
Abstract
The long-snouted bat (Platalina genovensium) is an endemic nectar-feeding bat from arid regions in western Peru and Chile. In certain areas of its distribution, it is likely the only bat pollinator and seed disperser of plants, and it is currently highly endangered. However, some aspects of its ecology, diet, and pollinator role remain unknown, especially in the unique and endangered lomas ecosystem. We conducted monthly evaluations of bats and plants over 15 months in the lomas of the National Reserve of Lachay (Peru) by sampling in three different plant communities along the Guayabito gorge. We collected fecal and pollen fur samples from 16 individuals of P. genovensium and recorded the phenology of its potential plant food resources. We differentiated fecal sample components and then identified and counted all pollen morphotypes in order to estimate their frequency and abundance in the samples. We also analyzed the niche breadth. Our results showed that the dietary components of P. genovensium included nectar, pollen, fruits, and insects, and that it utilized the cacti community for feeding. Pollen of the genus Haageocereus (Cactaceae) was the most frequent and abundant resource recorded, which was available all year long; moreover, the niche breadth analysis revealed that P. genovensium has a specialized diet on the Haageocereus cactus in Lachay. Finally, we suggest that P. genovensium could serve as a pollinator for Haageocereus acranthus, Haageocereus pseudomelanostele, and Puya ferruginea (Bromeliaceae) due to the large pollen loads it carries and the specific placement of pollen on different body zones.
Introduction
Along the arid regions of western South America dwells the endemic long-snouted bat Platalina genovensium (Phyllostomidae: Lonchophyllinae), a nectar-feeding species distributed from northern Peru to northern Chile, with an elevation from near sea level to 2,500 m above sea level (masl; Ossa, Vilchez, & Valladares, 2016; Sahley & Baraybar, 1996; Velazco, Cadenillas, Centty, Huamaní, & Zamora, 2013). This species attracts particular attention due to its rare occurrence along its described geographical distribution, unique morphology (long skull and long body size, Simmons & Wetterer, 2002), monotypic status (only species in the genus), phylogenetic position, and threatened status (Velazco et al., 2013). In addition, it may be the only potential bat pollinator and seed disperser of plants in some areas of its distribution (Sahley & Baraybar, 1996). Previous studies have suggested that its presence in arid regions is related to the presence of columnar cacti species, such as Weberbauerocereus weberbaueri, el candelabro Browningia candelaris (Sahley & Baraybar, 1996), Neoraimondia arequipensis (Aragón & Aguirre, 2007; Malo de Molina, Velazco, Pacheco, & Robledo, 2011), Corryocactus brevistylus (Aragón & Aguirre, 2007), Armatocereus procerus, and W. rauhii (Zamoraet al., 2013). Moreover, its particular morphology (long forearm, long skull, and great body weight) places it within the so-called cactophilic bats (Simmons & Wetterer, 2002). In fact, two studies in southern Peru that evaluated the diet of P. genovensium from fecal samples recorded pollen, fruits seeds, or fruit pulp belonging to the cacti species W. weberbaueri (Sahley & Baraybar, 1996), W. rauhii, and A. procerus (Zamora et al., 2013). Paticularly, Sahley and Baraybar (1996) recorded a high dependence of P. genovensium on the columnar cactus W. weberbaueri in arid zones of southwestern Peru. There are no other studies describing the diet and pollinator role of P. genovensium in different areas of its distribution.
Previous studies also described several threats affecting P. genovensium populations, which Velazco et al. (2013) summarized as follows: (a) a decline of columnar cacti populations due to the expansion of human population centers, (b) human disturbance of roost sites, and (c) hunting pressure due to traditional medicinal uses. These anthropogenic factors endanger P. genovensium populations; therefore, this species is included in red lists, as near threatened in the IUCN (2015) and critically endangered in Peruvian orders (Ministerio de Agricultura, 2004).
In arid regions where P. genovensium occurs, there is also a unique ecosystem called the lomas formation. This ecosystem is formed by coastal small hills (or lomas) located along the pacific desert, where islands of vegetation are supported by seasonal, marine fog (Dillon & Hoffmann, 1997). This ecosystem harbors the highest diversity of organisms along the coastal arid regions, many of which are endemic to the lomas. For instance, more than 40% of recorded plant species in the lomas are endemic species (Duncan & Dillon, 1991). The lomas ecosystem is threatened by the advance of human populations and human recreational activities (Cavieres et al., 2002; Pinto, Barria, & Marquet, 2006), cattle, and deforestation (Dillon & Hoffmann, 1997).
In this study, we addressed four specific questions about the diet and pollinator role of P. genovensium in the lomas, an ecosystem where the species has not been studied. First, what are the dietary components (e.g., insects, fruits, plants) of P. genovensium? Second, which plants does P. genovensium feed on? Third, does P. genovensium have a generalist or specialist diet? Finally, could P. genovensium be a pollinator for the plant species it feeds on? Answers to these research questions will help to more accurately inform appropriate conservation actions and improve current management efforts for this endangered bat species in the unique lomas ecosystem.
Methods
Study Area
We conducted the study in the Guayabito gorge (11°21′30″ S, 77°22′10″ W), located in the northeastern part of the National Reserve of Lachay (Lachay, from this point forward) in Lima, Peru (Figure 1). Lachay preserves more than 5000 ha of coastal lomas (0–800 masl; Cano et al., 1999). This ecosystem is characterized by a seasonal presence of fog coming from the ocean, which allows for the growth of vegetation with a high degree of endemism (Dillon & Hoffmann, 1997). The weather and vegetation vary seasonally throughout the year. The drier season (November–May) is characterized by high temperatures (20℃–23℃) and low relative humidity (90%–83%), while the humid season (June–October) is marked by low temperatures (13℃–16℃) and high relative humidity (95%–100%; Kunimoto, De La Cruz, Arana, & Ramírez, 2002). We conducted monthly evaluations of bat and plant communities between September 2012 and November 2013.
Bats Capture
We assembled mist nests for bat capture in three different plant communities in the Guayabito gorge: (a) herbaceous and semi-woody plants (i.e., where herbaceous plants were dominant with some semi-woody plants present), (b) woody plants (i.e., areas with trees even though they were not the dominant life forms), and (c) cacti and succulent plants (i.e., dominated by cacti). We assembled four to six mist nets per plant community (mist net sizes: length 6, 9, or 12 m; height 3 m) at different heights (0–4 m). We operated mist nets from 1800 to 0100 h during two nights per month. After capture, we extracted pollen from the fur by gently pressing transparent adhesive tape against four different body zones (Muchhala & Jarrín-V, 2002): snout, head (above the eyes), back, and chest. We kept captured bats in cloth bags for 2 h to allow for fecal sample collection, which were preserved in 70% ethanol (Maguiña, Amanzo, & Huamán, 2012). After samples collection, all individuals were released. All procedures complied with the “Guidelines of the American Society of Mammalogists for the use of wild mammals in research” (Sikes, Gannon, and the Animal Care and Use Committee of the American Society of Mammalogists, 2011); in addition, the Institutional Ethics Committee from Universidad Peruana Cayetano Heredia approved our capture and collection procedures.
Plant Phenology
We established 12 10 × 10 m plots to monitor plant phenology in the same plant communities mentioned earlier, with four plots per plant community separated from one another by at least 100 m. To evaluate phenology, we recorded and counted the numbers of buds and flowers of species potentially visited by bats. Additionally, we sampled flowers from all available plant species in Guayabito gorge to establish a pollen reference collection.
Sample Analysis
In the laboratory, we observed pollen samples collected from the fur under the microscope at 10× to 40× magnification. We subjected fecal samples to stereoscope observation and Erdtman’s acetolysis process before microscopic observations at 40× magnification (two slides per sample, following the methodology of Maguiña et al. [2012]). We recorded four categories of fecal sample components: (a) pollen, (b) plant tissue, (c) seeds, and (d) insects. We identified and counted all pollen morphotypes to calculate frequency and abundance per plant species. Based on frequencies and abundance of pollen, we calculated Levins standardized Index (BA) to analyze niche breadth. BA ranges from 0 (only one resource used) to 1 (all available resources used evenly; Krebs, 1999).
Results
Diet and Pollinator Role of P. genovensium
We captured 16 individuals of P. genovensium. We collected fecal samples from 7 individuals and pollen fur samples from 16 individuals (56 adhesive tape samples in total). Ten of the bats were captured in the herbaceous plant community while the remaining were captured in the cacti and succulent plant community. None were captured in the woody plant community. The fecal samples contained the following dietary components: pollen (100% of the samples), plant tissues (42.8%), and insects (14.3%); we did not record seeds in the samples. The pollen samples from both the fecal and fur samples belonged to nine plant taxa, with Haageocereus being the most frequent (N = 16) and abundant (>90% of all pollen grains recorded), followed by cf. Senecio lomincola (Asteraceae), Puya ferruginea (Bromeliaceae), and cf. Solanaceae (Figure 2).
Study area in the national reserve of Lachay. Frequency (left axis) and abundance (right axis) of plant species recorded in fecal and fur samples of Platalina genovensium based on pollen analysis. cf. SOL = cf. Solanaceae; Ind 3cp = Indeterminate 3-colporate; AST = Asteraceae; SOL = Solanaceae; POA = Poaceae; Ind 3 p = Indeterminate 3-porate.

Frequency and Average Abundance of Pollen Grains per Plant Species Carried on Platalina genovensium’s Fur.

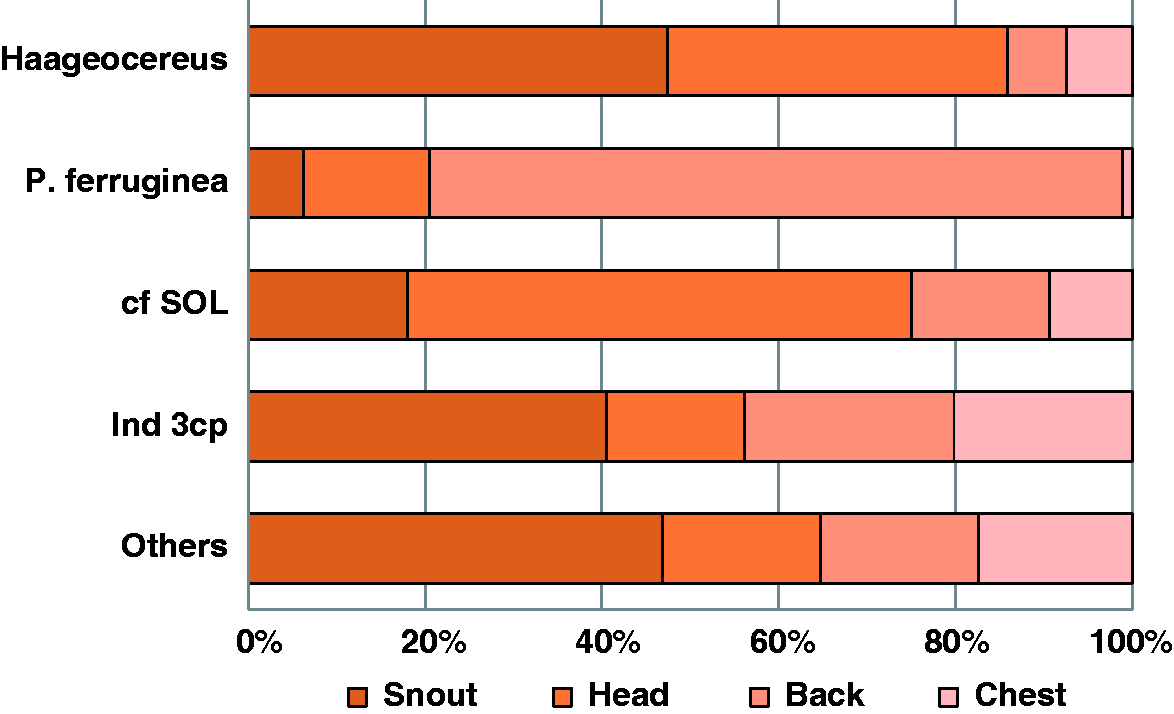
Percentage of pollen per plant species placed in different bat’s body zones. cf. SOL = cf Solanaceae; Ind 3cp = Indeterminate 3-colporate.
Plant Phenology
We recorded phenology for nine potentially bat-visited plant species: Stenomesson coccineum (Amarylidaceae), P. ferruginea (Bromeliaceae), Haageocereus acranthus subsp. olowinskianus, Haageocereus pseudomelanostele (Cactaceae), Capparis prisca (Capparidaceae), Caesalpinia spinosa (Fabaceae), Exodeconus prostratus, Nolana difusa, and Nicotiana paniculata (Solanaceae). The species of Cactaceae and Solanaceae families presented the longest blooming periods throughout the year, with H. acranthus blooming for almost the whole year. The remaining species had shorter blooming periods, which lasted between 2 and 6 months (Figure 4). We recorded four of these species in P. genovensium’s diet: H. acranthus, H. pseudomelanostele, and P. ferruginea.
Percentage of flowers and buds per plant species and percentage of captured bats along the study period (September 2012–November 2013).
Discussion
For the first time, we present information about the diet and pollinator role of P. genovensium in the lomas ecosystem. We found that its diet is mainly composed of floral resources and added at least four new plant species to the list of its consumed resources. This bat mainly feeds from the cacti species of the genus Haageocereus suggesting that its diet is fairly specialized on these resources. Finally, we found that P. genovensium carries large pollen loads and that the placement of pollen from different plant species is on different body parts of the bat, suggesting that P. genovensium likely serves as a pollinator for the cacti species Haageocereus acranthus and H. pseudomelanostele as well as the bromeliad species P. ferruginea.
Our capture results show that P. genovensium is most common in herbaceous plant communities within the lomas. We also captured several individuals in cactus-dominated communities, which was an expected result because cactus pollen and nectar serve as important food resources (Sahley & Baraybar, 1996). Bats captured in both herbaceous and cactus-dominated communities had abundant cacti pollen on their fur, implying that bats feed primarily in cactus-dominated plant communities and only use the herbaceous plant communities to reach other feeding areas. P. genovensium may prefer to commute across herbaceous, rather than woody, plant communities, as these bats are known to fly close to ground level (Velazco et al., 2013).
The diet of P. genovensium in the lomas of Lachay was composed mainly of floral resources (nectar and pollen) and complemented by other plant resources and insects. Contrary to the studies of Sahley and Baraybar (1996) and Zamora et al. (2013), we did not find seeds, but we did find vegetative tissues that were possibly derived from fruits. The BA based on pollen frequencies was higher (0.6965) than the BA based on pollen abundance (0.0164). We suspect that the low sample size (N = 16) inflated the value of BA based on pollen frequencies. Therefore, the abundance-based niche breadth might be a more precise representation of the species’ ecology. For this reason, we concluded that the population of P. genovensium in Lachay is fairly specialized on Haageocereus cacti. There is a scarcity of information about measurements of niche breadth for P. genovensium or other nectarivorous bats. We found only one study for the nectarivorous tailed tailless bat Anoura caudifer (Maguiña et al., 2012), and in its cloud forest ecosystem, the authors reported a low BA value (0.045, based on pollen abundance). On account of the large number of resources available for bats in cloud forests, we expected that P. genovensium would show a narrower niche breadth due to their more arid ecosystem, where the number of food resources is relatively limited.
In total, we recorded nine different plant taxa in fecal and fur pollen samples, from five plant families. Due to their ecology and floral characteristics, we consider that only Cactaceae, Bromeliaceae, and Solanaceae families are actively sought out by P. genovensium during foraging. Asteraceae flowers are rarely used and pollinated by bats (Pruski & Sancho, 2004); we suspect that its occasional appearance in pollen samples is due to its high abundance in the lomas and the low flying activity of bats, which would bring their bodies close to these plants. Similarly, pollen of Poaceae could be ingested or attached to bat’s fur during flight. Additionally, pollen abundance of Asteraceae and Poaceae was low compared with the other families registered (Table 1). Before our study, the known diet of P. genovensium included only four species of cactus: W. weberbaueri, the candelabro Browningia candelaris (Sahley & Baraybar, 1996), W. rauhii, and A. procerus (Zamora et al., 2013). Our findings add at least four new plant species to the species’ diet.
The genus Haageocereus had the highest frequency of occurrence in the samples. We recorded two species of Haageocereus in the Guayabito gorge, H. acranthus and H. pseudomelanostele, but due to their similarity, it was not possible to identify Haageocereus pollen to species level. Both species present floral characteristics (corolla funnel shape, pale colors, high nectar, and pollen production) similar to other cacti used by bats, such as the organ pipe cactus (Stenocereus thurberi) in northern Mexico (Fleming, Sahley, Holland, Nason, & Hamrick, 2001), W. weberbaueri in southwestern Peru (Sahley & Baraybar, 1996), Pachycereus pecten-aboriginum in western Mexico (Tschapka, Sperr, Caballero-Martínez, & Medellín, 2008), and Pilosocereus moritzianus, Subpilocereus repandus, and Subpilocereus horrispinus in northern Venezuela (Nassar, Ramirez, & Linares, 1997). Moreover, they both have long blooming periods, thus offering food resources to bats all year long. We highlight that this is the first record of Haageocereus being visited by nectarivorous bats. The species P. ferruginea (Bromeliaceae) was previously recorded as consumed by bats, particularly in lower elevations of its distribution (Kessler & Krömer, 2000). P. ferruginea possesses extremely large inflorescences capable of producing many flowers attractive to bats. Other bat-pollinated plant characteristics include its inconspicuous flower color and tube-shape corolla. Within the family Solanaceae, we recorded two taxa consumed by bats. Inspite of this, these plant species did not possess as many characteristics related to bat-consumption as the previous cases. It is possible that long-snouted bats feed on it to complement their diets.
The number of pollen grains carried by long-snouted bats showed their capacity for transferring large amounts of pollen. Logically, the amount of transported pollen will vary according to the amount of pollen produced per plant species. The largest amount of transported pollen was from Haageocereus, which could be explained by larger amounts of produced pollen, given the increased number of stamens (>100, Nassar et al., 1997) compared with other plant species recorded in the samples (≤6). Previous studies of cacti pollination biology report that a cactus stigma potentially holds more than 104 pollen grains, while the mean number of ovules per flower exceeds 1,000 (Nassar et al., 1997). We predict that P. genovensium likely acts as an efficient pollinator because the amount of Haageocereus pollen carried by individual bats would be large enough to fertilize the ovules. At the same time, additional pollinator exclusion experiments are required to confirm the pollinator role of P. genovensium for species of Haageocereus.
The placement of pollen on different body parts also supports the pollinator role of P. genovensium. In the case of Haageocereus pollen, which was mainly located on the snout and head, we posit that each anatomical position corresponds with the two Haageocereus species; the pollen on the snout possibly corresponds to H. pseudomelanostele, which has shorter flowers (corolla length = 5.05 cm ± 0.42, from the base of nectar chamber to upper-corolla), while the pollen on the head possibly corresponds to H. acranthus, which has larger flowers (corolla length = 7.7 cm ± 0.53). Further studies are necessary to test this hypothesis and the precision of pollen transfer for avoiding interspecific transfer between H. acranthus and H. pseudomelanostele, considering the similar morphology of the flowers, including actinomorphic corollas and the spiral arrangement of the numerous stamens. If this hypothesis is supported, this would be a new example of how variation in placement of pollen on the bodies of bats can reduce the competition for pollination among sympatric species, as demonstrated for other bat-pollinated species of the genera Burmeistera (Muchhala & Potts, 2007) and Marcgravia (Tschapka, Dressler, & von Helversen, 2006).
With regard to the other pollen morphotypes, P. ferruginea showed specific placement on the back of bats (80%). Given the plants’ large corolla and stamens, this placement diminishes the probability of foreign pollen reaching the stigma. It is unclear whether P. genovensium is a relevant pollinator for other species (e.g., Solanaceae). The small pollen size in Solanaceae (smaller than Haageocerus and P. ferruginea), the small amount of pollen carried by bats, and placement on the same bat body zones as Haageocereus and P. ferruginea pollen could complicate successful pollen delivery to the stigma of Solanaceae species.
Implications for Conservation
The lomas in Lachay offers important food resources for the P. genovensium population, and this study provides incentives to preserve this unique ecosystem and the species that inhabit it. The food resources for P. genovensium principally occur in cactus- and succulent plant-dominated communities. While we only evaluated one of the many gorges present in Lachay, others are commonly dominated by cacti and may also provide critical food resources for these endangered bats, as well as roosts and mating sites. Currently, the Peruvian National System of Protected Areas includes only the Natural Reserve of Lachay (5,070 ha) and the Reserved Zone Lomas de Ancon (10,962 ha). Although lomas occur in approximately 40 more sites along the coastal Peruvian desert (Sernanp-Perú, 2016). And in Chile, the National System of Wild Protected Areas do not include any lomas ecosystem (Conaf-Gobierno de Chile, 2016). As mentioned earlier, one of the threats affecting P. genovensium populations is the decline of columnar cacti due to the expansion of human population centers (Velazco et al., 2013). The Haageocereus species mentioned in this study are also under high risk; both species are endemic to the desert scrub in central and northern Peru (Calderón Moya-Méndez, Ceroni Stuva, & Ostolaza Nano, 2004). Moreover, H. acranthus spp. olowinskianus and H. pseudomelanostele were categorized as vulnerable and endangered, respectively, by the study of Calderón Moya-Méndez et al. (2004). Therefore, we suggest taking additional actions to preserve wider areas of lomas, especially if they harbor Haageocereus cacti species, in order to preserve P. genovensium populations.
Footnotes
Acknowledgments
The authors are grateful to all the volunteers for assistance during fieldwork. Carlos Ostolaza for helping in the identification of cacti species. Luis Huamán, director of Laboratorio de Palinología y Paleobotánica at Universidad Peruana Cayetano Heredia, where we conducted the pollen analysis. Michael O. Dillon for providing literature about the lomas ecosystem. Michael Montague, Leighton Reid, Nathan Muchhala, Mónica Carlsen, Serena Achá, Diana Gamba, Camilo Calderón-A, and anonymous reviewers for their suggestions to improve an early manuscript version of this study. The National Reserve of Lachay granted permission to work in the study area.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was partially supported by the Ann Kotowski Grant from Universidad Peruana Cayetano Heredia.