
Abstract
One way to draw attention to an area regarding conservation is to declare it as an Atlantic Forest Biosphere Reserve (RBMA). In this article, we analyzed attributes related to different forest remnants, including the only RBMA of Santa Catarina state (Brazil), a modified old-growth Subtropical Atlantic Forest remnant. We brought evidences that patterns of distribution of species abundance and species diversity are influenced by highly dominant species. We found a relevant proportion of endemic tree/shrub species and three endangered species. These findings demonstrate the relevant biodiversity of the Subtropical Atlantic Forest of Santa Catarina. The diversity profiles of the RBMA and other areas overlapped. This finding, which has an important conservational implication that the tree species diversity in these areas is similar. Therefore, we concluded that the areas considered in our study deserve attention regarding biological conservation. New priority areas for conservation are necessary, and the establishment of new RBMA may be a way to achieve this goal.
Introduction
Biological diversity is a central theme in ecology and is often expressed in terms of species diversity (Liu, Whittaker, Ma, & Malcolm, 2007). This concept considers two attributes: (a) species richness and (b) the uniformity of distribution of species’ abundance (Hill, 1978; Hurlbert, 1971; Melo, 2008). Therefore, understanding the interaction between these attributes and their relation with the conservation status of a biological community is crucial, even more when species diversity is regarded as an essential attribute for identifying priority areas for conservation (Buckland, Studeny, Magurram, & Newson, 2011; Durigan et al., 2009).
The growing importance of Brazilian’s Atlantic Forest is stimulating the scientific community and government agencies to select priority areas for conservation (Anacleto, Ferreira, Diniz Filho, & Ferreira, 2005; Brasil, 2002, 2007a, 2007b; Durigan et al., 2009; Durigan, Siqueira, Franco, & Ratter, 2006; Galetti et al., 2009; Santos & Mantovani, 1999). These areas are usually selected based on the occurrence of rare and endangered species, species richness, level of endemism, and vegetation types (Joppa, Visconti, Jenkins, & Pimm, 2013; Margules & Pressey, 2000; Mews, Pinto, Eisenlohr, & Lenza, 2014; Pullin, Sutherland, Gardner, Kapos, & Fa, 2013). However, studies in tropical forests have suggested that species diversity is an important criterion, for it can vary among fairly conserved forests (Giam, Scheffers, Sodhi, Wilcove, Ceballos, & Ehrlich, 2012; Gibson et al., 2011; Sevegnani et al., 2013; Sevegnani, Vibrans, & Gasper, 2013). In this sense, species diversity plays a key role in defining the conservation status of tropical forests remnants.
On the other hand, if the community has few species with great abundance, some of the aforementioned issues might become contradictory. Hence, species diversity metrics must be used carefully (Imai et al., 2014). Well-conserved forests, for example, may yield lower species diversity when compared with less conserved forests, especially when using indices that address greater weight to the uniformity of species’ abundance distribution (Magurran, 2004; May, 1975; Melo, 2008; Oliveira, 2015). In addition, in some subtropical forests, the abundance and relative dominance of dominant species can be inversely proportional to the values yielded by species diversity indices (Oliveira, 2015).
Patterns of distribution of species abundance should be carefully verified when selecting areas for conservation, as they are related to the vegetation’s conservation status (Magurran, 2007). Very altered tropical forests might present few species with great abundance, and this could be related to species’ ecological adaptations (Pitman et al., 2001), to interspecific competition for space and resources (McGill et al., 2007) or, still, to processes of ecological succession (Volkov, Banavar, Hubbell, & Maritan, 2007). Meanwhile, these factors can also influence the formation of distribution patterns in conserved forests, where resource-demanding species may present great abundance, such as the brazilian-pine (Araucaria angustifolia [Bertol.] Kuntze) and xaxim (Dicksonia sellowiana Hook.) in the Mixed Forest with Araucaria and palmiteiro (Euterpe edulis Mart.) in the Subtropical Rainforest of Southern Brazil (Klein, 1978).
In Brazil, a program supported by the government called Atlantic Forest Biosphere Reserve (RBMA) has been standing out for selecting priority areas for conservation. The RBMA features a set of guidelines defined by UNESCO (RBMA, 2008) for selecting outposts which are intended to become centers of dissemination of ideas, concepts, programs, and projects developed by the each reserve (RBMA, 2014). One of the priority areas for conservation in the state of Santa Catarina (Southern Brazil) is the RBMA Chácara Edith Private Reserve of Natural Heritage (RPPN Chácara Edith). This reserve is the only RBMA in the state (ICMBio, 2011; RBMA, 2014). In this study, we considered the RPPN Chácara Edith as the reference area regarding ecological aspects of the vegetation. Therefore, due to the high degree of anthropic modification in the Subtropical Rainforest of Santa Catarina, we ask: Would not it be relevant that other areas might be regarded as priorities for conservation? On the basis of this question, we developed a case study involving forests remnants in different conservation status to demonstrate how differences in the patterns of distribution of tree species’ abundance—but not in species diversity—would reveal relevant implications for selecting areas for conservation. Our hypothesis was that the conservation status of a given forest remnant is not necessarily related to its species diversity, once dominant species lead to less uniform communities and, therefore, diversity indices would yield small values (Magurran, 2004; May, 1975; Melo, 2008; Oliveira, 2015). In addition, based on data from the considered forests remnants, we listed endemic and endangered tree/shrub species aiming to support the ongoing calling for conservational efforts in the Subtropical Atlantic Rainforest of Santa Catarina.
Methods
Study Area
The study area is inserted in the Coastal Mountain Range of Santa Catarina, Southern Brazil (Figure 1). The Subtropical Rainforest of Santa Catarina (Oliveira-Filho, 2015) state is placed between two different geomorphological units. The first is composed by the Serra do Tabuleiro/Itajaí complex, which is characterized by mountain ranges disposed in form of subplots with dissected relief, deep valleys with steep slopes, separated by sharped ridges; the second geomorphological unit is composed by the Coal zone depression in the southern portion of the state, presenting hilly landscape with valleys, steep slopes, and concave–convex relief with open valleys (Santa Catarina, 1986).
Delimitation of forest remnants of the Subtropical Rainforest of Santa Catarina according to SOS Atlantic Forest Foundation & INPE (2013), and the location of the 12 studies considered in this research.
According to the Köppen classification, the Subtropical Rainforest of Santa Catarina is influenced by two climate types: Cfa—humid mesothermal subtropical climate without dry season with a hot summer and Cfb—humid mesothermal subtropical climate without dry season with a mild summer (Alvares, Stape, Sentelhas, Gonçalves, & Sparovek, 2013). Due to environmental and altitudinal variations, the average annual temperature ranges from 11℃ to 20℃. Rainfall patterns may vary according to the altitude, ranging between 1,100 and 1,300 mm in the northern portion of the state (Pandolfo et al., 2002).
Data Collection
Number of Sampled Individuals (n), Species Richness (S), Shannon’s Index (H′), and Simpson’s Index (1/D) for Different Studies Conducted in the Subtropical Rainforest of Santa Catarina, Southern Brazil.
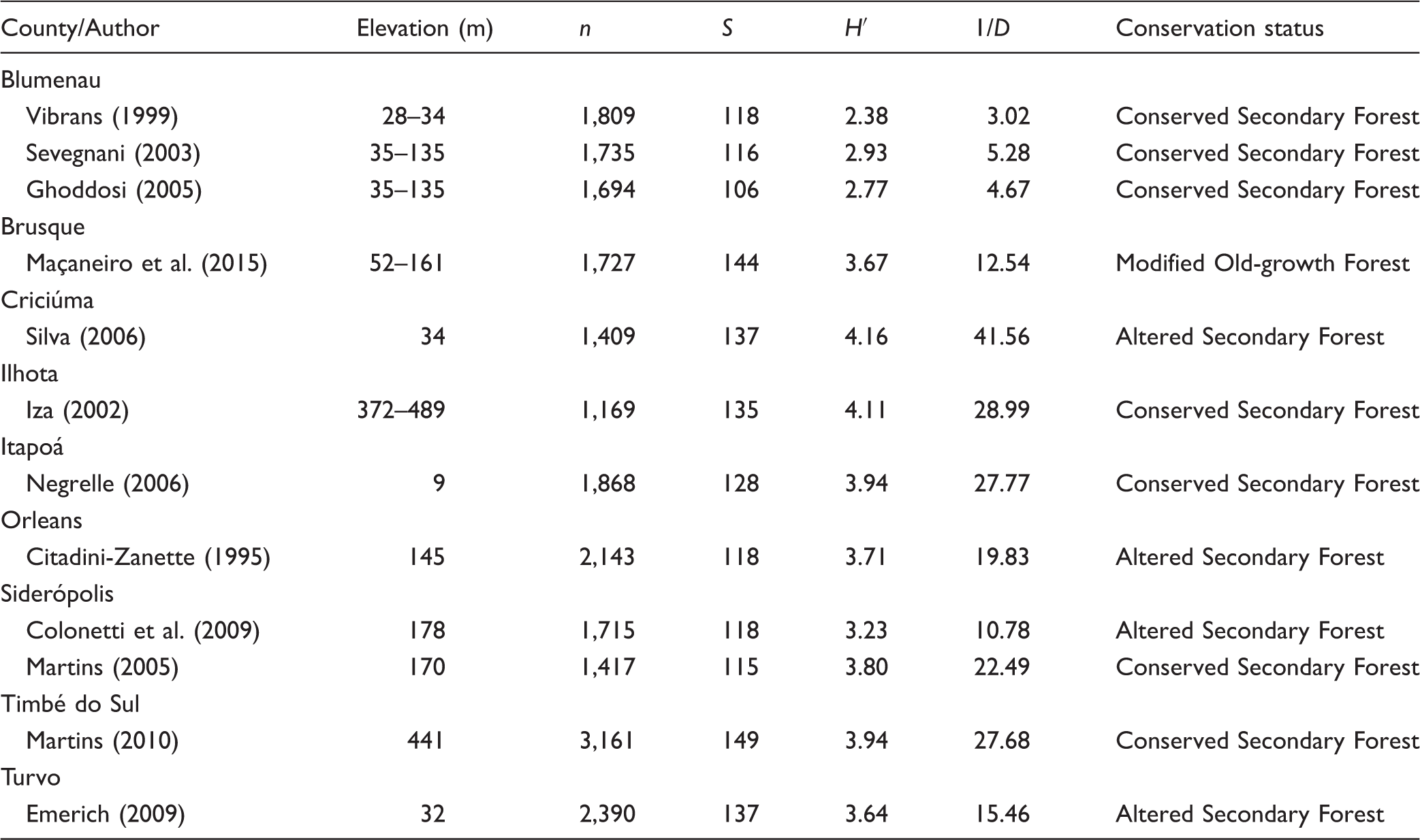
We classified the conservation status of the vegetation of each study based on the description provided by the author(s). We defined three categories: (a) Modified old-growth forest, where selective logging of species of economic interest was conducted, although the forest still presented composition, structure, and physiognomy of an old-growth vegetation; (b) Conserved secondary forest, where more intense logging was conducted when compared with the modified old-growth forest, but due to the abandonment of the area and regeneration time-lapse, the forest presented a well-conserved status; and (c) Modified secondary forest, where logging was conducted, although, because of the reduced regeneration time-lapse and other anthropic interventions, the forest still is considerably modified in its composition and structure.
We verified the patterns of distribution of species abundance (PDSA) through the Whittaker’s diagrams (Magurran, 2004). The similarities or differences among PDSA were tested by the Kolmogorov–Smirnov test for two samples, with α = .05 (Sokal & Rohlf, 1995). The Whittaker’s diagram is a useful tool to analyze PDSA in biological communities, such that contrasting patterns of species richness and vegetation uniformity may be addressed (Krebs, 2014; Magurran, 2004; Melo, 2008).
For the purpose of comparing the species richness of the studies, we constructed rarefaction curves using the Mao Tau method (Colwell, Mao, & Chang, 2004). We opted by the individual-based rarefaction, as multiple comparisons could be influenced by the tree density of each area (Gotelli & Colwell, 2001).
We estimated the heterogeneity (Krebs, 2014) through Shannon’s index (H', ln-base) and the reciprocal of Simpson’s index (1/D). These two indices differ in the weight given to rare species (Magurran, 2004). For generalizing the comparisons of species richness, H′ and 1/D, we built diversity profiles using the Rényi’s series (Tóthmérész, 1995). The diversity profiles allow effective comparisons among communities, because the influence of the weights attributed to rare and dominant species is not fixed (Leinster & Cobbold, 2012; Melo, 2008).
We investigated the relation between species diversity and the abundance of dominant species using Pearson’s correlation coefficient (r) and scatterplots through the following steps: (a) we correlated the sum of the relative densities (RD%) of the five species with greater abundance of each study with their respective H′ and 1/D; (b) we tested the significance (α = .05) of r through the corrected t test by Dutilleul (1993), which addresses the spatial autocorrelation amid observations, correcting the degrees of freedom; and (c) we built scatterplots of the diversity indices (y-axis) vs. RD% (x-axis).
To support the necessity of directing efforts towards the conservation of the studied area, we listed the tree/shrub species recorded in the 12 studies and classified them according to their endemism to Brazil and to the Brazilian Atlantic Forest using information from Flora do Brasil (Forzza et al., 2012). We defined the conservation status of the species based on the International Union for Conservation of Nature (IUCN) criteria (IUCN, 2003), using the database of Fundação Biodiversitas (www.biodiversitas.org.br).
Results
We found similar patterns of species richness among the study areas (Figure 2). According to the Whittaker’s diagram, the community sampled by Maçaneiro et al. (2015) presented greater uniformity (less inclined curve) and greater species richness (more extended curve) when compared with the other studies (e.g., Citadini-Zanette, 1995; Colonetti et al., 2009; Ghoddosi, 2005; Martins, 2005; Sevegnani, 2003; Vibrans, 1999). The PDSA found in Maçaneiro et al. (2015) was significantly different (Kolmogorov–Smirnov, p ≤ .05) when compared with other studies (Citadini-Zanette, 1995; Colonetti et al., 2009; Ghoddosi, 2005; Martins, 2005; Sevegnani, 2003; Vibrans, 1999). On the other hand, considering Emerich (2009), Iza (2002), Negrelle (2006), and Silva (2006), the PDSA found in Maçaneiro et al. (2015) was not significantly different (Kolmogorov–Smirnov, p > .05). With the standardization of 1,169 sampled individuals, the rarefaction curves indicated the studies with the greater species richness: Iza (2002) with 135 species; Maçaneiro et al. (2015) and Silva (2006), each one with 130 species, and Martins (2010) with 120 species (Figure 3).
Whittaker’s diagram showing the abundance (y-axis, Log10 scale), species richness, and the dominant species in different studies conducted in the Subtropical Rainforest of Santa Catarina, Southern Brazil. Individual-based rarefaction curves built by Mao Tau’s method, with their respective confidence intervals, for different studies conducted in the Subtropical Rainforest of Santa Catarina, Southern Brazil.

In the diversity profiles (Figure 4), we observed that species diversity varied among sites. Even though Maçaneiro et al. (2015) presented greater species diversity than other studies (e.g., Colonetti et al., 2009; Ghoddosi, 2005; Sevegnani, 2003; Vibrans, 1999), its diversity profile overlapped with the diversity profile of other sites (e.g., Emerich, 2009; Iza, 2002; Martins, 2005, 2010; Negrelle, 2006; Silva, 2006; Vibrans, 1999), suggesting that these sites are nonseparable (see Liu et al., 2007; Tóthmérész, 1995).
Diversity profiles build using Rényi’s series for different studies conducted in the Subtropical Rainforest of Santa Catarina, Southern Brazil. For the parameter α = 0, N0 = species richness; for α = 1, N1 = exp(H′), then, H′ = ln(N1); for α = 2, N2 = reciprocal of Simpson’s index (1/D).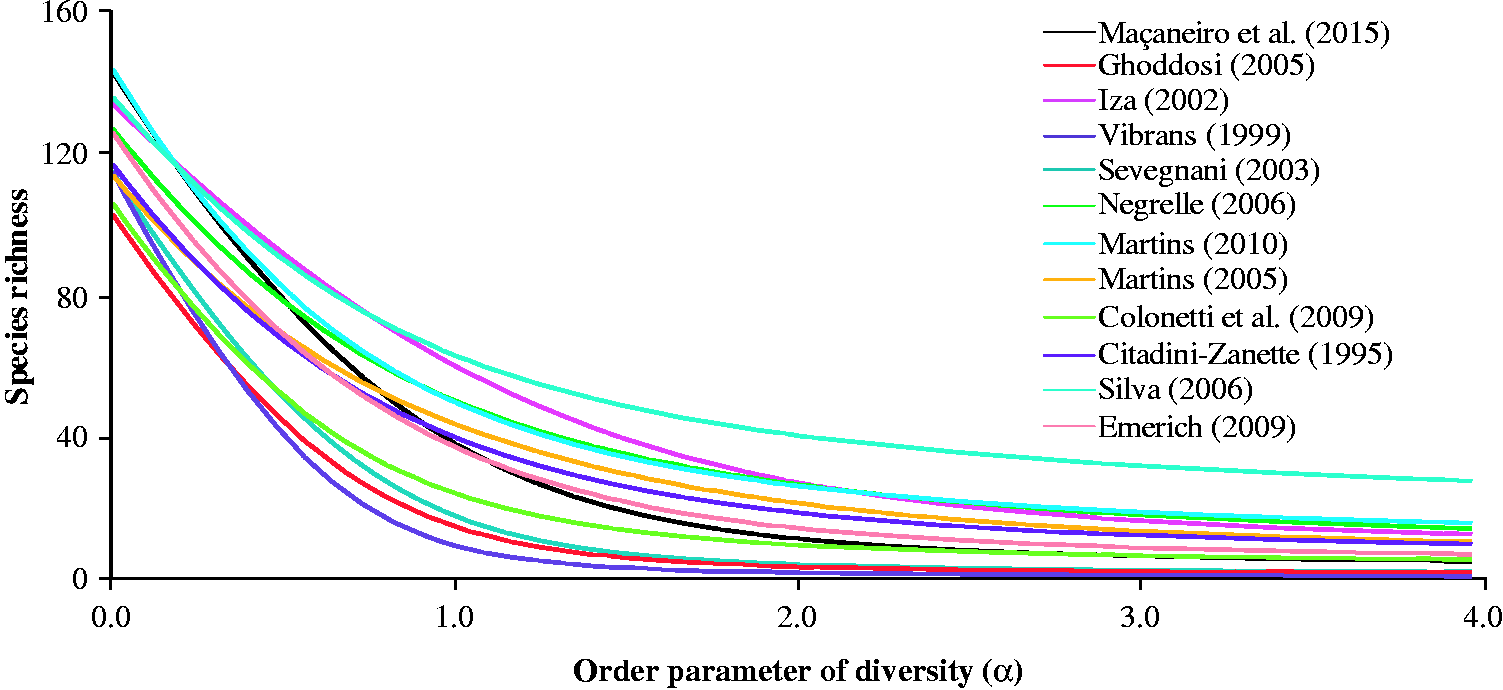
We found strong and highly significant correlations (p < .01) between the diversity indices (H′ and 1/D) and the RD% of the five most abundant species (Figure 5). Therefore, few dominant species may influence species diversity indices.
Relation between species diversity (H′ and 1/D) and the sum of the relative density of the five most abundant species in 12 studies conduct in the Subtropical Rainforest of Santa Catarina, Southern Brazil.
Among the 366 tree/shrub species registered in the compiled studies (Appendix), we detected three species classified as vulnerable (canela-preta—Ocotea odorifera [Vell.] Rohwer and imbuia—Ocotea porosa [Nees & Mart.] Barroso) and one species in danger (palmiteiro—Euterpe edulis Mart.), according to the Red List of Fundação Biodiversitas, which follows IUCN criteria (IUCN, 2003). Furthermore, when we analyzed the species’ endemism, we found 131 species (35.8%) occurring only in Brazil, 170 species (46.4%) occurring only in the Atlantic Forest, and 11 species (3%) occurring only in the Southern Brazilian Atlantic Forest.
Discussion
The tree species diversity of well-conserved sites in the Brazilian Atlantic Forest is not always superior to the diversity of less conserved areas in the same region—the species diversity of the RBMA (modified old-growth forest) is similar to secondary forests with different conservation status. However, it is important to highlight that this conclusion is based solely on tree species diversity with dbh ≥ 5.0 cm. Most likely, if other components were to be considered, such as understory plants, epiphytes and lianas, the species diversity would be greater in more conserved sites (Gasper et al., 2014; Leite & Klein, 1990). This hypothesis would be corroborated by studies contemplating various groups of plants using quantitative data collected in sample plots, instead of studies based solely on species checklists.
The translation of forest data into metrics of species diversity depends on the level of uniformity of the vegetation or, still, on the adopted diversity index. Despite the intrinsic characteristics of diversity indices regarding the weight given to rare and dominant species (Buckland et al., 2011; Hurlbert, 1971; May, 1975; Melo, 2008), our study brought evidence that H′ and 1/D are influenced by dominant species. This issue is relevant when dealing with selection of priority areas for conservation. In Brazil, for example, the use of diversity indices and their interpretation is not always meaningful. In many studies, the values yielded by diversity indices are simply reported and not discussed in-depth, they are rather merely compared with other studies. It is essential to clarify our perceptions about the concept of species diversity, as stressed by Hurlbert (1971), given the ongoing call for attention on secondary forests and their importance (Costanza et al., 1997; Gibson et al., 2011; Ricketts et al., 2005). Diversity indices could be usable as descriptors of species abundance distribution, especially in forest ecosystems with dominant species. However, multiple criteria must be considered when selecting the most suitable index (Buckland et al., 2011; Magurran, 2004). Although being very popular, the H′ is extensively criticized for being derived from the information theory and not from ecological perspectives (Hurlbert, 1971; Magurran, 2004). Thus, its value and unit do not have ecological meaning. Conversely, the Simpson’s index is recommended by several authors (Buckland et al., 2011; Hurlbert, 1971; Lande, 1996), for the reason that it generates more meaningful ecological information. Even though, like H′, this index is also influenced by dominant species. Therefore, when a more sharpened analysis about the attributes of a community is necessary, diversity profiles (Chao et al., 2014) and diversity partitioning (Lande, 1996) could be useful tools.
Our results revealed that PDSA are influenced by dominant species, such as the palm Euterpe edulis. Considering this species, two patterns could be established for the Subtropical Rainforest of Santa Catarina: (a) forests with great abundance of Euterpe edulis (e.g., Colonetti et al., 2009; Ghoddosi, 2005; Sevegnani, 2003; Vibrans, 1999) and (b) forests with less abundance of Euterpe edulis (e.g., Emerich, 2009; Iza, 2002; Maçaneiro et al., 2015; Martins, 2005, 2010; Negrelle, 2006; Silva, 2006; Vibrans, 1999). These patterns have important implications for the conservation of these areas, for differences in the abundance of Euterpe edulis could be related to variations in environmental conditions (Mélo, Budke, & Henke-Oliveira, 2013; Oliveira-Filho & Fontes, 2000; Pitman et al., 2002; Souza, Lelis, Schaefer, Souza, & Meira-Neto, 2012), anthropic influences (Laurance et al., 2006; Pitman et al., 2001; Schorn & Galvão, 2006, 2009), and vegetation type (McGill et al., 2007). For example, Mews et al. (2014) found that species abundance in different environments is a relevant criterion for selecting priority areas for conservation in the Brazilian Savanna (Cerrado), due to the fact that areas with different soil types have particularities regarding species abundance. Besides environmental predictors, the intensity of anthropic activities on the vegetation may also influence PDSA (Pereira, Oliveira-Filho, Eisenlohr, Miranda, & Lemos Filho, 2015), as we found that the compared areas presented distinct conservation status.
In addition to the conservation status of the vegetation, the uniformity of populations might explain the similarities (or differences) in PDSA (Magurran, 2004). According to Melo (2008), communities with greater uniformity will present greater species diversity—dominant species have less chance to be observed. In less uniform communities, the species diversity will always be lower (Melo, 2008).
Even though species diversity is not always related to conservation status, the Subtropical Rainforest of Santa Catarina deserves attention regarding conservational efforts. Our study pointed a significant amount of endemism—46.4% of the listed species occur exclusively in the Atlantic Forest and 35.8% of the species are endemic to Brazil. The Subtropical Rainforest of Santa Catarina is inserted in the Serra do Mar endemism center (Silva & Casteleti, 2003), and the aforementioned statistics seem to support this fact. Our results add dimension to the findings of Rezende, Oliveira-Filho, Eisenlohr, Kamino, and Vibrans (2014), who pointed that ∼72% of Santa Catarina’s tree species have restrict geographical distribution (see also Oliveira et al., 2016). The species records summarized in our study corresponds to 65% of the species recorded by the Forest and Floristic Inventory of Santa Catarina, a systematic regional scale survey conduct in 418 sample plots (0.4 ha) among the state (Gasper et al., 2014). Therefore, these areas are relevant for conservation because they support populations of endemic species, including those registered in the IUCN Red List (IUCN, 2003; e.g., Euterpe edulis, Ocotea catharinensis, Ocotea odorifera, and Ocotea porosa).
Implications for Conservation
Despite the fact that the species diversity of well-conserved forests may be inferior to the diversity of less conserved ones, such descriptor could be adopted for selecting priority areas for conservation in the neotropics (Gibson et al., 2011). Although, issues like those addressed in this study should be considered. In communities in which species diversity might not be the most adequate descriptor, the PDSA should be carefully analyzed. Facing the few studies regarding the Brazilian Subtropical Rain Forest, new studies, with similar approach, could concentrate efforts in investigating species diversity—especially PDSA—in different successional stages and environments, using a standardized sampling methodology to allow comparisons among sites.
Finally, we will try to conciliate the categories of less conserved and well-conserved forests, as one may ask: If the tree species diversity of a less conserved forest is greater than a well-conserved forest, is it really less conserved? The underlying issue in this question is that less conserved secondary forests could present greater diversity, yet its physiognomy, structural diversity, and even composition might not resemble a well-conserved forest, due to reasons such those presented in the description of the three categories of conservation (see Data Collection section). We still maintain a skeptic position regarding results generated by diversity indices, especially when they are employed as an isolated criterion to distinguish between different sites’ conservation status. If species diversity is a concern for defining reference areas or areas for conservation, we encourage the linking of PDSA to the ecological knowledge about the ecosystem and its historical and current aspects of resources usage. More refined approaches considering structural diversity attributes and anthropic interventions indicators might be useful, indeed (e.g., McRoberts, Winter, Chirici, & la Point, 2012; Pereira et al., 2015). Nevertheless, our findings regarding the relevant tree species diversity of secondary forests bring an encouraging perspective for future conservation.
We conclude that conservation policies should be elaborated for the Brazilian Subtropical Rain Forest remnants, taking the example of the new forest policy of Santa Catarina state, which is grounded on the protection and management of secondary forests (Vibrans & Beilfuss, 2013). Facing the expressive (∼29%) native forestland cover of Santa Catarina (Vibrans, McRoberts, Moser, & Nicoletti, 2013), areas destined for biological conservation are mandatory, and there are no reasons for having a single RBMA in the state. Secondary forests also deserve attention because of the environmental services they perform. By these reasons, we encourage the establishment of new RBMAs, which will call for attention to the importance of biological conservation as a whole.
Appendix. List of the Native Tree/Shrub Species Recorded in Different Studies Conducted in the Subtropical Rainforest of Santa Catarina, Brazil

Note. The nomenclature adopted for genera and epithets follows the Angiosperm Phylogeny Group IV (2016). Species identification (epithets) follows Flora do Brasil (http://reflora.jbrj.gov.br/). End. = endemic to Brazil; GD = geographical distribution; N = North; NE = Northeast; CW = Central West; SE = Southeast; S = South; PA = phytogeographical area; A = Amazon; CA = Caatinga; CE = Cerrado; AF = Atlantic Forest; P = Pampa; PA = Pantanal; * = lacking data for being classified.
Footnotes
Acknowledgments
We are grateful to Luiz Henrique da Silva for reviewing the English language of the manuscript.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: We are grateful to Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for the scholarship granted to the first author and to Programa de Pós-Graduação em Engenharia Florestal — Universidade Regional de Blumenau (FURB) for the financial support.