
Abstract
Introduction
Dental caries, one of the most prevalent infectious diseases in the world, is a biofilm-mediated, sugar-driven, multifactorial dynamic disease. 1 Biofilm is a single or multispecies microbial community attached to the surface of teeth and wrapped in the self-produced matrix. 2 The formation of dental plaque biofilm is one of the important causes of dental caries. 3 The biofilm formation process will produce anaerobic and acidic environment, leading to the formation and development of dental caries. 4 Therefore, the eradication of dental plaque biofilm and the control of acid production by bacteria are effective ways to control dental caries. 5 Streptococcus mutans (S mutans) is an important cariogenic bacterium in the oral cavity, whose cariogenic factors are adhesion, production of polysaccharides, strong acid resistance, and acid production.6,7 Existing medicines, such as fluoride and chemical agents, have preventive effects on dental caries, but long-term use is associated with toxicity and strong side effects.8–14 Therefore, finding a safe and effective antimicrobial agent to inhibit the formation of biofilm in the basal phase of bacterial growth has always been a hot topic in the prevention and treatment of dental caries.15–17
Natural products, especially plant essential oils, are suitable leads for antimicrobial drugs that can inhibit bacterial biofilms. Cistus munbyi essential oil made Pseudomonas aeruginosa biofilm reduce significantly, with distorted architecture and cell shrinkage. 18 Areca nut essential oils prevented S mutans from forming biofilms in multiple ways, such as decreasing the surface hydrophobicity, inhibiting glucosyltransferase activity and so on. 19 Essential oils have a variety of physiological activities and are expected to be a source of new drugs against dental caries, which have received extensive attention.19–22 Oregano essential oil (OEO) has been proven to have good antibacterial activity.23–27 OEO has a good antibacterial effect on common infectious disease pathogens, including Candida albicans, Escherichia coli, Staphylococcus aureus, Candida krusei, Salmonella enterica, Serratia marcescens, and so on.28,29 However, the antibacterial mechanism of OEO against S mutans has not been thoroughly studied, especially its inhibition and removal mechanism of biofilm in anti-caries has not been clearly elucidated. The objectives of this study were to explore the antibacterial activity of OEO against S mutans in vitro and to evaluate its antibacterial activity against planktonic S mutans and its biofilm inhibition and clearance activities against S mutans during dental caries formation, which provides new insights into reducing caries occurrence and solving the problem of drug resistance in S mutans. At the same time, the physiological toxicity of OEO on oral cells was studied, which laid a theoretical foundation for the development of OEO as an anti-caries drug.
Materials and Methods
Bacterial Strain and Growth Condition
Streptococcus mutans (ATCC 25175) was obtained from the Guangdong Institute of Microbiology. S mutans was grown in brain heart infusion agar (BHI, Guangdong Huankai Microbial Sci. & Tech. Co., Ltd) at 37 °C for 24 to 48 h. The OEO (Origanum vulgare L.) was provided by Xiangsi Xinqing Health Technology Co., Ld.
Determination of Minimum Inhibitory Concentration (MIC)
The MIC was calculated in triplicate on 96-well microtiter plates using the broth-based microdilution technique described before. 30 In a 96-well microtiter plate, 10 stages of 2-fold successive dilutions of the OEO were carried out before sucrose-supplemented BHI medium had been added to the wells. The microtiter dishes were then inoculated with a predetermined amount of cultured bacteria and left to grow for 24 h.
Growth Curves Assay
As previously published, 31 a growth curves experiment was performed to investigate S mutans susceptibility to the OEO surrounding the MIC threshold. S mutans was grown overnight in a 96-well microplate, as well as OEO was added to reach final different concentrations (0.0625-0.5 mg/mL). As a vehicle control, a suspension of bacteria without OEO was implemented. A Microbiology Growth Curves Analyzer (FP-1100-C, Bioscreen, Finland) was used to track bacterial growth. At 30-minute intervals, an optical density (OD) in the 600 nm wavelength of each well was recorded.
Bacteria Colony Forming Units Count
After varied incubation periods (0, 12, and 24 h) with the OEO ranging from 0.0625 to 0.5 mg/mL, the bacteria colony-forming unit (CFU) was determined. A total of 100 μL of the bacterial sample was grown onto BHI agar plates for a further 2 days using 10-fold successive dilutions of both untreated and treated samples. CFUs were determined and recorded, to ascertain the amount of viable bacteria under the bactericidal action of the OEO.
Fluorescence Staining
The bacterial membrane integrity was assessed by 4′,6-diamidino-2-phenylindole (DAPI), and Nile Red (NR) staining. S mutans were prepared into bacterial suspensions. When NR is paired with OEO as a marker, the characteristic red fluorescence of the planktonic bacteria can be viewed under a fluorescence microscope. The suspensions (1 × 107 CFU/mL) were initially treated for 1 h at 37 °C with 0.25 mg/mL OEO. The bacteria were rinsed 3 times with a 0.9% solution of sodium chloride before being fixed for 1 h at 4 °C in 4% paraformaldehyde. Then, 200 μL of DAPI (10 mg/mL) and 200 μL of NR (6 mg/mL) were added, followed by 30 min of incubation in darkness at the ambient temperature. The bacteria were viewed under an inverted fluorescent microscope (Olympus IX71) after being washed with 0.9% NaCl solution. PBS-treated bacteria acted as a control.
Transmission Electron Microscopy (TEM)
TEM was used to explore the influence of OEO on bacterium cellular morphology, as reported by Duan et al. 32 Regarding the mature biofilm experiment, confocal dishes (BS-20-GJM, Biosharp) were first filled with overnight cultured bacteria diluted in BHI broth, and the plates were subsequently left to incubate at 37 °C for 24 h in order to generate mature biofilms. In brief, OEO was introduced into the BHI medium in increments of 0.0625 to 0.5 mg/mL for 24 h at 37 °C. As controls, groups with no OEO were employed. The microorganisms were gathered in a 1.5 mL centrifuge container and fixed for 6 h at 4 °C in 200 to 400 μL of 2.5% (v/v) glutaraldehyde. Following fixation, samples were washed 3 times with PBS and centrifugation after each wash. The supernatant was eventually removed after being centrifuged (8000 r/min for 3 min). Gradient dehydration was performed for 10 min using ethanol concentrations of 30%, 60%, 70%, 80%, 90%, and 100% (v/v). Subsequently, the mesh copper grid was filled with 2 or 3 drops of the suspensions and allowed to dry. Using a field emission electron microscope with transmission (Talos F200S, Fei), the sample morphologies were studied.
Scanning Electron Microscopy (SEM) Analysis
Following the method presented, 33 SEM was used to study the physical characteristics of S mutans biofilm underneath the influence of OEO. Mature biofilms were grown as described in the above section. In a dish, glass coverslips were put with a diluted suspension of S mutans (105 CFU/mL). The OEO had been added into each well at concentrations ranging from 0.0625 to 0.5 mg/mL. BS was included as an adverse control. The cell culture dish was then left to incubate for 24 h at 37 °C. After removing the excess media, 2.5% (v/v) of the substance was added to the plate, which was then stored at 4 °C for 12 h. The plate was subsequently treated with successive variations in ethanol (30%, 40%, 60%, 70%, 80%, 90%, and 100%, v/v) for 15 min to dehydrate the biomass at each concentration. The coverslips were examined using SEM (LYRA 3 XMU, Teascan) after gold sputtering.
Confocal Laser Scanning Microscope (CLSM) Analysis
The impacts of OEO on constructed S mutans biofilm were evaluated using a CLSM using the method previously outlined. 34 Mature biofilms were grown as described in the above section. Biofilm bacteria were stained by using the LIVE/DEAD Bacterial Dual Stain Kit (MX4234-40 T, Maokang Biotechnology Co., Ltd), which consists of the nucleic acid dyes SYTO 9 and propidium iodide (PI). SYTO 9 accesses both vital and dead cells and displays green fluorescence, whereas PI enters only nonviable cells with damaged membranes, exhibits yellow-red fluorescence which and satisfies the fluorescence generated by SYTO 9. Images were acquired using an LSM 800 armed with Airscan (Carl Zeiss) and dual-channel scanner observation. The micro-images and biofilm quantification findings were evaluated using the Zeiss Zen confocal laser scanning microscope software. For additional investigation, 3-dimensional in-nature reconstruction projections were created.
Crystal Violet Assay
Following biofilm development, the biomass of the biofilm was estimated using crystal violet staining. 35 Regarding the mature biofilm experiment, 24-well plates were first filled with overnight cultured bacteria diluted in BHI broth (Guangdong Huankai Microbial Sci. & Tech. Co., Ltd), and the plates were subsequently left to incubate at 37 °C for 24 h in order to generate mature biofilms. In short, the biofilm was allowed to air dry and the supernatant was disposed of after the biofilm was fixed on 24-well plates using methanol for 15 min. After that, each well received 100 μL of 0.1% crystal violet, which was then incubated. The biofilm was then washed with sterilized ionized water and left to air dry. In order to do a quantitative study, the biofilm's crystal violet stain was dispersed in a 33% acetic acid solution, and the absorbance at 595 nm was quantified by a Flash microplate reader (SuPerMax 3100).
Bacteria Metabolic Assay
The MTT (methyl thiazolyl tetrazolium) assay is effective for determining bacterial metabolic activity by using the colorimetric technique. Mature biofilms were grown as described in the above section. The procedure was carried out as previously reported, with various adjustments. 36 Following OEO incubation, the growth medium was gently removed, washed 3 times with sterile PBS, and replaced with 100 L of MTT (0.5 mg/mL, Biosharp) for 3 h in the dark. After washing the wells with PBS, the tetrazolium precipitates in the biofilm were extracted using 150 L dimethylsulfoxide (DMSO). The absorbance of the sample was determined at 450 nm using visible spectrophotometers (721, Shanghai Jinghua Technology Instrument Co., Ltd) after 10 min on an orbital shaker.
Extracellular Polysaccharides (EPS) Production Assay
The approach used to assess the influence of the OEO on EPS was described in the published research. 37 Mature biofilms were grown as described in the above section. In brief, 100 L of the suspension of bacteria and 1 mL BHI solution were combined and cultivated for 24 h at 37 °C with or without various doses of the OEO ranging from 0.0625 to 0.5 mg/mL. The supernatant dissolved in NaOH (0.1 M) was collected by centrifugation. These supernatants were mixed, and overnight at 4 °C, on 3 separate occasions, a volume of 95% ethanol was added. The mixture was centrifuged for 30 min at 4 °C at 12 000×g, and the resultant precipitate was insoluble in water EPS. To assess the content of the precipitate, it was dissolved in NaOH. The phenol-sulfuric acid technique was used to detect the concentration of water-insoluble EPS at the wavelength of 492 nm by an ultraviolet-visible (UV–vis) spectrophotometer (TU-1950, Purkinje General Instrument, Beijing).
Acid Production Measurement
The pH measurement represents the capacity of the S mutans biofilm to produce acid. 38 In brief, 100 L of microbial suspensions and 1 mL BHI culture were combined and cultivated for 24 h at 37 °C with or without a variety of doses of OEO ranging from 0.0625 to 0.5 mg/mL. A pH meter (PHS-3C, REX) was used to test the pH of the growing medium.
Extracellular DNA Isolation and Interactions
Since EPS and eDNA are crucial for bacterial biofilm defense, the interaction of OEO with eDNA is a useful technique for breaking down mature biofilm. eDNA was isolated using a procedure described in a published study. 39 Sonication is in a bath of ice water for 5 min dispersed the mature S mutans biofilm, followed by removal using a 0.22 m filter to remove the bacteria. Following that, the cell-free supernatant was combined with 1/10 of a volume of sodium acetate solution (3 M, pH 5.2) and 2 volumes of 100% ethanol. The eDNA was extracted by centrifugation (1200×g, 4 °C, 10 min) and dissolved in sterile water before measuring it utilizing an ultramicro spectrophotometer (TY-WLF900). The extracted eDNA (5 mg/mL, 1 mL) was stained with ethidium bromide (EB, 1 mg/mL) and incubated at room temperature for 10 min. The fluorescence intensity of EB was evaluated with a microplate reader (SuPerMax 3100, Flash) with eDNA and the OEO. The wavelength of excitation was set to 480 nm, while the size of the slit was set to 10 nm. The emitted fluorescence was measured between 520 and 780 nm.
Chemical Components
Using a mass spectrometer (5977B, Agilent) and gas chromatography (7890A, Agilent), the chemical composition of OEO was examined using the methodology outlined. 40 The following were the chromatographic conditions: The temperature program ramps from 60 °C to 300 °C with a gradient of 5 °C/min (holding the beginning and final temperatures for 5 min); the injector and detector temperatures are 260 °C and 280 °C, respectively. The sample preparation is 1 mL in 1 mL of ethanol; the injection volume is 0.4 μL; the split ratio is 120:1; the helium flow rate is 1.6 mL/min. By comparing volatile molecules’ retention indices (RI) to those of n-alkanes (C6-C19), volatile chemicals were found. To qualitatively examine unknown substances, the chemical workstation's NIST 14 standard mass spectrometry library was searched for the spectrum peak. Peak area normalization was used to compute each component's relative content.
Cytotoxicity
The cytotoxicity of OEO on Human Oral Epithelial Cells (HOEC, NEWGAINBIO) was determined using the CCK-8 assay according to the manufacturer's directions (Cell-Counting-Kit 8, Biosharp). The cells were grown in DMEM (Gibco, USA) with 15% FBS (Excell Bio, Uruguay) at 37 °C in 5% CO2. The cells were grown at a concentration of 5000 cells per well on a 96-well plate. The OEO concentrations ranged from 0.0625 to 0.5 mg/mL. The group serving as the negative control received the same supply of PBS as the positive control group. After 1 h of incubation at 37 °C in 5% CO2, the cells were rinsed with sanitized PBS, and CCK-8 solution was applied to each well. After 1 h of incubation at 37 °C, the absorbance at 450 nm was measured with a reader for micro-plates (Labserv K3, Fisher Scientific). The OD of pretreated control cells was deemed to represent 100% vitality.
Statistical Analysis
Three duplicates of each experiment were run, and the results were reported as means ± deviations from the average. The IBM SPSS 25.0 software (Armonk, NY) was used for the statistical analyses. One-way ANOVA with Waller-Duncan's multiple range test (P < .05) was used to look at group differences.
Results
Antibacterial Activity
When compared to the control group, the OEO demonstrated substantial inhibition at doses > 0.125 mg/mL and dose-dependent antibacterial action against S mutans (P < .05). With a MIC of 0.25 mg/mL, the antibacterial capacity of the OEO versus S mutans was displayed. 41 The results of the growth curve test showed that OEO significantly slowed down the pace at which S mutans developed in comparison to the carrier control group at doses of 0.0625 mg/mL or higher concentrations (Figure 1). We collected the colony-forming units at various times throughout the 24-h incubation to further measure the growth-inhibitory impact on the OEO's planktonic bacteria (Figure 1B). Bacteria revealed red fluorescence caused by the attached NR after 3 min of incubation with NR-OEO (Figure 1C). It showed that when the OEO was added to the bacterial solution, it quickly bound to the outer layer of S mutans. After incubating the bacteria with NR-OEO for 40 min, a stable platform in fluorescence density was attained (Figure 1C). Because of this quick contact, the OEO successfully inhibited bacterial growth. As a consequence, the bacteria clumped together, limiting their adherence to the underlying surface. Furthermore, the OEO damaged the cell membranes from the inside, causing the bacteria to drop drastically, as seen by fluorescent microscope imaging (Figure 1D).
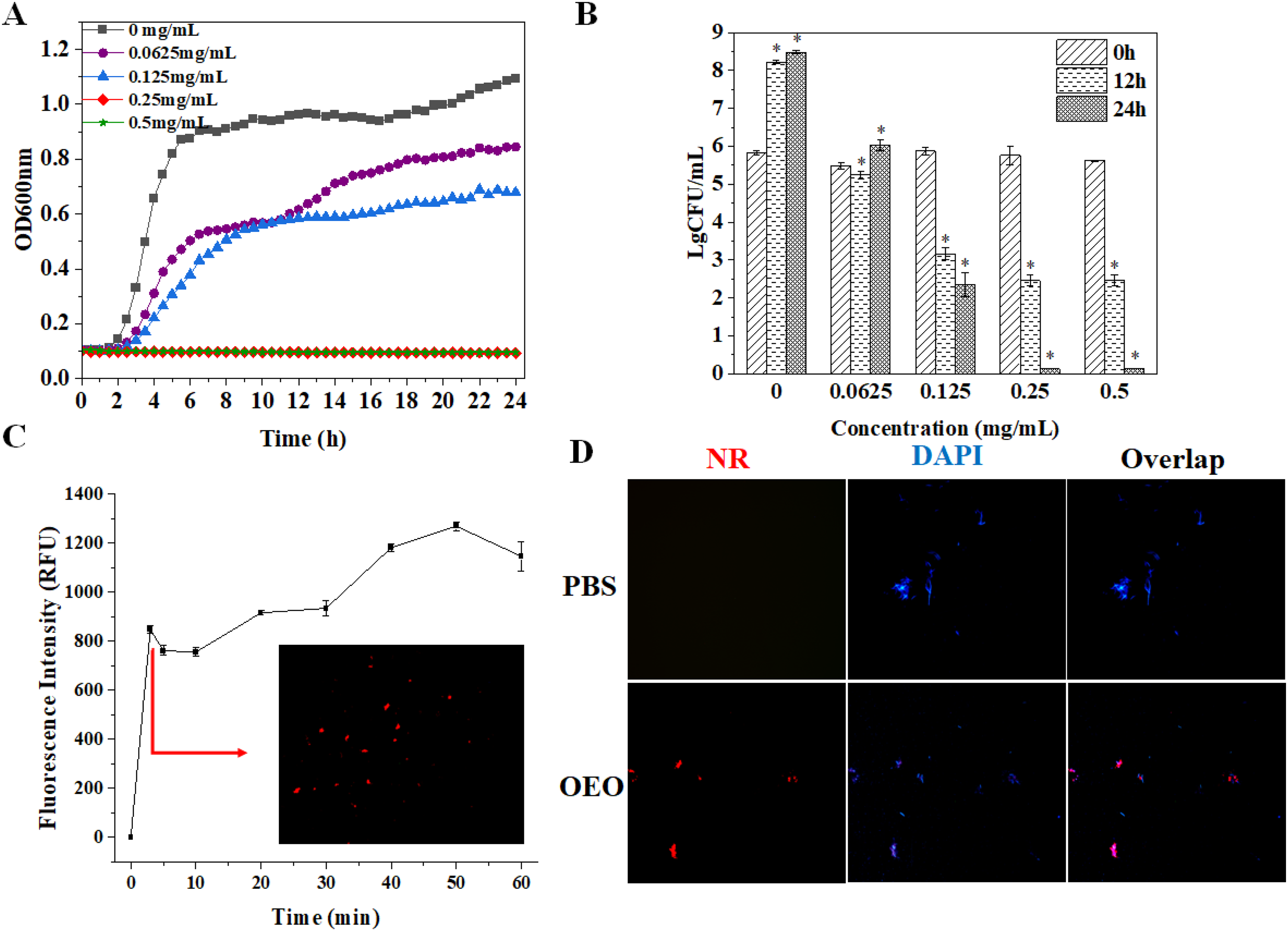
Antibacterial activity with different concentrations of OEO: (A) growth curves; (B) CFU of S mutans; (C) fluorescence microscopy pictures of S mutans following a one-hour incubation period at 37 °C, with PBS and 0.25 mg/mL OEO present. NR and DAPI were used to stain the bacteria to perform fluorescence microscopy; (D) S mutans (1 × 107 CFU/mL) fluorescence intensity following 0.25 mg/mL NR-OEO incubation for the specified periods of time. After 3 min of NR-OEO incubation, fluorescence microscopy pictures of S mutans was displayed in the inset.
Transmission Electron Microscopy
Figure 2A demonstrates that without the OEO treatment, the cell surface of S mutans appeared smooth, the cells were plump, undamaged, and free of damage, and the contents were homogeneous and dense. The outer layer of the S mutans became wrinkled and shrunken after treatment with OEO at 0.0625 to 0.5 mg/mL. Figure 2C illustrates the preliminary deterioration of the cell wall, which is seen more clearly in Figure 2D and E. Therefore, the OEO treatment acted on S mutans’ cell wall to cause morphological changes in the cell's outermost layer and disrupt the cell membrane, resulting in intracellular contents, causing bacterial cell death.

Transmission electron microscopy (TEM) micrograph of S mutans: (A) 0 mg/mL, (B) treated with 0.0625 mg/mL, (C) treated with 0.125 mg/mL, (D) treated with 0.25 mg/mL, and (E) treated with 0.5 mg/mL.
Scanning Electron Microscopy
SEM was used to assess how the OEO affected the amounts of bacteria present and the surface shape. The biofilm produced by S mutans aggregation is shown in Figure 3A. Bacteria exposed to 0.125 mg/mL OEO also showed noticeable structural changes, which were more severe at increasing concentrations. In particular, the bacteria treated with the OEO exhibited irregular oval forms with surfaces that were rough, wrinkled, dented, and distorted; also, the quantity of bacteria drastically dropped at 0.5 mg/mL OEO.

Scanning electron microscopy (SEM) micrograph depicting the effect of the OEO on biofilm formation: (A) 0 mg/mL, (B) 0.0625 mg/mL, (C) 0.125 mg/mL, (D) 0.25 mg/mL, and (E) 0.5 mg/mL.
Confocal Laser Scanning Microscope
As shown in Figure 4, a series of dimensional micrographs were created by merging the pictures using Zeiss Zen software. The living bacteria were colored green and dead organisms were labeled red to further ascertain the impact of the OEO on mature biofilm. The CLSM analysis supported the experimental findings on the OEO's ability to suppress biofilm formation, as shown in Figure 4. When the quantity of OEO was increased from 0.0625 to 0.5 mg/mL, the amounts of dead bacteria in the groups that were treated increased significantly in comparison to the control group. Additionally, as indicated in Figure 4F, the percentage of living and dead bacteria in the S mutans the biofilms on the monocular dish was computed from the bottom to the top. The architectural analysis revealed a roughly 62% decrease in the number of biofilm bacteria in the untreated and 0.5 mg/mL treated biofilms.

The CLSM images of S mutans biofilm treated with the OEO concentration of (A) 0 mg/mL, (B) 0.0625 mg/mL, (C) 0.125 mg/mL, (D) 0.25 mg/mL, and (E) 0.5 mg/mL. The green and red colors indicated live and dead bacteria, respectively. (F) Image analysis shows the proportion of viable cells in the biofilm formed with different concentrations of the OEO treatment.
EPS Production and pH Decrease
Figure 5A depicts the inhibitory impact of OEO on water-insoluble EPS. In comparison to the control group, the water-insoluble EPS produced by S mutans was reduced by 14.27% to 80.56% with OEO treatments, from 0.0625 to 0.5 mg/mL. The pH decrease under the OEO treatment was also quantified. The pH rose from 5.40 ± 0.01 to 7.35 ± 0.01 as the OEO concentration increased, as shown in Figure 5B. Thus, there was no discernible decrease in pH among the groups with increasing concentrations of OEO as its concentration rose. There was a discernible pH reduction, which indicated increased acid generation. These showed that the OEO successfully prevented the pH decline at concentrations up to 0.25 mg/mL.

The effect of OEO on S. mutans after 24 h incubation, (A) Water-insoluble EPS production. (B) pH value. Data represent the mean ± standard deviation of 3 independent tests performed in triplicate.
Oregano Essential Oil of Anti-Biofilm Mechanism Analysis
The developed biofilm by S mutans is measured (Figure 6) at OEO concentrations ranging from 0.0625 to 0.5 mg/mL. There was a significant difference in the total biomass of biofilm between the OEO treatment groups and the control group. During the 24-hour incubation period, the biofilm mass decreased as the concentration of OEO rose (Figure 6A and C). Figure 6A depicts OEO's inhibitory impact on biofilm development, whereas Figure 6C depicts OEO's destructive effect on mature biofilms. Furthermore, when the concentration of OEO grew, the rate of metabolism of the biofilm decreased significantly (Figure 6B). The amount of metabolism involved in the biofilm development was reduced by the OEO (Figure 6B), and the proportion of bacteria with damaged membranes increased (Figure 6D) while the thickness of the mature biofilm decreased. These findings showed that the OEO destroyed bacteria, prevented the production of biofilms, and eliminated mature biofilms. Apart from the aforementioned modifications in the mass of biofilms, bacterial survival ratio, extracellular matrix synthesis, and acid generation, fluorescence studies verified that the connections between OEO and eDNA were critical for inhibiting biofilm formation and removing biofilm. When EB bound to the duplexes portion of DNA, it produced a high fluorescence increase at 580 to 630 nm, making it a regularly used fluorescent marker for nucleic acids. When EB was attached to eDNA, a hyperchromic reaction occurred, as shown in Figure 6E. However, after adding OEO to the isolated eDNA, a significant quenching effect was seen, suggesting that the OEO intercalation had destabilized the eDNA's duplex structure.
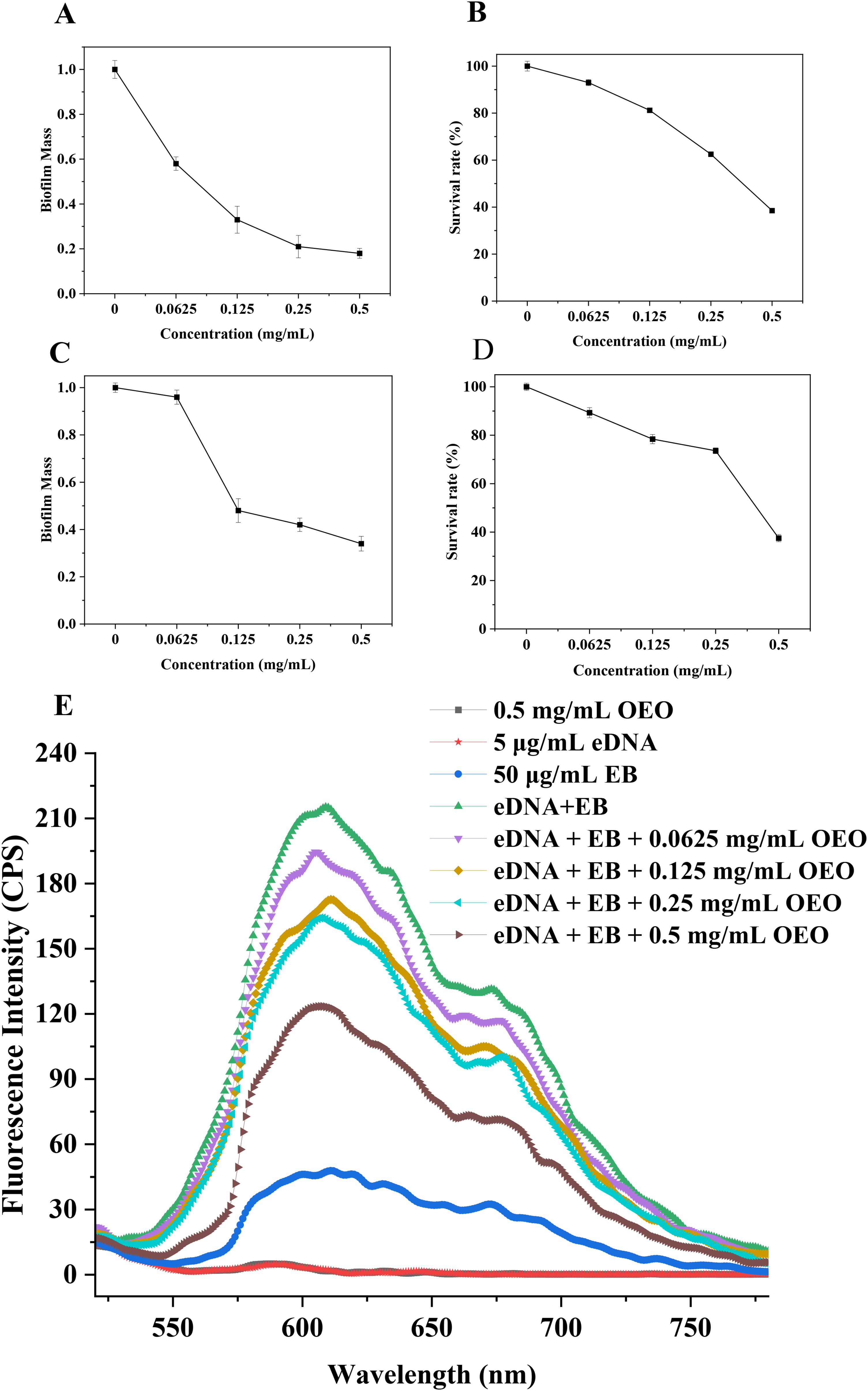
Anti-biofilms of the OEO to inhibit S mutans. (A) Crystal violet staining of S mutans biofilm mass after 24 h incubation. (B) Bacterial viability of the biofilm as measured by MTT incubation for 24 h. (C) Mature biofilm mass from various concentrations of OEO-treated S mutans. (D) Bacterial survival within the mature biofilms. (E) EB intercalated into eDNA fluorescence spectra with varying OEO concentrations present.
Chemical Components Analysis
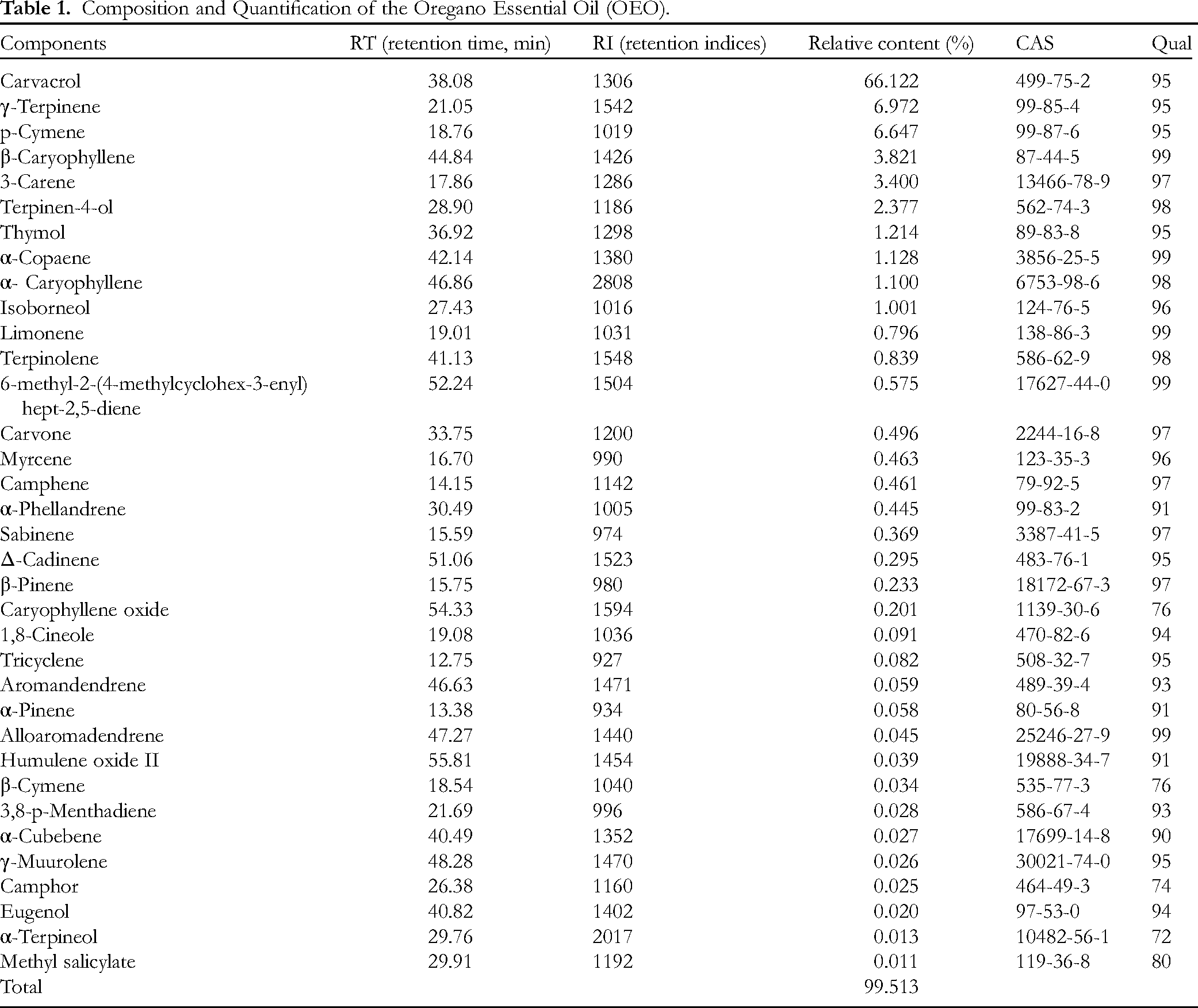
Forty volatile chemicals were found in the OEO using GC-MS technology in conjunction with the retention indices; these compounds made up 99.629% of the entire contents. Table 1 shows that the most common chemical by identification and quantification was carvacrol (66.121%), followed by γ-terpinene (6.972%), p-cymene (6.647%), β-caryophyllene (3.821%), 3-carene (3.400%), and terpinen-4-ol (2.377%). Other chemicals were detected in trace amounts, including isoborneol (1.001%), α-copaene (1.128%), thymol (1.214%), and α-caryophyllene (1.100%). In OEO, there are still 30 volatiles with extremely low contents, each with < 1% of substance.
Composition and Quantification of the Oregano Essential Oil (OEO).
Cytotoxicity
HOEC were subjected to OEO dosages that varied from 0.0625 to 1 mg/mL to investigate the cytotoxicity of OEO (Figure 7). The toxicological potential rose with increasing dosage during our incubation period, although OEO did not significantly limit HOEC growth from 0.0625 to 0.5 mg/mL. On the other hand, HOEC viability was damaged by OEO at 1 mg/mL, and this was different from the control group. However, it's worth noting that 0.5 mg/mL demonstrated strong bactericidal activity to S mutans based on the earlier antibacterial tests (Figure 1).

Cytotoxicity of the OEO to the HOEC. Cell viable rate of the HOEC under the OEO based on CCK-8. Data represent the mean ± standard deviation of 3 independent tests performed in triplicate. Significant differences (*P < .05) from the control group are indicated, while no significant differences (NS; P > .05) from the control group are indicated.
Discussion
Tests were conducted on S mutans of the OEO at doses ranging from 0.0625 to 0.5 mg/mL, with the inhibitory effects on bacteria being more pronounced at higher OEO concentrations. Notably, when comparing the OEO-treated cells to their control counterparts, the growth arrest phenomena were seen. As anticipated, the control bacteria grew regularly, followed by an increase as time passed from 0 to 24 h. Bacteria treated to the OEO at 0.0625 mg/mL exhibited a growth reduction after 12 h, followed by a rise in colonists at 24 h due to the low OEO concentration. The OEO led to a progressive decrease in the quantity of viable bacteria over the course of 24 h at concentrations ranging from 0.125 to 0.5 mg/mL. The OEO resulted in a 98% decrease in live bacteria after 24 h, particularly in 0.25 and 0.5 mg/mL concentrations. This data suggested that OEO has a bactericidal impact in addition to a bacteriostatic effect at higher doses.
The OEO often damages cellular membranes, which interferes with the bacteria's ability to function normally. This causes the bacterium to lose cytoplasm and vacuolate, which finally results in its death. The OEO can permeate the cell wall and alter the cytoplasmic membrane's structure since it is hydrophobic. 42 According to the TEM results, the OEO changed the morphology of the cell surface and obstructed the cell membrane. In accordance with earlier studies, changes to the shape of the cell membrane may have an impact on the physiological and metabolic functions of the cell, perhaps halting the growth or death of the cell. S mutans would die if treated with 0.25 mg/mL of the OEO. However, there was no discernible change in the morphological destruction of the organism between 0.25 and 0.5 mg/mL with the concentration rise. The single-cell morphology and ultrastructure alterations of S mutans can be immediately reflected in the TEM results. These results corroborate those of Min, 43 who noted that the OEO effectively inactivates clinical strains of multidrug-resistant bacteria mainly by causing cell destruction and morphological changes.
Usual bacteria had intact, uniform surfaces and a catenulate shape, while the OEO-treated cells rapidly lost their usual morphology and were lysed. However, notable surface structure alterations were seen in both OEO-treated and -untreated bacteria, with the changes being more pronounced at increasing the OEO concentrations. These surface structure changes coincided with significant variations in the ultimate biomass of the bacterial biofilm. In terms of CFU and growth curve, this result confirmed the antibacterial activity of OEO and is consistent with the previous study's findings (Figure 1). Higher OEO concentrations were associated with an increase in dead bacteria. These pictures showed that the OEO-treated group's inhibition of planktonic bacterial proliferation reduced the amounts of bacterial cells that accumulated in the biofilm. At the same time, after treatment with OEO, the biofilms seemed looser and more scattered in contrast to the control group, which showed a thick and compact structure (Figure 4A).
The water-insoluble extracellular polymer generated by S mutans promotes adherence and growth within the oral cavity. Furthermore, in the dental matrix, insoluble EPS is essential to the growth and composition of the biofilm. 44 The inhibitory impact became more apparent as the OEO concentration increased. The considerable reduction in insoluble EPS synthesis suggested that the OEO might hinder adhesion and biofilm development.
To provide more insight into how OEO affects other cariogenic traits, we also investigated the effect of OEO on acid production in S mutans. The primary metabolic route responsible for producing acids is glycolysis. 45 Lactic acid is created by the glycolysis enzyme lactate dehydrogenase (LDH), which is generated by S mutans. From this point of view, we may speculate that the pH reduction was associated with the OEO's inhibitory influence on LDH activity.
Besides regulating EPS and acid production, OEO exerted anti-biofilm effects through multiple mechanisms. The intensity of the fluorescence was greatly reduced with the introduction of OEO, and this reduction was contingent upon the concentration of OEO. According to these findings, OEO may interact with the eDNA of the biofilm by integrating into its duplex area and upsetting the structure of the double helix. The anti-biofilm results corresponded with the CLSM and SEM pictures (Figures 3 and 4). SEM and CLSM measurements of the OEO treatment showed a considerable decrease in the number of microbial cells sticking to the substratum, indicating that it had an impact on biofilm inhibition. Furthermore, a slower rate of population growth in biofilm-adhered cells may be the cause of the reduced biofilm matrix in OEO-treated circumstances. These findings showed that OEO can effectively limit biofilm formation by reducing bacterial viability. 46 OEO might prevent biofilm development by lowering EPS and acid generation during the first adhesion period. OEO might weaken the stiff structures of mature biofilms and eventually destroy them.
The antibacterial action of the OEO was related mostly to the presence of volatile chemicals, particularly carvacrol and thymol, which were shown to be responsible for the OEO's antimicrobial function as separate molecules. 47 However, many intrinsic and extrinsic factors such as agricultural factors, species, harvest time, plant part, season, climate, geography, extraction methods and storage care influence the composition of the OEO and concentration of each volatile compound, majority, or minority. Because OEO has a variety of antibacterial substances, it has a scavenging effect on mature biofilm in this paper. The anti-biofilm mechanism of OEO is probably caused by the binding of small molecular substances to the proteins related to the synthesis of biofilm, which can be investigated by computer molecular simulation software in the future. 48 In the meantime, the cytotoxicity experiment of the OEO showed that OEO was well-biocompatible when used orally in humans, suggesting that it may be turned into anti-carcinogenic therapies and eventually eliminated.
In terms of the limitation of the study, there is no in-depth target research on the interaction between OEO and virulence factor-related enzymes in the anti-biofilm of S mutans. In addition, animal dental models and clinical trials in dental patients have not been conducted in this study.
Conclusion
To summarize, the current study discovered that OEO is anti-microbial to S mutans, inhibiting the growth of planktonic bacteria. Additionally, cariogenic virulence factors such as acid generation, external polysaccharide synthesis, biofilm development, and structure were also detected. By killing the S mutans, the OEO prevented the production of biofilms on the one hand, but it also degraded the mature biofilms’ structural integrity and eventually eliminated the biofilm on the other hand. The groundwork for the OEO as a novel natural substance that prevents tooth caries by stifling the natural metabolic processes of cariogenic bacteria has been established by this study.
Footnotes
Acknowledgments
The Guangdong Provincial Key Laboratory of Plant Resources Biorefinery (2021GDKLPRB01), the Science and Technology Planning Project of Guangzhou City (202103000078, 202206010181), and the Guangdong Province Science and Technology Plan Project (22070510471437) provided funding for this work. Thank Dr Wang Pingping for her valuable contributions to this research.
Authors Contributions
Kegang Wu: conceptualization, methodology, writing – review and editing, supervision, and funding acquisition. Yuqiang Huang: writing – original draft, writing – review and editing, methodology, data curation, formal analysis, software, validation, investigation, and visualization. Tong Zhang: methodology, investigation, and supervision. Xianghua Chai: project administration, supervision, and resources. Xuejuan Duan: resources, validation, and project administration.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Guangdong Provincial Key Laboratory of Plant Resources Biorefinery, Guangdong Province Science and Technology Plan Project, Science and Technology Planning Project of Guangzhou City (grant numbers 2021GDKLPRB01, 22070510471437, 202103000078, and 202206010181).
Ethical Approval
Ethical approval is not applicable to the article.
Informed Consent
Not applicable to the article.
Statement of Human and Animal Rights
Not applicable to the article.