Recently, saponins derived from marine sources have received much attention because of their promising bioactivities, such as anticancer, anti-angiogenesis, and anti-inflammation. In particular, a triterpene saponin from the sea cucumber Cercodemas anceps Selenka, cercodemasoide A (CAN1), showed potent cytotoxicity against various cancer cell lines. Recent evidence has indicated that cancer stem cells (CSCs) could be a novel target for efficient cancer therapies. In order to improve the biopharmaceutical properties of CAN1, the compound was loaded into nanoliposomes as an ideal drug carrier. CAN1 was successfully incorporated into nanoliposomes as small unilamellar liposome vesicles with an average size of 73.39 ± 1.57 nm, zeta potential of −0.299 ± 0.046 mV, polydispersity index of 0.336 ± 0.038, and with an encapsulation efficiency of up to 62.9%. For the first time, CAN1 and its nanoliposomal forms have been shown to have a promising cytotoxic activity against NTERA-2 CSCs, with half-maximal inhibitory concentration (IC50) =1.03 ± 0.04 and 0.41 ± 0.03 µM, respectively. The CAN1 nanoliposomes also presented significantly improved activities in suppressing the growth of NTERA-2 3-dimensional tumorspheres (IC50 = 1.71 ± 0.06 µM) in comparison with the free form (P < .05). The anti-CSC effects of CAN1 nanoliposomes on NTERA-2 cells were due to their apoptotic induction through enhancing caspase-3 activity (more than 2-fold) and arresting the cell cycle at the S phase (P < .05). The obtained CAN1-encapsulated nanoliposomes suggest valuable applications in CSC-targeting treatment for more efficient clinical therapy.
Over the past several decades, cancer science has developed robustly, thereby increasing our understanding of the disease as well as making a great contribution to the development of new strategies for cancer therapy.1 Recently, a fundamental theory of cancer has been referred to cancer stem cells (CSCs). CSCs are identified as a small population of tumor cells containing self-renewing and differentiating properties.2 Thus, they have the ability to initiate and proliferate tumors as well as metastasize and recur.3 Therefore, CSCs have become an effective target for cancer treatment. The main facets of CSCs, such as ATP-binding cassette transport, signaling pathways, microenvironment, and surface markers, are all potential targets for tumor therapeutic treatments.4 Therefore, the successful use of many natural active ingredients in cancer therapy is a precondition for the application of natural compounds for targeting CSCs as well.
Marine organisms are rich sources of potential bioactive compounds for disease intervention. The classes of compounds isolated from marine organisms have unique biological properties that differ from those isolated from terrestrial organisms.5 Saponins are found in higher terrestrial plants, but also in marine invertebrates, such as sea cucumbers. Marine saponins comprise different structures and offer many health benefits, especially anticancer, anti-angiogenesis, and anti-inflammatory.6,7 Cercodemasoide A (CAN1) is a triterpene saponin, isolated from the Vietnamese sea cucumber Cercodemas anceps. In our previous study, this compound showed promising cytotoxic potency against various cancerous cell lines, such as MCF-7, HepG2, KB, LNCaP, and SK-Mel-2 cells, with half-maximal inhibitory concentration (IC50) values ranging from 0.04 to 0.24 µM.8 However, causing hemolysis is a typical feature of most saponins.9,10 In addition, because saponins are water soluble, they will decrease intestinal permeability and absorption, which can limit the bioavailability and biological activities of these compounds.11,12 Currently, the use of nanoliposomal carriers is an effective solution for improving bioavailability of natural compounds, which usually have many limitations, such as toxicity and short cardiovascular circulation. Nanoliposomes are often synthesized from cholesterol and natural phospholipids, so that the particles are relatively biocompatible with cell membranes, less frequently eliminated by the immune system, and highly biodegradable.13 Moreover, the use of nanoliposomes as drug carriers could be a new approach to treating CSCs toward influencing molecular targets, controlling drug distribution, and release.14 Therefore, in this study, we have determined the possible effects of CAN1-loaded nanoliposomes on NTERA-2 CSCs.
Material and Methods
Materials
CAN1 (a white powder, purity NLT 98%) was isolated from the Vietnamese sea cucumber C. anceps (family Cucumariidae) following the previously described procedures.8 Cholesterol was purchased from Acros Organics (Fisher Scientific, UK), and 1,2-distearoyl-sn-glycero-3-phosphoethanolamine-N-[amino(polyethylene glycol)−2000] (DSPE-PEG-2000) from Avanti polar lipids Inc. (Alabaster, AL, USA). The cell culture medium and supplements including Dulbecco’s modified Eagle medium (DMEM), fetal bovine serum (FBS), antibiotic- antimycotic (Fisher Scientific, UK), and trypsin-ethylenediamine tetraacetic acid (EDTA), fluorescein isothiocyanate (FITC) Annexin V/Dead Cell Apoptosis Kit were purchased from Invitrogen (Carlsbad, CA, USA). Caspase-3 Colorimetric Assay Kit was received from Biovision Inc. (Milpitas, CA, USA), and phosphatidylcholine (PC) and other chemical reagents from Sigma Chemical Co. (St. Louis, MO, USA).
CAN1 Nanoliposomal Fabrication and Characterization
Nanoliposomal encapsulation of CAN1 was conducted by using the Bangham hydrated-dehydrated thin film method.15 In detail, CAN1 was completely dissolved in phosphate-buffered saline (PBS; pH 7.5) at a concentration of 20 µM. The lipid mixture of PC, DSPE-PEG2000, and cholesterol (ratio of 5:2:0.05) in chloroform:methanol (2:1) was prepared in a round-bottom flask at room temperature (RT). The solvent was thoroughly removed using a rotary evaporator under an aspirate vacuum (25 mm Hg) in a water bath at 35 °C. The formed thin film on the flask walls was dispersed in 2 mL of PBS (pH 7.5) containing CAN1 (20 µM) and rotated at 10.47 rad s−1 at 25 °C. The multilamellar liposome vesicles–entrapped CAN1 produced in this process was sonicated 3 times at 20-second intervals for resizing before being filtered through a 0.22-µm membrane. The filtered CAN1 nanoliposomal mixture was washed twice with normal saline. The physicochemical features of CAN1 nanoliposomes, including polydispersity index (PDI), particle size distribution, and zeta potential were determined by using a Zetasizer Nano-Z intrument (Malvern Instruments, Malvern, UK).
Determination of CAN1 Encapsulation Efficiency
The free amount of CAN1 in the supernatant after centrifugation was determined using a liquid chromatography coupled with electrospray ionization-quadrupole-time of flight-mass spectrometry (LC-ESI-QTOF-MS) system (Agilent technology, Santa Clara, CA, USA). The sample (injection volume 2.0 µL) was automatically injected into the high-performance liquid chromatography instrument and fractionated on a Zorbax extend C18 column (2.1 × 50 mm, particle size 1.8 µm) with a gradient system of ultrapure water and acetonitrile (5%-95% acetonitrile). The mobile phase flow rate was set at 0.3 mL/min during 10 minutes of acquisition course. The QTOF instrument was set at 2 GHz extended dynamic range resolution mode, a positive ESI capillary voltage of 4000 V was used, a fragmentor voltage of 200 V, and MS acquisition rate of 2.5 spectra per second. LC-MS chromatograms were extracted with a mass range of 20 ppm around the quasi-molecular ion peak of CAN1 [M + Na]+ at m/z 1443.4544. The amount of CAN1 in the sample was calculated based on the peak area of CAN1 precursor [M + Na]+ in the extracted ion chromatogram and the standard curve, which was built from CAN1 serial diluted samples. Encapsulation efficiency of CAN1 into nanoliposomes was determined from the following formula:
where Cf is the unconjugated amount of CAN1 into liposomes measured in the supernatant, Ci is the CAN1 amount added to the lipid mixture, and EE is the encapsulation efficiency.
NTERA-2 Cell Culture
In the present study, the NTERA-2 pluripotent human embryonal carcinoma was kindly provided by Dr P. Wongtrakoongate, Mahidol University, Thailand. Cells were cultured in DMEM supplemented with 10% FBS and 1% antibiotics (anti-anti solution, Invitrogen, Carlsbad, CA, USA) in an incubator at 37 °C, 5% CO2, and 100% humidity.
Cytotoxic Assay
The cytotoxic effect of CAN1 nanoliposomes on NTERA-2 cells was identified using the Sulforhodamine B (SRB) method.16 Accordingly, cells were seeded in a 96-well plate with a concentration of 5000 cells/well and treated with CAN1 nanoliposomes at different concentrations. After 96 hours of incubation, cells were fixed with 20% trichloroacetic acid for 1 hour at 4 °C and thoroughly washed under tap water. After air-drying at RT, the cells were stained with SRB for 30 minutes. The unbound SRB was removed by washing in 1% acetic acid, 3 times. SRB dye stained with cellular proteins was dissolved in 10 mM Tris base solution. The optical density (OD) was obtained using a microplate reader at 540 nm (BioTek, ELx800).
Tumorsphere Assay
Tumorspheroids are formed when cells are cultured in a 96-well ultrathin surface plate. Five thousand single cells/well were grown in DMEM containing 20 ng/mL of epidermal growth factor, 10 ng/mL of basic fibroblast growth factor, and 6 mg/mL of bovine serum albumin. At day 3 after seeding, cells were treated with either CAN1 or its liposomes at different concentrations. New culture medium was then added to the culture wells every 48 hours to maintain cells’ growth. Cell proliferation under spherical culture conditions was evaluated after 7 days of treatment using MTT (3-(4,5-dimethylthiazol-2-yl)−2,5-diphenyltetrazolium) assay. Briefly, the medium was discarded before 50 µL of MTT solution (5 mg/mL) was added to each well. After 4 hours, all medium was removed and the formazan crystals formed were dissolved by adding 50 µL of dimethyl sulfoxide per well. The OD was measured using a microplate reader at 540 nm (BioTek, ELx800).
Detection of Apoptosis Using AnnexinV-FITC Flow Cytometry Analysis
The Annexin V-FITC assay was performed using the FITC Annexin V/Dead Cell Apoptosis Kit by following the instructions of the manufacturer (Invitrogen, Carlsbad, CA, USA). NTERA-2 cells were seeded into 6 wells plated at 1 × 106 cells/mL and incubated for 24 hours. Normal saline-incubated cells served as the negative control. Cells were then harvested by trypsinization and PBS washed to remove any remaining medium. Cell pellets were resuspended in 100 µL of binding buffer, and stained with 5 µL of AnnexinV-FITC for 15 minutes in the dark. The stained cells were washed again with binding buffer. Finally, the cells were stained with propidium iodide (PI) solution in binding buffer. The ratio of apoptotic cells was identified from an approximately counted 10 000 cells using a NovoCyte Flow cytometer system and NovoExpress software (ACEA Bioscience Inc., San Diego, CA, USA).
Cell Cycle Analysis by Flow Cytometry
NTERA-2 cells were grown in 6-well plates at a density of 1 × 106 cells/well for 24 hours at 37 °C in 5% CO2 before treatment of the samples for a further 24 hours. Cells were then harvested with trypsin-EDTA and washed 3 times with PBS (pH 7.0) before being fixed with 70% ethanol for at least 4 hours at 4 °C. The fixed cells were then washed 3 times again before being incubated with RNase A (1 mg/mL) in a 37 °C water bath for 15 minutes, followed by PI staining for 1 hour. The cell cycle arrest activity was carried out by analyzing 10 000 cells per sample using a NovoCyte flow cytometer system and NovoExpress software (ACEA Bioscience Inc.).
Caspase 3 Inducible Assay
The caspase 3 inductive activity of CAN1 nanoliposomes was determined using a Caspase-3 Colorimetric Assay Kit (Biovision Inc., Milpitas, CA, USA). Briefly, after 24 hours of either CAN1 or CAN1 nanoliposomes treatment, 1 × 106 cells were lysed in lysis buffer for 10 minutes in ice and centrifuged at 10,000 × g for 1 minute to remove the residual cellular pellet. After determination of the protein concentration of each sample, to equal protein amounts of the cell lysate from the different treatments in 50 µL assay buffer was added 50 µL DTT (10 mM) and 5 µL of DEVD-pNA (200 µM) in each well of a 96-well plate in triplicate. The plate was further incubated at 37 °C for 1 hour. The OD values were obtained using a microplate reader (BioTek, ELx800) at 405 nm wavelength.
Statistical analysis
Results were reported as mean ± standard deviation. GraphPad Prism 7 software using unpaired t-test and one-way analysis of variance was used for data analysis and determination of statistical significance with *P < .05.
Results
Physicochemical Characteristics of CAN1 Nanoliposomes
As shown in Table 1 and Figure 1, CAN1-incorporated nanoliposomes were small unilamellar liposome vesicles (SUVs) with an average size of 73.39 ± 1.57 nm, and having a zeta potential of −0.299 ± 0.046 mV. Because the PDI value of the nanoliposome particles was 0.336 ± 0.038, the CAN1-loaded nanoliposomes presented as monodispersed and monomodal particles.
Physiochemical Characteristics of CAN1-Loaded Nanoliposomes.
aBlank liposome including PC, cholesterol, and DSPE-PEG2000.
Characteristics of obtained CAN1 nanoliposomes with size and zeta voltage information measured by Nanosizer ZS90 system (Malvern, UK).
Cytotoxicity of CAN1 Nanoliposomes on NTERA-2 Cells
The toxic effects of CAN1 and CAN1 nanoliposomes on cells were determined at different concentrations. As shown in Figure 2, the cytotoxicity of CAN1 was concentration dependent. The proliferation of NTERA-2 cells was totally inhibited when treated with CAN1 at a concentration of 2 µM. This result was similar to that of CAN1 nanoliposomes at the same concentration. However, at the same lower concentration (1 µM) CAN1 liposomes still presented strong cytotoxic action to inhibit more than 90% of cells’ survival in comparison with 50.64% by unloaded CAN1. This outcome shows that CAN1 liposomes possess significantly stronger cytotoxic activity than free CAN1 (P < .05). This result is even more clearly evident from the IC50 value of CAN1 nanoliposomes (IC50 = 0.41 ± 0.03 µM), which was much lower than that of CAN1 (1.03 ± 0.04 µM) (P < .05). Blank nanoliposomal particles did not show significant effects on the cells’ growth. Therefore, this blank will not be applied in further experiments.
Effects of unloaded CAN1, CAN1-encapsulated liposomes, and blank liposomes on NTERA-2 cells after 72 hours incubation. Cultured cells (0.5 × 104 cells/well) were treated with different concentrations ranging from 0.25 to 2 µM of either liposomal or free CAN1. Normal saline served as the negative control. Each value represents the mean ± standard deviation, #P < .05 compared to CAN1 unencapsulated.
Anti-tumorspheroidal Activities of CAN1 Nanoliposomes
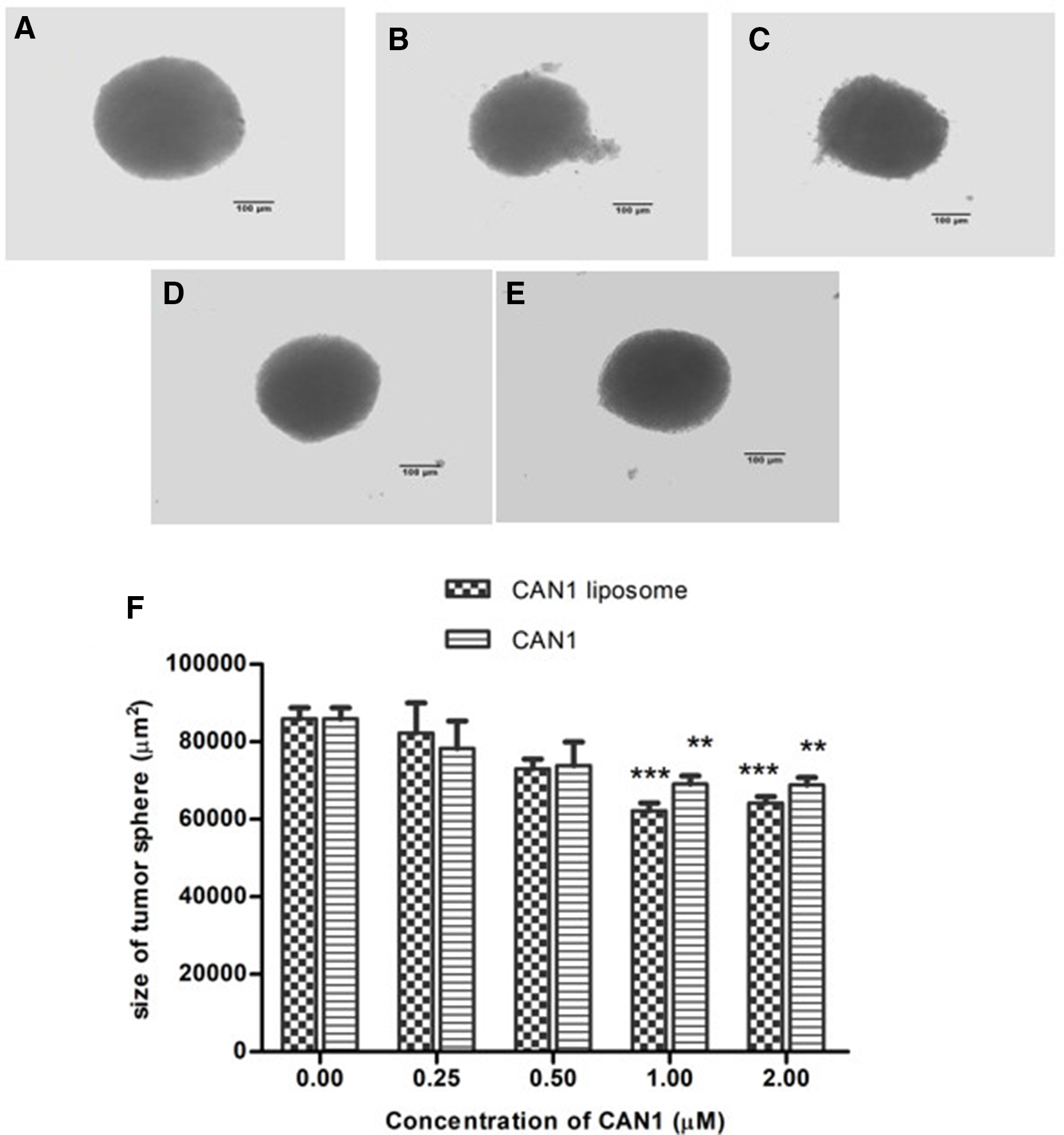
Effects of CAN1 nanoliposomes on the growth of NTERA-2 3-dimensional (3D) tumorspheres were also determined at day 7 of the experiment and compared with the free form. The results in Figure 3 show that CAN1 nanoliposomes at a 2 µM concentration significantly reduced the size and density of the NTERA-2 tumorspheroids, as well as destroying the spheroidal structure. The unloaded CAN1 at the same 2 µM concentration decreased the size, but did not demolish the structure of the spheroids. Further decreasing the concentration to 1 µM, CAN1 nanoliposomes and free CAN1 still had an effect on reducing the size of the 3D spheroids, but not their density. Through an MTT assessment, the survival rate of NTERA-2 cells in tumorspheres under sample treatment was also determined (Figure 4). The CAN1 nanoliposomes exhibited a significant cell growth-repressive activity compared with that of the unloaded form at 2 µM treatment (P < .05). The CAN1 nanoparticles presented an IC50 value of 1.74 ± 0.06 µM, whereas it was not available for CAN1 in a free form (IC50 >2 µM).
Effect of CAN1 and CAN1 nanoliposomes on NTERA-2 3-dimensional tumorsphere formation after 7 days of treatment at different concentrations; (A) negative control; (B, D) NTERA-2 tumorspheroids under treatment with CAN1 nanoliposomes at 2 and 1 µM, respectively; (C, E) NTERA-2 tumorspheres under treatment with free CAN1 at 2 and 1 µM, respectively; (F) area of tumorspheres at difference concentrations of CAN1 nanoliposomes and free CAN1; ***P < .001 and **P < .01 when compared with control.
Effects of CAN1 and CAN1 nanoliposomes at different treated concentrations ranging from 0.25 to 2 µM on the proliferation of NTERA-2 cells inside tumorspheroids. The cell viability was measured on day 7 of treatment. Normal saline served as the negative control. Each value represents the mean ± standard deviation. **P < .01 and *P < .05 compared to negative control, #P < .05 compared to CAN1 free form.
Cell Cycle Arrest Activity of CAN1 Nanoliposomes
Due to the strong inhibition of NTERA-2 proliferation in both cytotoxic and anti-tumorsphere assays of CAN1 and its nanoliposomes, we further studied their cell cycle arrest activities. Aberrant cell cycle activity is a characteristic of cancer.17 Thus, cell cycle regulators are considered attractive candidates for cancer therapy. The activities on cell cycle arrest of CAN1 and its nanoliposomes at 1 µM treatment (less toxic concentration) on NTERA-2 cells are presented in Figure 5. The majority of the NTERA-2 cells appeared in the G1 phase (51.85%), followed by the S phase (28.09%) and the G2 phase (16.83%) in the control group. Treatment with 1 µM CAN1 reduced cell number in the G1 phase (38.58%) and induced the cell number in the G2 phase (35.22%). Upon CAN1 liposome treatment, the cell number in the G1 phase also decreased significantly to 25.77%, which is a more than a 2-fold reduction compared to the control. However, cells did not transfer to the G2 phase, but were arrested in the S phase (48.14%). For the first time, the significant effects have been reported of CAN1 and CAN1 nanoliposomes on the working manner of the NTERA-2 cell cycle.
Cell cycle effects on NTERA-2 cells after 24 hours incubation with (A) negative control; (B) 1 µM of unloaded CAN1; (C) 1 µM of CAN1 in nanoliposomes using a Novocyte flow cytometry system and NovoExpress software (ACEA Bioscience Inc.).
Apoptotic Induction of CAN1 Nanoliposomes
To continue examining the potential activity against CSCs of CAN1 nanoliposomes, the NTERA-2 cell apoptotic inducibility of these samples was investigated using flow cytometry. Free CAN1 was also applied in this investigation. In this study, an apoptosis kit was used for accessing apoptosis cells under CAN1 and CAN1 liposome treatment. Cells were stained with 2 colored dyes, PI and FITC, for determination of necrotic and apoptotic cells, respectively. Herein, the apoptotic cells included a combination of cells that were FITC+/PI−/+, whereas necrotic cells were PI+ only. As shown in Figure 6, in the case of CAN1 treatment, the FITC+/PI−/+ apoptotic cell percentage efficiently increased to 44.17%, compared to 4.63% in the control group (P < .01). Moreover, a significantly higher apoptosis cell number was also recorded under CAN1 liposome treatment (59.47%) when compared to the control (P < .01). In addition, 34.87% necrosis cells were found in CAN1 liposome treatment, more than double that of free CAN1 treatment. As a result, both CAN1 and CAN1 nanoliposomes exhibited efficacious apoptosis induction on NTERA-2 CSCs.
Flow cytometry analysis of apoptotic inducibilities of NTERA-2 cells after 24 hours incubation with (A) negative control; (B) 1 µM of unloaded CAN1; (C) 1 µM of CAN1 in nanoliposomes using Novocyte flow cytometry system and NovoExpress software (ACEA Bioscience Inc.). FITC, fluorescein isothiocyanate.
The significant apoptosis activities of CAN1 and CAN1 nanoliposomes were strongly confirmed through caspase-3 activity measurement. Caspase-3 is the key protein related to programmed cell death. In our survey, caspase-3 activity was enhanced in both cases by the 0.5 and 1 µM treatments (Figure 7). However, the significance between CAN1 and CAN1 nanoliposomes’ activities was only observed at the highest treated concentration of 1 µM (P < .05).
Caspase 3 inducible effects of unencapsulated CAN1 and CAN1 nanoliposomes at different treated concentrations ranging from 0.25 to 1 µM on NTERA-2 cells after 24 hours of incubation. Normal saline served as the negative control. Each value represents the mean ± standard deviation. **P < .01 and *P < .05 compared to negative control, #P < .05 compared to unencapsulated CAN1 group.
Discussion
As reported, current cancer chemotherapies based on natural compounds have limitations, including poor specificity and poor pharmacokinetic profiles.18,19 Fortunately, the use of nanoparticles as drug carriers, especially nanoliposomes, provides new strategies for more efficient cancer therapy. Nanoliposomes have been accredited with improving circulation time, enhancing biological distribution, and even controlling drug release.20,21 Herein, nanoliposomal technology was applied for ameliorating the biomedical activities of CAN1. According to Bozzuto and Molinari, physicochemical properties of liposomes, such as surface charge, size, and composition of lipid bilayer, would affect liposomal stability and activity.20 The CAN1 liposomes were also exhibited as SUVs with an average size of 73.39 nm. Nanoliposomes smaller than 200 nm have been reported as accumulating better in tumors and circulating longer in blood vessels. They usually amplify the effects of drugs on tumors.22 Therefore, the obtained size of CAN1 nanoliposomes would help the compound improve its pharmaceutical properties, such as prolonging circulation in the vascular system and more easily accumulating in the tumors. The surface voltage of the liposome nanoparticles was also reported to have an effect on circulation time. Further, as Campos’s study shows, neutral liposomes exhibit better bioavailability in plasma, even up to 4 hours after administration.23 In the present study, CAN1 nanoparticles also expressed a zeta potential of −0.299 mV. In addition, the CAN1 nanoliposomal formulation containing PEG ensures that the nanocomplex can avoid uptake by the mononuclear phagocytic system, thus extending blood-circulation time.24
After incorporation into nanoliposomal particles, the anticancer activity of CAN1 and CAN1 liposomes was determined using the NTERA-2 cell line. Being human pluripotent embryonic carcinomas, NTERA-2 cells highly express stem signals such as Nanog homeobox, octamer-binding transcription factor 4, SRY-box 2, and teratocarcinoma-derived growth factor 1.25 Therefore, this cell line has been popularly used for accessing the anti-CSC targeted activity of potential compounds. Herein, CAN1 presented a promising cytotoxic activity in NTERA-2 cells with IC50 = 1.03 ± 0.04 µM. This parameter was higher than that on other cancerous cell lines reported by Cuong et al.8 The reason is explainable due to CSCs’ recognized drug resistance. However, CAN1 nanoliposomes showed significant cytotoxicity against NTERA-2 cells compared to the unloaded form, indicating an effective enhancement of anti-CSC activity of liposomal formulation of CAN1.
To determine further the specific cytotoxic activity of CAN1 and CAN1 liposomes on CSCs, we used an antitumor spheroidal assay. Both CAN1 and CAN1 nanoliposomes expressed the potential inhibition of the CSCs’ renewable property. Likewise, CAN1 and CAN1 nanoliposomes also effectively suppressed the proliferation of NTERA-2 cells in the spheroids. According to Bahmad, CSCs have the ability to form multicellular 3D tumorspheres when they grow in nonadherent and serum-free conditions. Thus, inhibition of tumorspheroidal formation is a useful assay to determine the effects of a compound on CSCs’ renewable potency, as well as on a cellular population that is located inside the tumorspheres.26 In this study, CAN1 has been recognized for its effects on tumorspheroidal size and internal cellular proliferation, whereas the activities of CAN1 nanolipsomes were significantly enhanced in comparison with the unloaded form.
In cancer, cells that do not undergo apoptosis, a destruction program that controls cell death, become immortal.27,28 Apoptosis is a potential target for cancer therapy. Flow cytometry analysis, which is based on annexinV-FITC intensity, helps to determine the percentage of apoptosis-induced cells. The principle of this assay is based on the inner or outer location of phosphatidylserine (PS) on the cell membrane. PS only exposes on the outer leaflet of the plasma membrane during apoptosis. This lipid binds with annexin V, which is labeled fluorescent so that it can be used to detect apoptotic cells.29 Using this method, we found that CAN1 at 1 µM strongly induced apoptosis. CAN1 liposomes also enhanced the percentage of apoptotic cells more effectively than free CAN1. However, both CAN1 and CAN1 nanoliposomes also induced cell death by necrosis. In apoptotic progression, the proteases called caspases cause the morphological and metabolic changes in apoptotic cells. There are 2 apoptosis pathways, extrinsic and intrinsic, but both are activated through the effector protease, caspase-3.30 Therefore, caspase-3 is considered the key enzyme for apoptosis-inducing pathways. In our experiments, CAN1 and CAN1 liposomes effectively and significantly induced caspase-3 activity compared to the untreated control. This result again provides strong evidence for the apoptosis inducibility of CAN1 and CAN1-loaded nanoliposomes.
Conclusions
CAN1 was for the first time loaded into nanoliposomal particles with EE = 62.91%. The obtained CAN1 nanoliposomes were SUVs with an average size of 73.39 ± 1.57 nm, zeta potential of −0.299 ± 0.046 mV, and PDI of 0.336 ± 0.038. The compound and its nanoliposomes were assessed for anti-CSC activities on NTERA-2 cells. The acquired results showed potential cytotoxic as well as 3D anti-tumorsphere activities of both CAN1 and CAN1 nanoliposomes. The efficient anti-CSC abilities of CAN1 and its encapsulated liposomes might be related to their strong apoptotic induction and cell cycle arrest at a very low concentration of 1 µM. Significant anti-CSC enhanced potency was also observed in the CAN1-loaded nanoliposomes compared to the free form. These results suggest improved bioavailability and applicable capacities of CAN1 nanoliposomes for CSC-targeting treatment.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Vietnam Academy of Science and Technology (VAST) (VAST04.10/18-19).
ORCID iD
Phan Van Kiem
References
1.
ArteagaCL.AdamsonPC.EngelmanJAet al. Aacr cancer progress report 2014. Clin Cancer Res. 2014;20(19 Suppl):S1-S112.doi:10.1158/1078-0432.CCR-14-2123http://www.ncbi.nlm.nih.gov/pubmed/25228531
2.
YuZ.PestellTG.LisantiMP.PestellRG. Cancer stem cells. Int J Biochem Cell Biol. 2012;44(12):2144-2151.doi:10.1016/j.biocel.2012.08.022http://www.ncbi.nlm.nih.gov/pubmed/22981632
3.
ChenK.HuangY-hui.ChenJ-long.ChenJ. Understanding and targeting cancer stem cells: therapeutic implications and challenges. Acta Pharmacol Sin. 2013;34(6):732-740.doi:10.1038/aps.2013.27http://www.ncbi.nlm.nih.gov/pubmed/23685952
4.
HuY.FuL. Targeting cancer stem cells: a new therapy to cure cancer patients. Am J Cancer Res. 2012;2(3):340-356.http://www.ncbi.nlm.nih.gov/pubmed/22679565
5.
LindequistU. Marine-derived pharmaceuticals-challenges and opportunities. Biomol Ther. 2016;24(6):561-571.doi:10.4062/biomolther.2016.181http://www.ncbi.nlm.nih.gov/pubmed/27795450
6.
BahramiY.ZhangW.ChatawayT.FrancoC. Structure elucidation of five novel isomeric saponins from the viscera of the sea cucumber Holothuria lessoni. Mar Drugs. 2014;12(8):4439-4473.doi:10.3390/md12084439http://www.ncbi.nlm.nih.gov/pubmed/25110919
7.
HeY.HuZ.LiAet al. Recent advances in biotransformation of saponins. Molecules. 2019;24(13):2365.doi:10.3390/molecules24132365
8.
CuongNX.VienLT.HanhTTHet al. Cytotoxic triterpene saponins from Cercodemas anceps. Bioorg Med Chem Lett. 2015;25(16):3151-3156.doi:10.1016/j.bmcl.2015.06.005http://www.ncbi.nlm.nih.gov/pubmed/26099533
9.
PodolakI.GalantyA.SobolewskaD. Saponins as cytotoxic agents: a review. Phytochem Rev. 2010;9(3):425-474.doi:10.1007/s11101-010-9183-zhttp://www.ncbi.nlm.nih.gov/pubmed/20835386
10.
BissingerR.ModicanoP.AlzoubiKet al. Effect of saponin on erythrocytes. Int J Hematol. 2014;100(1):51-59.doi:10.1007/s12185-014-1605-zhttp://www.ncbi.nlm.nih.gov/pubmed/24924952
11.
GaoS.BasuS.YangZ.DebA.HuM. Bioavailability challenges associated with development of saponins as therapeutic and chemopreventive agents. Curr Drug Targets. 2012;13(14):1885-1899.doi:10.2174/138945012804545498http://www.ncbi.nlm.nih.gov/pubmed/23140297
12.
ChenC.HanX.DongPet al. Sea cucumber saponin liposomes ameliorate obesity-induced inflammation and insulin resistance in high-fat-diet-fed mice. Food Funct. 2018;9(2):861-870.doi:10.1039/C7FO01599Bhttp://www.ncbi.nlm.nih.gov/pubmed/29292463
13.
AkbarzadehA.Rezaei-SadabadyR.DavaranSet al. Liposome: classification, preparation, and applications. Nanoscale Res Lett. 2013;8(1):102.doi:10.1186/1556-276X-8-10223432972
14.
LuB.HuangX.MoJ.ZhaoW. Drug delivery using nanoparticles for cancer stem-like cell targeting. Front Pharmacol. 2016;7:84.doi:10.3389/fphar.2016.0008427148051
15.
BanghamAD.StandishMM.WatkinsJC. Diffusion of univalent ions across the lamellae of swollen phospholipids. J Mol Biol. 1965;13(1):238-IN27.doi:10.1016/S0022-2836(65)80093-6http://www.ncbi.nlm.nih.gov/pubmed/5859039
16.
SkehanP.StorengR.ScudieroDet al. New colorimetric cytotoxicity assay for anticancer-drug screening. J Natl Cancer Inst. 1990;82(13):1107-1112.doi:10.1093/jnci/82.13.1107http://www.ncbi.nlm.nih.gov/pubmed/2359136
17.
OttoT.SicinskiP. Cell cycle proteins as promising targets in cancer therapy. Nat Rev Cancer. 2017;17(2):93-115.doi:10.1038/nrc.2016.138http://www.ncbi.nlm.nih.gov/pubmed/28127048
18.
PandeyHRaniRAgarwalVMotilal Nehru National Institute of Technology, India. Liposome and their applications in cancer therapy. Braz Arch Biol Technol. 2016;59:e16150477.doi:10.1590/1678-4324-2016150477
19.
DadwalA.BaldiA.Kumar NarangR. Nanoparticles as carriers for drug delivery in cancer. Artif Cells Nanomed Biotechnol. 2018;46(sup2):295-305.doi:10.1080/21691401.2018.1457039http://www.ncbi.nlm.nih.gov/pubmed/30043651
20.
BozzutoG.MolinariA. Liposomes as nanomedical devices. Int J Nanomedicine. 2015;10:975-999.doi:10.2147/IJN.S68861http://www.ncbi.nlm.nih.gov/pubmed/25678787
21.
SercombeL.VeeratiT.MoheimaniF.WuSY.SoodAK.HuaS. Advances and challenges of liposome assisted drug delivery. Front Pharmacol. 2015;6:286.doi:10.3389/fphar.2015.00286http://www.ncbi.nlm.nih.gov/pubmed/26648870
22.
NagayasuA.UchiyamaK.KiwadaH. The size of liposomes: a factor which affects their targeting efficiency to tumors and therapeutic activity of liposomal antitumor drugs. Adv Drug Deliv Rev. 1999;40(1-2):75-87.doi:10.1016/S0169-409X(99)00041-1http://www.ncbi.nlm.nih.gov/pubmed/10837781
23.
Campos-MartorellM.Cano-SarabiaM.SimatsAet al. Charge effect of a liposomal delivery system encapsulating simvastatin to treat experimental ischemic stroke in rats. Int J Nanomedicine. 2016;11:3035-3048.doi:10.2147/IJN.S107292http://www.ncbi.nlm.nih.gov/pubmed/27418824
24.
ImmordinoML.DosioF.CattelL. Stealth liposomes: review of the basic science, rationale, and clinical applications, existing and potential. Int J Nanomedicine. 2006;1(3):297-315.http://www.ncbi.nlm.nih.gov/pubmed/17717971
25.
SchwartzCM.SpivakCE.BakerSCet al. NTera2: a model system to study dopaminergic differentiation of human embryonic stem cells. Stem Cells Dev. 2005;14(5):517-534.doi:10.1089/scd.2005.14.517http://www.ncbi.nlm.nih.gov/pubmed/16305337
26.
BahmadHF.CheaitoK.ChalhoubRMet al. Sphere-formation assay: three-dimensional in vitro culturing of prostate cancer stem/progenitor sphere-forming cells. Front Oncol. 2018;8:347.doi:10.3389/fonc.2018.0034730211124
27.
WangY-H.ScaddenDT. Harnessing the apoptotic programs in cancer stem-like cells. EMBO Rep. 2015;16(9):1084-1098.doi:10.15252/embr.201439675http://www.ncbi.nlm.nih.gov/pubmed/26253117
28.
AungTN.QuZ.KortschakRD.AdelsonDL. Understanding the effectiveness of natural compound mixtures in cancer through their molecular mode of action. Int J Mol Sci. 2017;18(3):656.doi:10.3390/ijms1803065628304343
29.
CrowleyLC.MarfellBJ.ScottAP.WaterhouseNJ. Quantitation of apoptosis and necrosis by annexin V binding, propidium iodide uptake, and flow cytometry. Cold Spring Harb Protoc. 2016;2016(11):pdb.prot087288-1072.doi:10.1101/pdb.prot087288http://www.ncbi.nlm.nih.gov/pubmed/27803250
30.
ReedJC. Mechanisms of apoptosis. Am J Pathol. 2000;157(5):1415-1430.doi:10.1016/S0002-9440(10)64779-7http://www.ncbi.nlm.nih.gov/pubmed/11073801