
Abstract
Objective
To comprehensively investigate the chemical composition and potential anti-inflammatory mechanisms of Nauclea officinalis in the management of chronic pneumonia.
Methods
Untargeted metabolomics was used to identify the chemical constituents of aqueous and ethanol extracts of N. officinalis. Network pharmacology and KEGG pathway enrichment analyses were performed to predict potential targets and signaling pathways associated with its anti-inflammatory activity. Molecular docking was conducted to evaluate the binding potential between representative bioactive compounds and core inflammation-related targets. In addition, an LPS-induced RAW264.7 macrophage inflammatory model was established to validate the anti-inflammatory effects of the extracts in vitro.
Results
A total of 420 chemical compounds were identified from the aqueous and ethanol extracts of N. officinalis, among which flavonoids and alkaloids were the predominant active components. Network pharmacology combined with KEGG enrichment analysis indicated that N. officinalis may exert anti-inflammatory effects through multiple targets and pathways, particularly cytokine-cytokine receptor interaction, PI3K-AKT, MAPK, NF-κB, and AGE-RAGE signaling pathways. Molecular docking suggested favorable binding between representative bioactive compounds and key inflammation-related targets, including STAT3, AKT1, IL-6, and TNF, providing supportive evidence for possible compound-target interactions. In vitro experiments showed that both aqueous and ethanol extracts of N. officinalis significantly suppressed the secretion of pro-inflammatory cytokines in LPS-stimulated RAW264.7 macrophages. Although this model represents acute inflammatory stimulation, it reflects inflammatory responses relevant to chronic pulmonary inflammatory disorders and thus provides supportive evidence for the anti-inflammatory potential of N. officinalis in chronic pneumonia.
Conclusion
Nauclea officinalis contains multiple bioactive compounds and may exert anti-inflammatory effects through multi-target and multi-pathway mechanisms. These findings provide preliminary evidence supporting its potential role in the treatment of chronic pneumonia. Further in vivo pharmacodynamic studies are needed to confirm these effects and facilitate its clinical translation.

1. Introduction
The plant Nauclea officinalis, belonging to the Rubiaceae family and the genus Nauclea, is a perennial woody plant also known as Wu Tan, Shan Xiong Dan, or Xiong Dan Shu. It typically grows in shaded, moist areas near the tops or mid-slopes of mountains and is distributed in regions such as Hainan, Guangxi, and Guangdong in China, as well as in countries like Laos, Thailand, Vietnam, Cambodia, Indonesia, and Malaysia. 1 N. officinalis is a well-known medicinal plant in southern China and has a long history of use in traditional medicine.
Nauclea officinalis has a bitter and cold nature. 2 The branches, trunk, and bark are used medicinally, and can be harvested year-round. After harvesting, they are washed, sliced, and used fresh or dried. 3 Traditionally, N. officinalis has been employed to treat conditions such as malaria, indigestion, inflammation, and wound healing. In recent years, increasing attention has been paid to its potential for managing chronic inflammatory diseases, particularly chronic pneumonia, given the plant’s rich content of bioactive compounds with proven anti-inflammatory properties. 4
Chronic pneumonia, as a persistent pulmonary inflammatory condition, contributes to ongoing respiratory symptoms and deteriorating lung function, posing a significant public health challenge. Long-standing inflammation in the respiratory tract leads to airway remodeling and recurrent pulmonary infections, underscoring the urgent need for effective, low-toxicity anti-inflammatory therapies. Due to its diverse array of bioactive constituents such as flavonoids, alkaloids, and benzene derivatives, N. officinalis has attracted interest as a candidate for managing chronic pulmonary inflammatory disorders. 5
This study conducts a comprehensive analysis of the chemical composition of ethanol and water extracts from N. officinalis using untargeted metabolomics. By integrating network pharmacology and molecular docking techniques, the research explores the anti-inflammatory pathways and potential mechanisms of N. officinalis in the disease context of chronic pneumonia. In the present study, the in vitro validation was performed using an LPS-induced macrophage inflammatory model, which reflects an acute inflammatory response relevant to the inflammatory component of chronic pneumonia. Therefore, this work provides preliminary mechanistic support for the potential application of N. officinalis in chronic pneumonia, rather than direct proof of efficacy in a chronic disease model.
2. Materials and Methods
2.1. Experimental Instruments
TripleTOF 6600+ mass spectrometer (SCIEX) coupled to a UHPLC system LC-30A (Shimadzu); refrigerated microcentrifuge 5424R (Eppendorf); temperature-controlled metal shaker MU-G02-0448 (Hangzhou MIO Instrument Co., Ltd.); analytical balance MS105DM (Mettler Toledo; readability 0.0001 g); centrifugal concentrator CentriVap (LABCONCO); vortex mixer VORTEX-5 (Kyllin-Bell); ultrasonic cleaner KQ5200E (Kunshan Instrument Co., Ltd.). The RAW 264.7 murine macrophage cell line was obtained from the Shanghai Institutes for Life Sciences, Chinese Academy of Sciences.
2.2. Experimental Materials
Methanol and acetonitrile (both HPLC grade; Merck, Cat. Nos. 1.06007.4008 and 1.00030.4008, respectively); formic acid (HPLC grade; Aladdin, Cat. No. 695076-100ML); anhydrous ethanol (Xilong Chemical, China).
2.3. Collection and Screening of N. officinalis Components
2.3.1. Preparation of Test Samples
Accurately weigh 5.0 g of N. officinalis and add 50 mL distilled water. Reflux in a water bath for 2 h, allow to cool, and filter. Transfer the filtrate to a pre-dried, constant-weight evaporating dish, evaporate on a water bath to dryness, then dry at 105 °C for 3 h. Cool in a desiccator for 30 min and weigh promptly. Dissolve the residue in an appropriate solvent to obtain a 1 mg/mL solution. Aliquot 100 μL into a 1.5 mL microcentrifuge tube, vortex for 15 min, and centrifuge at 12,000 rpm for 3 min at 4 °C. Collect the supernatant, pass through a 0.22 μm membrane filter, and transfer to an autosampler vial for UPLC–MS/MS analysis.
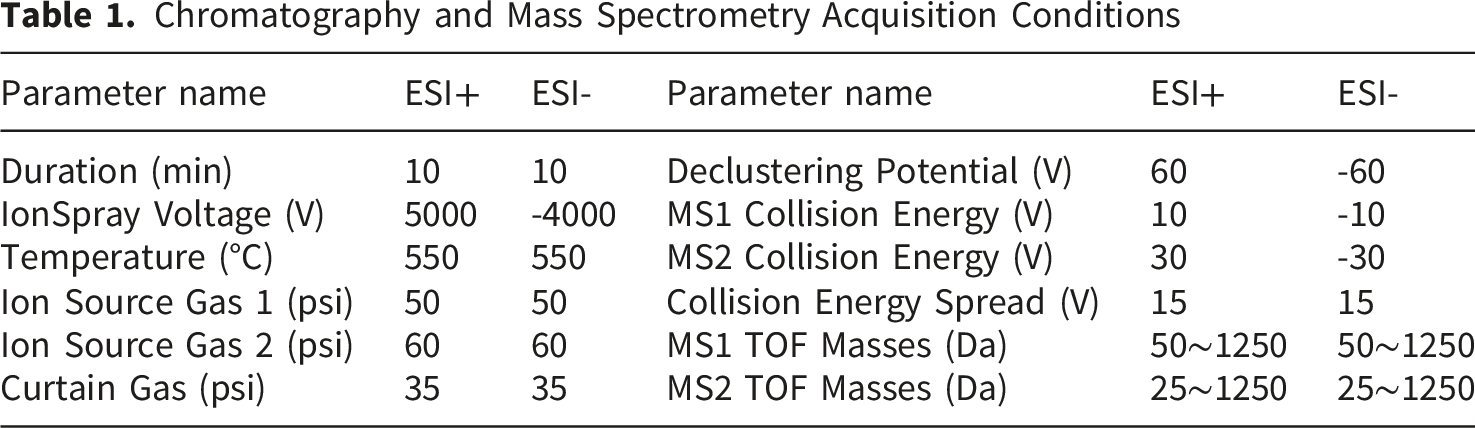
2.3.2. Chromatography-Mass Spectrometry Collection Conditions
Chromatography and Mass Spectrometry Acquisition Conditions
2.3.3. Screening of N. officinalis Chemical Components
For the detected chemical components of N. officinalis, perform retention time correction and filter out peaks with a missing rate greater than 50% in the samples. Metabolite identification is carried out by searching and integrating public databases, prediction databases, and using the metDNA method. Finally, extract substances with a comprehensive identification score greater than 0.5 and a coefficient of variation (CV) less than 0.3 in QC samples. In the present untargeted metabolomics workflow, QC samples were used for data quality assessment; however, no dedicated extraction blank or solvent blank was included for background subtraction. Merge the results from both positive and negative ion modes, retaining the substances with the highest identification level and the smallest CV value. No authentic reference standards were used in the present untargeted metabolomics workflow; therefore, the reported metabolites should be regarded as putatively annotated compounds based on database- and spectrum-matching results. Using the PubChem database (https://www.https://pubchem.ncbi.nlm.nih.gov/), 6 retrieve the SMILES codes for the active components. Then, use the SwissADME database (https://www.swissadme.ch/) to conduct an initial screening with the following criteria: iLogP ≤ 5, nOHNH ≤ 5, nOH ≤ 10, and oral bioavailability (OB) ≥ 30%. 7 Apply Lipinski’s Rule of Five to eliminate compounds that do not meet these criteria. Perform a final screening based on “High GI absorption” and “Leadlikeness” from the Medicinal Chemistry filters to identify the active components of N. officinalis. 8
2.3.4. Prediction of Active Component Targets
Putative targets for N. officinalis constituents were retrieved from SwissTargetPrediction (https://www.swisstargetprediction.ch/) and ChEMBL database (https://www.ebi.ac.uk/chembl/). 9 Target gene names were harmonized against UniProt (https://www.uniprot.org), and only entries annotated for Homo sapiens were kept. 10
2.4. Construction of Target Sets for Chronic Pneumonia
Disease-related genes were compiled from GeneCards (https://www.genecards.org/), and DisGeNet(https://www.disgenet.org/) databases to search for disease genes related to chronic pneumonia. Perform separate searches for disease targets using the keywords “chronic pneumonia”. 11
2.5. Construction and Analysis of the Protein-Protein Interaction Network
Candidate therapeutic targets were defined as the intersection between compound-associated targets and the chronic-pneumonia gene set. These were submitted to STRING (https://cn.string-db.org/) specifying Homo sapiens and a high-confidence cutoff of 0.9. Isolated nodes were removed, and the resulting network was visualized in Cytoscape v3.10.0. 12 Node importance was ranked by degree centrality to nominate core targets for N. officinalis against chronic pneumonia.
2.6. GO Functional Enrichment and KEGG Pathway Enrichment Analysis
Potential targets were analyzed for functional enrichment using Metascape (https://metascape.org/) and the DAVID database (https://david.ncifcrf.gov/). 13 Gene Ontology terms were summarized across Biological Process (BP), Cellular Component (CC), and Molecular Function (MF) categories, 14 and KEGG pathways were used to infer putative mechanisms of action. 15
2.7. Molecular Docking
Representative active compounds and core proteins were selected based on target coverage and network centrality. Ligand structures (SDF) were downloaded from PubChem; protein structures (PDB) were obtained from the RCSB PDB (https://www.rcsb.org/). 16 Docking was performed with AutoDock, and complexes were inspected in PyMOL. Binding affinity (kcal·mol-1) was used as the primary docking metric. In general, more negative binding energy values indicate more favorable predicted ligand–protein interactions. In the present study, docking scores around -6 to -7 kcal·mol-1 were interpreted as indicating acceptable to favorable binding affinity, rather than as definitive evidence of strong binding.
2.8. In Vitro Experimental Validation
RAW 264.7 (Shanghai Institute of Biochemistry and Cell Biology, Chinese Academy of Sciences, No:20250314768) murine macrophages were assigned to a blank group (basal medium), a vehicle control (medium without drug), and drug-treated groups at varying concentrations. Cell viability was assessed by the CCK-8 assay to estimate IC50. For inflammation, an LPS model was established: blank (3 mL medium), model (3 mL LPS at 1 µg·mL-1), and treatment (1 mL LPS at 1 µg·mL-1 plus 2 mL drug at 0.025, 0.05, 0.1, or 0.2 mg·mL-1). After 24 h, supernatants were collected, and TNF-α, IL-1β, and IL-6 were quantified.
Cell viability (%) = ((OD of drug group - OD of blank group)/(OD of control group - OD of blank group)) × 100
Inhibition rate (%) = ((OD of control group - OD of drug group)/(OD of control group - OD of blank group)) × 100
2.9. Statistical Analysis
All in vitro experiments were performed in triplicate, and data are presented as mean ± standard deviation (SD). Statistical analyses were performed using GraphPad Prism 9.0. Comparisons among multiple groups were conducted using one-way analysis of variance, followed by Tukey’s multiple comparison test. A value of P < 0.05 was considered statistically significant. 17
3. Results
3.1. Composition of N. officinalis Chemical Components and Confirmation of Active Compounds
Using untargeted UHPLC–Q-TOF-MS, we annotated 362 constituents in the aqueous extract of N. officinalis and 371 in the ethanolic extract (Figure 1). For the water extract, the major classes were benzene derivatives (n = 76), amino acids and related metabolites (45), flavonoids (36), alkaloids (42), organic acids (33), heterocycles (31), terpenes (14), alcohols/amines (13), phenolic acids (9), lignans/coumarins (9), and single representatives of quinones and lipids, with an additional 52 molecules grouped as “other.” The ethanol extract showed a comparable distribution: benzene derivatives (78), amino acids and derivatives (51), flavonoids (34), alkaloids (45), organic acids (32), heterocycles (30), terpenes (14), alcohols/amines (14), phenolic acids (12), lignans/coumarins (9), one quinone, one lipid, and 50 others. Composition of chemical components of N. officinalis Pierre ex Pitard
Across both matrices, 420 nonredundant compounds were compiled (Supplementary Table 1). Compounds were then filtered for oral drug-likeness and absorption using iLogP ≤ 5, nOHNH ≤ 5, nOH ≤ 10, and OB ≥ 30%, in line with Lipinski’s rule-of-five and a “High GI absorption” prediction. This yielded 102 putative actives.
Among these 102 candidates, benzene derivatives were most frequent (24), followed by alkaloids (14). To connect chemistry with disease biology, “chronic pneumonia” was used as the query term to aggregate pneumonia-related genes from multiple databases, resulting in 764 unique disease targets. Intersecting these with targets of the 102 high-absorption compounds identified 122 targets implicated in chronic pneumonia.
3.2. Construction and Analysis of the Protein-Protein Interaction Network
Based on the STRING protein interaction network, a PPI (protein-protein interaction) network was constructed for the 102 active components of N. officinalis in the treatment of chronic pneumonia, with optimization performed at the highest confidence level of 0.9.
In the PPI network for the treatment of pneumonia, there were 122 nodes and 470 edges, with an average node degree of 5.05 and an average clustering coefficient of 0.47. The top 10 core targets with the highest degree of overlap in the chronic pneumonia treatment network were: STAT3 (24), AKT1 (24), TNF (24), IL6 (22), PIK3CA (22), JUN (21), PIK3CB (20), PIK3CD (20), EGFR (17), and MAPK3 (17) (Figure 2). The PPI network of 122 intersecting target genes
3.3. Results of Gene Ontology (GO) Functional Enrichment and KEGG Pathway Enrichment
The top ten items from the gene ontology functional enrichment based on P-values were visually processed. In the molecular function analysis, Nauclea officinalis primarily involves Cytokine receptor binding (p=1.15*10-19), Phosphotransferase activity (p=1.55*10-19), Endopeptidase activity (p=6.61*10-18), Transcription factor binding (p=2.95*10-16), Chemokine binding (p=7.41*10-15). Regarding biological processes, it mainly affects the Positive regulation of locomotion (p=2.95*10-47), Response to molecule of bacterial origin (p=2.00*10-40), Positive regulation of response to external stimulus (p=2.82*10-40), Cellular response to cytokine stimulus (p=1.62*10-39), Response to hormone (p=1.12*10-35). In cellular components, it primarily relates to the External side of plasma membrane (p=2.75*10-21), Membrane raft (p=2.45*10-15), Endocytic vesicle (p=1.32*10-11), Cell leading edge (p=1.86*10-11), Lytic vacuole (p=2.75*10-11) (Figure 3). Gene Ontology (GO) functional enrichment and KEGG pathway enrichment analysis (A): GO Molecular Functions; (B): GO Cellular Components; (C): GO Biological Processec; (D): KEGG Pathway
In the shared target genes for chronic pneumonia, 29 target genes are directly involved in the AGE-RAGE signaling pathway, with the top four in terms of overlap being AKT1 (15), JUN (13), IL6 (12), and TNF (12) as listed in Table 1. Within the PPI network constructed from these 29 target genes, there are 29 nodes and 307 edges, with an average node degree of 21.2 and an average clustering coefficient of 0.849 (Figure 4). The PPI network of 29 intersecting target genes
3.4. Molecular Docking Analysis
Active Ingredient and Core Target Docking Information


Schematic of molecular docking of N. officinalis Pierre ex Pitard (A): AKT1 with two compounds; (B): AKT2 with two compounds
3.5. Effects of N. officinalis Extracts on RAW264.7 Cells
The cytotoxic effects of different concentrations of N. officinalis extracts on RAW264.7 cells were reflected through cell viability. As shown in Figure 6A, when the water extract concentration of N. officinalis ranged from 0.0125 to 0.05 mg/mL, the cell viability was between 103.40% and 105.38%, indicating a significant proliferative effect. At concentrations of 0.1 and 0.2 mg/mL, the cell viability decreased to 97.52% and 88.11%, respectively, suggesting no apparent toxicity in this range. However, at a concentration of 1.25 mg/mL, the cell viability dropped to 53.80%, indicating toxic effects at this concentration. In contrast, when the ethanol extract concentration of N. officinalis ranged from 0.0125 to 0.2 mg/mL (Figure 6B), the cell viability was between 76.61% and 95.10%, showing no significant toxicity. At a concentration of 1.25 mg/mL, the cell viability reduced to 53.16%, indicating toxicity at this concentration. Therefore, concentrations of 0.025, 0.05, 0.1, and 0.2 mg/mL of N. officinalis extracts were selected for subsequent studies (Figure 6). Effect of different concentrations of drugs on the proliferation of RAW264.7 cells (P<0.05, n=3) (A): Water extract cell viability; (B): Alcoholic extract cell viability
Several important findings were observed in the enzyme-linked immunosorbent assay (ELISA). A comparison between the control and model groups revealed that, upon lipopolysaccharide (LPS) stimulation, the levels of pro-inflammatory cytokines, such as TNF-α, IL-6, and IL-1β, were significantly higher in the model group than in the control group. Further experimental results showed that, compared to the model group, the water and ethanol extracts of N. officinalis significantly reduced the expression levels of these three inflammatory cytokines in treated cells (p < 0.05). Specifically, at a concentration of 0.2 mg/mL, the water extract of N. officinalis reduced the expression levels of these cytokines by 27.91%, 62.93%, and 33.85%, respectively, while the ethanol extract reduced them by 28.87%, 58.16%, and 51.61% (Figure 7). Inhibitory effects of N. officinalis on cytokine release by cells (P < 0.05, n = 3) (A): Inhibitory effect of N. officinalis on TNF-α release by cells; (B): Inhibitory effect of N. officinalis on IL-6 release by cells; (C): Inhibitory effect of N. officinalis on IL-1β release by cells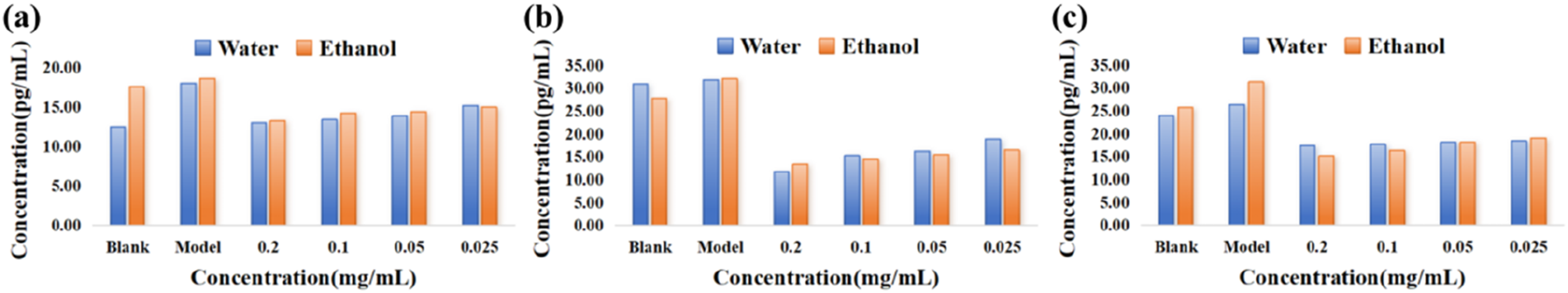
4. Discussion
4.1. Chemical Constituents of Nauclea officinalis and Its Mechanism in the Treatment of Chronic Pneumonia
In this study, untargeted metabolomics combined with network pharmacology and molecular docking approaches was employed to systematically analyze the chemical constituents of Nauclea officinalis and investigate its potential mechanisms in the treatment of chronic pneumonia. A total of 420 chemical compounds were identified from both aqueous and ethanol extracts of N. officinalis, primarily including flavonoids, alkaloids, benzene derivatives, organic acids, and terpenoids, with flavonoids and alkaloids being the most abundant categories. These compounds have been widely documented in previous studies for their anti-inflammatory, antioxidant, and immunomodulatory properties.
Network pharmacology analysis identified 186 potential targets of N. officinalis related to chronic pneumonia, with STAT3, AKT1, TNF, IL-6, and JUN recognized as key hub targets. GO and KEGG enrichment analyses indicated that these targets are predominantly involved in cytokine–cytokine receptor interaction, inflammatory mediator regulation of TRP channels, PI3K-AKT, MAPK, and NF-κB signaling pathways, all of which are closely associated with the progression of chronic pulmonary inflammation.18,19
Molecular docking results demonstrated favorable binding affinities between several major active compounds and inflammation-related proteins such as STAT3, AKT1, and TNF, suggesting the possibility of compound–target interactions relevant to anti-inflammatory activity. However, these docking results alone are insufficient to demonstrate direct interference with specific signaling cascades. 20 In vitro experiments using an LPS-induced RAW264.7 macrophage model confirmed that both aqueous and ethanol extracts of N. officinalis significantly reduced the secretion of pro-inflammatory cytokines, including TNF-α, IL-6, and IL-1β, while maintaining good cellular safety at effective concentrations. 21 Collectively, these findings preliminarily demonstrate that N. officinalis exerts therapeutic effects against chronic pneumonia through a multi-component, multi-target, and multi-pathway synergistic mechanism, modulating inflammatory cytokines and suppressing pulmonary inflammatory signaling activity. 22
4.2. The Role of the AGE-RAGE Signaling Pathway in Inflammatory Responses and the Intervention Mechanism of N. officinalis
The AGE-RAGE signaling pathway has been increasingly recognized as a key regulatory axis in the development of chronic inflammation-related diseases. Advanced glycation end-products (AGEs), formed through non-enzymatic glycation of sugars, lipids, or proteins under oxidative stress, tend to accumulate in chronic inflammatory environments. The binding of AGEs to their receptor RAGE activates downstream signaling cascades including NF-κB, MAPK, PI3K-AKT, and AP-1, which subsequently induce the overexpression of pro-inflammatory cytokines such as TNF-α, IL-6, and IL-1β, ultimately leading to sustained local inflammation and tissue damage, thereby forming a persistent positive feedback loop of chronic inflammation. 19
In the pathogenesis of chronic pneumonia, prolonged airway infections and oxidative stress accelerate the deposition of AGEs in lung tissues. Persistent activation of RAGE-mediated inflammatory signaling exacerbates airway remodeling, alveolar wall thickening, and fibrotic progression, contributing to chronic obstructive pulmonary disease (COPD) and declining lung function. Thus, disrupting the AGE-RAGE signaling axis is considered an important therapeutic strategy for managing chronic pulmonary inflammation.
In this study, network pharmacology and KEGG pathway enrichment analyses revealed that several active components in N. officinalis were highly enriched in targets associated with the AGE-RAGE signaling pathway, notably AKT1, JUN, IL-6, and TNF. Molecular docking suggested that representative flavonoids, alkaloids, and benzene derivatives may bind favorably to several proteins associated with the AGE-RAGE-related inflammatory network. These results provide supportive evidence for possible compound–target interactions, but do not directly demonstrate that the AGE-RAGE signaling cascade is functionally modulated (Figure 8).
23
However, the present study did not include direct pathway-validation experiments, such as Western blot analysis of p-AKT, NF-κB p65, or RAGE expression, qPCR analysis of inflammation-related genes, intracellular ROS measurement, or inhibitor-based verification using a RAGE antagonist. Therefore, the involvement of the AGE-RAGE/PI3K-AKT/NF-κB signaling axis should still be regarded as a mechanistic hypothesis requiring further experimental confirmation. AGE-RAGE signalling pathway
In vitro experiments using LPS-induced RAW264.7 macrophages demonstrated that N. officinalis extracts significantly reduced the expression of key pro-inflammatory cytokines TNF-α, IL-6, and IL-1β associated with the AGE-RAGE pathway. These results preliminarily confirm that N. officinalis can alleviate macrophage-mediated inflammatory responses by inhibiting the activity of the AGE-RAGE axis and modulating the release of inflammatory cytokines, providing a promising new approach for the treatment of chronic pulmonary inflammation. 24
Further predictive analysis indicates that, beyond directly blocking RAGE-mediated inflammatory signaling, the active components of N. officinalis may also regulate AGEs production, reduce oxidative stress, and enhance endogenous antioxidant defense systems, indirectly reducing AGEs accumulation in lung tissues and mitigating continuous RAGE activation. This multi-target, multi-pathway, and multi-step intervention mechanism offers substantial potential for the long-term management of chronic pneumonia through safe and effective plant-derived therapeutics. Future studies employing in vivo chronic pneumonia animal models will be conducted to validate these mechanisms and comprehensively evaluate the effects of N. officinalis on AGEs formation, RAGE expression, oxidative stress, and pulmonary histopathological improvement, further expanding its application prospects in functional herbal beverages and botanical drug preparations for chronic respiratory diseases. 25
4.3. Study Limitations and Prospects for Functional Beverage Applications
Although this study systematically analyzed the chemical constituents of N. officinalis and explored its potential anti-inflammatory mechanisms in the context of chronic pneumonia, several limitations should be acknowledged. First, the present work was mainly based on computational prediction and in vitro experiments, without in vivo validation. Second, the cellular validation was performed using an LPS-induced RAW264.7 macrophage model, which represents an acute inflammatory stimulation model rather than a bona fide chronic pneumonia model. Therefore, the current findings should be interpreted as preliminary evidence of anti-inflammatory activity relevant to chronic pneumonia, rather than direct proof of efficacy in a chronic disease setting. Further in vivo pharmacodynamic, safety, and pharmacokinetic studies are needed to validate its therapeutic relevance.
Despite these limitations, the identified anti-inflammatory activity and bioactive compound profile of N. officinalis provide preliminary support for its future development in respiratory health-related botanical products, including functional herbal beverages.
5. Conclusions
This study comprehensively explored the chemical composition and potential anti-inflammatory mechanisms of Nauclea officinalis in the treatment of chronic pneumonia through untargeted metabolomics, network pharmacology, molecular docking, and in vitro validation. A total of 420 chemical compounds were identified from both aqueous and ethanol extracts, with flavonoids and alkaloids being the most abundant and pharmacologically significant.
Network pharmacology and KEGG enrichment analyses suggested that N. officinalis may exert anti-inflammatory effects via multi-target and multi-pathway mechanisms, potentially involving cytokine–cytokine receptor interactions, PI3K-AKT, MAPK, NF-κB, and the AGE-RAGE signaling pathway. Molecular docking provided supportive evidence for favorable interactions between major active compounds and several core inflammation-related proteins. However, the present study did not directly verify modulation of the AGE-RAGE pathway at the molecular level.
These findings provide a scientific basis for the traditional use of N. officinalis in respiratory health and highlight its potential as a natural therapeutic agent for the management of chronic pulmonary inflammation. Moreover, the study offers valuable theoretical support for the development of N. officinalis-based functional herbal beverages targeting chronic pneumonia. Future studies will focus on in vivo pharmacodynamic validation and comprehensive pharmacokinetic evaluations to further confirm its efficacy and clinical application potential.
Supplemental Material
Supplemental material - Chemical Components and Anti-Inflammatory Activity of Nauclea officinalis Branches: A Multidimensional Analysis Through Bioinformatics, Metabolomics, and In Vitro Experiments
Supplemental material for Chemical Components and Anti-Inflammatory Activity of Nauclea officinalis Branches: A Multidimensional Analysis Through Bioinformatics, Metabolomics, and In Vitro Experiments by Xueying Lin, Guifei Chen, Zixiao Jiang, Dandan Chen, Sakda Daduang, Arunrat Chaveerach, and Yangyang Liu in Natural Product Communications.
Footnotes
Consent for Publication
All authors have read and agreed to the published version of the manuscript.
Author Contributions
Conceptualization, Z.X. J. and Y.Y. L.; methodology, Z.X. J.; software, X.Y. L.; validation, X.Y. L. and G.F. C.; formal analysis, Z.X. J. and D.D. C.; investigation, Z.X. J.; resources, X.Y. L. and G.F. C.; data curation, X.Y. L. and S. D.; writing—original draft preparation, X.Y. L., Z.X. J., and G.F. C.; writing—review and editing, Y.Y. L., and A. C.; visualization, X.Y. L. and G.F. C.; supervision, X.Y. L. and G.F. C.; project administration, Y.Y. L.; funding acquisition, Y.Y. L. The authors confirm that no paper mill and artificial intelligence was used.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was funded by the Hainan Provincial Joint Project for Health and Medical Science and Technology Innovation (grant number: WSJK2024QN085); Hainan Provincial Natural Science Foundation of China (grant number: 825QN562); Hainan Province Science and Technology Special Fund (grant number: ZDYF2025SXLH015); Hainan Provincial Key Research and Development Project (grant number: ZDYF2026SHFZ065); Hainan Province Nanhai New Star Science and Technology Innovation Talent Program (grant number: NHXXKJCX202531).
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.