G:IU mismatches located in CpG dinucleotides.
The significance of MBD4 in cancer is largely indirect as deficiency results in a mutator phenotype and subsequent alterations to bona fide tumor suppressor genes, evidenced by increased susceptibility conferred by MBD4 deficiency to intestinal tumorigenesis in the ApcMin/+ model. 127 In this model, loss of function of MBD4 increases the number of “second hit” mutations at CpG sites in the wild-type Apc allele. This observation is echoed in man in whom an increased mutation rate at CpGs confers susceptibility to colorectal cancer,128,129 stomach cancer,130,131 and progression of liver cancer. 132 Consistent with a role in mutation suppression, early downregulation of the gene has been observed in precancerous tissues, implicating it in tumor initiation rather than progression. 133 Whether this loss is required for the onset of genomic instability due to repair defects or suppression of apoptosis is unclear.
Although it is best known for its repair activities, MBD4 has also been shown to repress p16INK4a and MLH1 genes in a methylation-dependent fashion 134 in vitro, as it associates with the Sin3A repressor complex, which MeCP2 also interacts with. Its presence at the promoters of these genes is proposed to maintain the integrity of the CpG methylation signal. However, this repression (along with repression by the MBD2/Mi-2/NuRD complex) is enhanced by an interaction with the RET finger protein (RFP; also known as TRIM27). 135 This interaction with RET potentially links MBD4 to a much more extensive range of cancers. 136
MBD5 and MBD6
MBD5 on chromosome 2 and MBD6 on chromosome 6 were initially 2 uncharacterized proteins that were renamed in 2003 due to them containing a MBD domain.19,63 There is a limited amount of data available on the functions of these proteins. Neither of them is likely to bind methylated DNA, but they do associate with heterochromatin, where they may contribute to the formation or function of heterochromatin. 137 As with other MBP proteins, MBD5 deficiency is linked to a developmental disorder, the 2q23.1 microdeletion syndrome,138,139 but there have been no reports linking either of them to cancer.
HAT_MBD
BAZ2A and BAZ2B
There are 2 proteins in this final group: BAZ2A (also known as TIP5), encoded by the BAZ2A gene on chromosome 12, and BAZ2B, encoded by BAZ2B on chromosome 2. These proteins were identified by searching the EST and cDNA libraries for genes that encoded bromodomains. 140 Bromodomain proteins have been identified as integral components of chromatin remodeling complexes and frequently possess histone acetyltransferase activity. They both contain a MBD, DDT, PHD, and bromodomain consecutively arranged in the same order. 30 The MBD domain in these proteins differs at specific residues from other superfamily members, and it is unable to bind to a methylated oligonucleotide, which would be recognized by MBD2. 141
The main identified role of these proteins is in the epigenetic silencing of ribosomal DNA (rRNA) within the nucleolus.141-144 The BAZ2A protein is a major component of the nucleolar remodeling complex (NoRC) that represses ribosomal gene transcription via histone deacetylation. This NoRC complex associates with the histone deacetylase HDAC1 in vivo and in vitro via the bromodomain of BAZ2A. It appears that NoRC silences the rDNA locus by targeting the SIN3 corepressor complex to the rDNA promoter, which alters the local chromatin structure and leads to methylation of critical CpG residues. 145
There are very few reports of these proteins having a role in cancer. It has been shown that overexpression of miRNAs, which can regulate expression of BAZ2A, correlates with progression to metastasis in prostate cancer 146 and may also have a role in chronic lymphocytic leukemia. 147 However, it currently remains unclear if these proteins will have a significant role in cancer.
Methyl-CpG Binding Zinc Fingers
This branch of MBPs is based on a 3–zinc finger motif found in the C-terminus of the Kaiso protein, which is capable of binding a pair of methylated CpG dinucleotides. 27 The protein also binds unmethylated DNA and has a greater affinity for CTGCNA than mCGmCG. 148 The Kaiso protein is the archetypal member of this branch, and the zinc finger motif has been used to identify 2 other members, ZBTB4 and ZBTB38, 149 that are also capable of binding methylated DNA.
Kaiso
Kaiso is encoded by the ZBTB33 gene on the X chromosome. It belongs to the zinc finger and broad-complex, tramtrack, and bric-a-brac/poxvirus and zinc finger (BTB/POZ) protein family. Kaiso was first identified as a binding partner of p120ctn in a yeast 2-hybrid screen. 27 This p120ctn protein associates with Kaiso at the cell surface (as part of the E-cadherin-catenin cell adhesion complex) and in the nucleus.27,148,150 At the cell surface, it regulates the expression and stability of cadherins and thereby influences their roles in adhesion and signaling.151,152 In the nucleus, it has been shown to function as a transcriptional repressor by directing the nuclear corepressor complex N-CoR 153 to methylated and unmethylated gene promoters. N-CoR is a complex that contains histone deacetylase 3,154,155 which operates to generate a transcriptionally inactive chromatin structure.
Given its multiple functions, it is perhaps not surprising that altered Kaiso activity is a feature of some cancers, particularly given that Kaiso functions as a regulator of the target genes of the canonical and noncanonical Wnt pathways.150,156-158 In Wnt-driven colon cancer cell lines, Kaiso mediates the silencing of the tumor suppressor genes CDKN1A, HIC1, 159 and Rb. 160 Its relevance in vivo has been demonstrated by crossing the viable Kaiso–/y mutant mouse to a mouse model of Wnt-driven intestinal cancer, which leads to a suppression of tumorigenesis that is thought to be due to derepression of methylated or unmethylated Wnt target genes. However, an equally plausible hypothesis is that tumor suppression arises as a consequence of perturbed p120ctn as it has been recently demonstrated that the complex that marks β-catenin (the key mediator of Wnt signaling) for destruction also has the ability to degrade p120ctn. 161
Much of the current work focuses on the identification of the relevant Kaiso-silenced genes in cancer. Given Kaiso’s ability to repress transcription at both methylated and unmethylated promoters, it is currently difficult to assess how important its mCpG binding role is in cancer. For example, in lung cancers, Kaiso mediates silencing of both the methylated MTA2 gene and the unmethylated CCND1 and MMP7 genes. 162 Given that all these genes are implicated in tumorigenesis, the precise significance of mCpG binding remains to be elucidated.
Beyond its role in transcriptional repression, Kaiso has also been localized to mitotic spindles and centrosomes at different stages of the cell cycle. 163 Altering the ability of Kaiso to locate to these regions resulted in defective centrosomes, whereas overexpressing Kaiso resulted in mitotic arrest, and knocking it down led to elevated proliferation. Given that Kaiso is often mislocalized in cancer, clearly these observations are of high potential significance to cancer; combined with the observation that the Kaiso-deficient mouse is healthy and viable, it makes a potentially attractive therapeutic target. However, given its diversity of roles, great care will need to be taken in the use of such a therapeutic.
ZBTB4 and ZBTB38
The genes for ZBTB4 and ZBTB38 are located on chromosomes 17 and 3, respectively. Unlike Kaiso, these proteins can bind a single methylated CpG in vitro and in vivo. 149 As yet, there are very little published data on these proteins in terms of biological roles, although ZBTB38 has been shown to play a role in human height determination. 164
With respect to cancer, there is a single report of ZBTB4 associating with the Sin3/HDAC complex and silencing the CDKN1A gene in neuroblastoma cells. 165 As would be expected, silencing of this gene resulted in cell cycle arrest due to p53 activation, suppression of apoptosis through regulation of p21CIP1, and promotion of long-term cell survival. This was reflected in the primary human neuroblastoma tumors, which showed decreasing ZBTB4 levels as the tumors progressed.
A p53-independent apoptotic role has also been confirmed for ZBTB38 in the mouse through its interaction with CtBP. 166 A role in apoptosis for ZBTB38 is also suggested as it is a substrate for caspase-3, a feature of mitochondrial-driven apotosis. 167 However, the significance of either of these genes in cancer therefore remains largely undetermined.
SRA Domain–Containing Proteins
There are 2 members of this family, UHRF1 and UHRF2, which contain a SET and RING finger–associated (SRA) domain as well as a ring finger motif, PHD finger, and ubiquitin-like domain. The ability of the SRA domain to bind methylated CpG was first recognized in human breast cancer cells 28 as decreased expression and increased promoter methylation of EGR2 correlated with increased expression of UHRF1, which associated with the methylated EGR2 promoter. UHRF2 was subsequently identified as a MBP due to its homology with the SRA domain of UHRF1 168 ; however, there is little similarity between this domain and the MeCP2_MBD in terms of recognizing and binding mCpGs as the SRA domain prefers hemimethylated DNA, and its mode of recognition involves flipping methylcytosine out of the DNA helix.169-171 By contrast, the MeCP2_MBD prefers symmetrically methylated DNA. 172 It folds into an α/β sandwich, which presents a large, positively charged surface 173 that recognizes hydration of the major groove of methylated DNA rather than cytosine methylation. 172
UHRF1
Prior to the discovery that UHRF1 (also known as Np95 and ICBP90) on chromosome 19 could bind methylated DNA, it had been identified as a protein of many functions. These include regulation of topoisomerase IIα gene expression 174 and regulation of cell proliferation, 175 the cell cycle, 176 apoptosis, 177 and tumor biology.178,179 Initially, its roles in these functions were thought to be related to its role as a transcriptional repressor as UHRF1 interacted with HDAC1 and recruited it to methylated tumor suppressor genes. 180 However, its role in human biology and cancer has taken on a wider significance due to the observation that UHRF1 associates with DNMT1 to maintain the epigenetic inheritance of DNA methylation.181,182 UHRF1 has an affinity for hemimethylated CpG DNA and recruits DNMT1 to ensure that the sequence becomes completely methylated following replication. This role may also apply to the maintenance of histone marks. 183 Additionally, it also has been shown to play a role in DNA damage response as it localizes to DNA damage and double-strand breaks.184,185
In human cancer, all of these functions have been shown to be relevant to different types of cancer. The maintenance of methylation plays a significant role in cancers by maintaining promoters in a hypermethylated state. In lung cancer, UHRF1 controls the cell cycle through maintenance of promoter methylation at CDKN2A and RASSF1. 186 It has also been shown to silence the CMV promoter in ES cells prior to methylation due to its interaction with Dnmt3a and Dnmta3b. 187 On the other hand, its roles in cancer may prove beneficial as, for example, depletion of UHRF1 has been shown to activate the DNA damage response leading to cell cycle arrest at G2/M and subsequent caspase 8–induced apoptosis. 188 In this respect, it is interesting that polyphenols in grapes have been shown to inhibit growth and induce apoptosis through downregulation of UHRF1.189,190 These apoptotic effects may also account for the radiosensitizing effect of UHRF1 depletion in human breast cancer cells. 191 Furthermore, its function as a transcriptional repressor has the potential to overcome drug resistance as overexpression of UHRF1 inhibits the upregulation of MDR1, which normally engenders drug insensitivity by actively removing cytotoxic drugs from breast cancer cells. 192
The many roles for UHRF1 in cancer have marked it for therapeutic targeting.193,194 However, as for all the MBPs in this review, their wide-ranging functions in multiple tissues again urge caution when targeting them. In this respect, the role played by UHRF1 in maintaining DNA methylation patterns required for progenitor maintenance and self-renewal of somatic tissue is of particular importance. 195
UHRF2
UHRF2 (also known as NIRF or Np97) was identified due to its role in the cell cycle. 168 Its expression is highest in proliferating cells, where it ubiquitinates PCNP,168,196 and its overexpression results in G1 arrest, presumably through an interaction with Cdk2. 197 Any other roles have yet to be described for this protein, but it may play a role in lung cancer cells as UHRF2 expression has been shown to be repressed by let7-a miRNA, which subsequently led to elevation of p21waf1 levels in vitro and in vivo and in growth suppression. 198 Any wider consequences of UHRF2 activity in cancer remain to be established.
Summary
This review has highlighted the many and intricate roles that MBPs play in human biology. Primarily, this involvement in gene regulation has led to them being utilized by normal cells to exert transcriptional control but has also led to them being exploited by cancer cells to escape such control. The dramatic effects of suppressing or enhancing their roles in cancer make them eminently suitable targets for therapeutic intervention. However, their many basic regulatory roles in a variety of tissues in the human body also present us with limitations and hazards of any such targeting. For this reason, we now need a much better understanding of the specific changes that these proteins induce in cancer cells to permit their propagation. The hope is that we can identify a subset of targets that are highly specific to cancer cells and so begin to develop a range of novel targeted therapies.
Footnotes
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The author(s) received no financial support for the research, authorship, and/or publication of this article.