
Abstract
The effect of emodin on complete Freund’s adjuvant (CFA)-induced inflammatory pain in rats and its potential molecular mechanism was investigated. For this, a rat model of inflammatory pain induced by CFA was established and rats were treated with emodin by intraperitoneal injection. The pain threshold was evaluated by the von Frey, thermo hyperalgesia, and cold plate tests. The mRNA expression of transient receptor potential channel ankyrin type-1 (Trpa1) and transient receptor potential vanilloid 1 (Trpv1) was detected by quantitative reverse transcription polymerase chain reaction, and the level of inflammatory cytokines was determined by enzyme-linked immunosorbent assay. The mechanical and thermal pain thresholds of CFA-treated rats were significantly lower than those of the control rats, while the paw withdrawal responses in response to cold stimulation were higher than that of the control group. Emodin treatment significantly improved CFA-induced hyperalgesia. Further results showed that emodin inhibits the upregulation of Trpa1 and Trpv1 mRNA expression in the dorsal root ganglion (DRG) of rats with inflammatory pain compared with the control group. Emodin also significantly reduced the levels of tumor necrosis factor alpha (TNF-α), interleukin 1 beta (IL-1β), and interleukin 6 (IL-6) in the serum of rats with inflammatory pain. Thus, emodin may inhibit hyperalgesia induced by inflammatory stimulation by downregulating the mRNA expression of Trpa1 and Trpv1 in DRG neurons and reducing the levels of TNF-α, IL-1β, and IL-6.
Pain is one of the most common problems of daily life in the clinic, and inflammatory pain is the most common type. 1,2 However, chronic inflammation can cause persistent tissue damage and unwarranted pain due to the release of inflammatory mediators, including bradykinin, prostaglandins, and various cytokines, released locally during tissue damage or by inflammatory cells. 3 Inflammatory pain is characterized by increased activity of primary afferent nerves (nociceptors), resulting in spontaneous pain, hyperalgesia, and allodynia. 4 As we all know, inflammatory pain will seriously affect the quality of life and labor ability of patients, which undoubtedly brings great suffering to patients’ families. It has been reported that physical factors such as mechanical injury and high temperature, as well as inflammatory mediators (such as tumor necrosis factor alpha [TNF-α], interleukin 1 beta [IL-1β], and interleukin 6 [IL-6]) in the nervous system can cause inflammatory pain, 5,6 but the detailed mechanism of pain transduction is still unclear. Meanwhile, limited to the current research foundation and treatment level, the clinical treatment of inflammatory pain is not satisfactory, and is still a huge problem. 6,7
The ion channels and receptors in the dorsal root ganglions (DRGs) are involved in the transmission of electrical signals in the nervous system and the detection of harmful external stimuli. 8 The occurrence and development of pain are usually closely related to changes in the plasticity of DRGs. 9 Extensive research has shown that pain is often accompanied by changes in the expression of transient receptor potential (TRP) channels in DRGs. 10 More importantly, transient receptor potential channel ankyrin type-1 (TRPA1) and transient receptor potential vanilloid 1 (TRPV1) are widely regarded as important molecular sensors for pain signal generation in DRG neurons in various TRP channels. 11,12
Emodin (1,3,8-trihydroxy-6-methylanthraquinone), a natural anthraquinone derivative, is the main component of rhubarb and other medicinal materials. 13 A large number of studies showed that emodin has a variety of biological characteristics. 14 Rhubarb is an important medicinal material for the treatment of many diseases, and its active ingredient, emodin, has become a hot spot of research. This compound has a variety of pharmacological effects, such as antiviral, antibacterial, antiallergic, antidiabetic, immunosuppressive, and hepatoprotective activities. 15 More importantly, in a large number of clinical practices of Chinese medicine, emodin is often used for the treatment of constipation and pain-related diseases. 14
In this study, we sought to elucidate the influence of the effect of emodin on complete Freund’s adjuvant (CFA)-induced inflammatory pain in rats, and further clarify its potential molecular mechanism of analgesia.
Materials and Methods
Animals and Treatments
Animal experiments conformed to the Animal Care Committee of Yangtze University and the guidelines for pain research with laboratory animals. Male Sprague-Dawley rats (200-250 g) were selected for behavioral experiments and the induced inflammatory pain model. Rats were housed in individual cages in temperature-controlled micro-isolator filter cages (25 ± 1 °C) under a 12-hour light–dark cycle, with free access to food and water.
Before model induction, baseline nociceptive thresholds were measured for 3 days. Inflammatory pain in rats was induced by subcutaneously injecting 50 µL of CFA (Sigma, St Louis, MO, USA) into the dorsal surface of the rat’s left hind paw, as described previously. 16 Finally, rats were assessed for behavioral pain every other day for 7 days after CFA injection.
Emodin (25, 50, or 100 mg/kg) was introduced into rats in the CFA group (day 3 after CFA injection) and sham group through intraperitoneal injections (0.5 mL, n = 6 rats/group). After repeated treatment with emodin for 5 days, its effect (day 5 after emodin treatment) on CFA-induced inflammatory pain was evaluated by behavioral testing 1 hour after treatment on day 5. The preventive effect of emodin on CFA-induced inflammatory pain was investigated by treatment with 0.5 mL emodin (50 mg/kg) at the same time as either CFA or normal saline injection (n = 6). Emodin was administered repeatedly for 7 days, and its preventive effect on CFA-induced inflammatory pain on days 0, 1, 3, 5, and 7 of emodin was evaluated by behavioral evaluation 1 hour after administration.
Behavioral Tests
Mechano-allodynia (von Frey) test
Mechanical sensitivity was measured using the von Frey test to assess the paw withdrawal response to mechanical stimulation to serially increasing filament stiffness. Rats were placed in plastic cages with wire mesh at the bottom, and each rat was allowed to adapt before the test. von Frey filaments were used on the middle surface of the sole of the left hind paw of each rat, starting from 1 g and ranging from 0 to 50 g. The filament touches the hind paw until it bends, while the filament should remain perpendicular to the side of the plantar of the paw for about 6-8 seconds. 17 The withdrawal threshold data were determined in grams as paw withdrawal thresholds (PWTs).
Thermal pain sensitivity test
For the thermal pain sensitivity test, a hot plate was used to test the paw retraction latency (PWL) of all rats to evaluate the thermal pain threshold. The hot plate consists of a 25 cm × 25 cm metal plate and a cage with glass. Before the test, each rat was placed in a hot plate device for 10 minutes to allow it to adapt without heating. It was ensured that the 2 hind claws of the rat gently touched the surface of the plate without any force applied, and the hot plate was set at 54 °C with a cutoff time of 30 seconds. The pain threshold was measured by the response time of shaking, retracting, or licking the paw, and the threshold data were determined from the average interval of 15 minutes in 3 experiments. 18
Cold plate test
As previously described 19 behavioral tests of cold abnormal pain were performed in rats. The rats in each experimental group were placed on a glass plate kept at a low temperature (4 ± 1 °C). The stimulation time of each mouse’s alternately stimulated paws was 5 minutes, and each paw was tested 4 times in total. The number of withdrawal/licking responses was recorded over 20 seconds.
Reverse Transcription-Quantitative Polymerase Chain Reaction
According to the manufacturer’s instruction manual, the DRGs of the saline group (n = 6) and the CFA group (n = 6) were collected after 1 hour of daily treatment with emodin. Total RNA of each sample was extracted using TRIzol reagent (Invitrogen, Carlsbad, CA, USA), and RNA concentration of each sample was quantified by Nanodrop 2000 (Thermo Fisher Scientific, USA) spectrophotometry. Next, 1 µg of each sample was reverse transcribed into cDNA using the PrimerScript RT Master Mix kit (Takara Bio, Japan), followed by quantitative polymerase chain reaction (qPCR) experiments. The qPCR reaction system was prepared according to the SYBR Premix Ex Taq (Takara Bio, Japan) kit instructions. The qPCR reaction system was incubated at 95 °C for 3 minutes, and then the target gene fragment was amplified by 40 cycles at 95 °C for 10 seconds and 58 °C for 10 seconds. The primer sequences used in qPCR experiments were as follows: Trpa1 forward primer 5’-TGG ATG AGG ACA ATG ATG GA-3’, reverse primer 5’-TTT TGG AAT GAA CGG ACA CA-3’. Trpv1 forward primer 5’-GAC ATG CCA CCC AGC AGG-3’, reverse primer 5’-TCA ATT CCC ACA CAC CTC CC-3’. Gapdh forward primer 5’-TGC TGA GTA TGT CGT GGA G-3’, reverse primer 5’-GTC TTC TGA GTG GCA GTG AT-3’. The expression level of the gene was calculated by the quantitative cycle (Cq) value according to the formula 2-ΔΔCq, and the expression level of the GAPDH gene was used as an internal control.
Enzyme-Linked Immunosorbent Assay
The levels of serum TNF-α, IL-1β, and IL-18 were measured by a Parameter TNF-α, IL-1β, and IL-6 enzyme-linked immunosorbent assay (ELISA) kit (R&D Systems, USA) according to the manufacturer’s protocol (n = 6 in each group). Each experiment was repeated at least 3 times.
Data and Statistical Analysis
Experimental data are expressed as the mean ± standard error of the mean. Groups were compared using either 1- or 2-way analysis of variance or Student’s t-test where appropriate, except for time course experiments that used a repeated-measures 2-way analysis of variance. P < .05 was taken as a significant difference. All data were processed in GraphPad Prism 5 software (La Jolla, CA, USA).
Results
Establishment of CFA-Induced Inflammatory Pain Model in Rats
To establish the inflammatory pain model for rats, a certain dose of CFA was injected into the plantar part of the hind paw. Several hours after the injection, obvious signs of local inflammation (such as swelling) were observed at the injection site. As previously reported, these results have demonstrated the ability of CFA to induce inflammatory pain. Compared with the baseline measurement (sham control), the pain threshold (PWT) of the CFA group in the mechanical sensitivity and thermal sensitivity tests was decreased significantly on the first day after injection and lasted until the seventh day (Figure 1(A) and (B)). At the same time, the cold plate test showed that paw withdrawal responses to cold stimulation in the CFA group on the seventh day was significantly higher than that of the sham control group (Figure 1(C)). Our results show that the CFA-induced inflammatory pain model in rats had been successfully established.
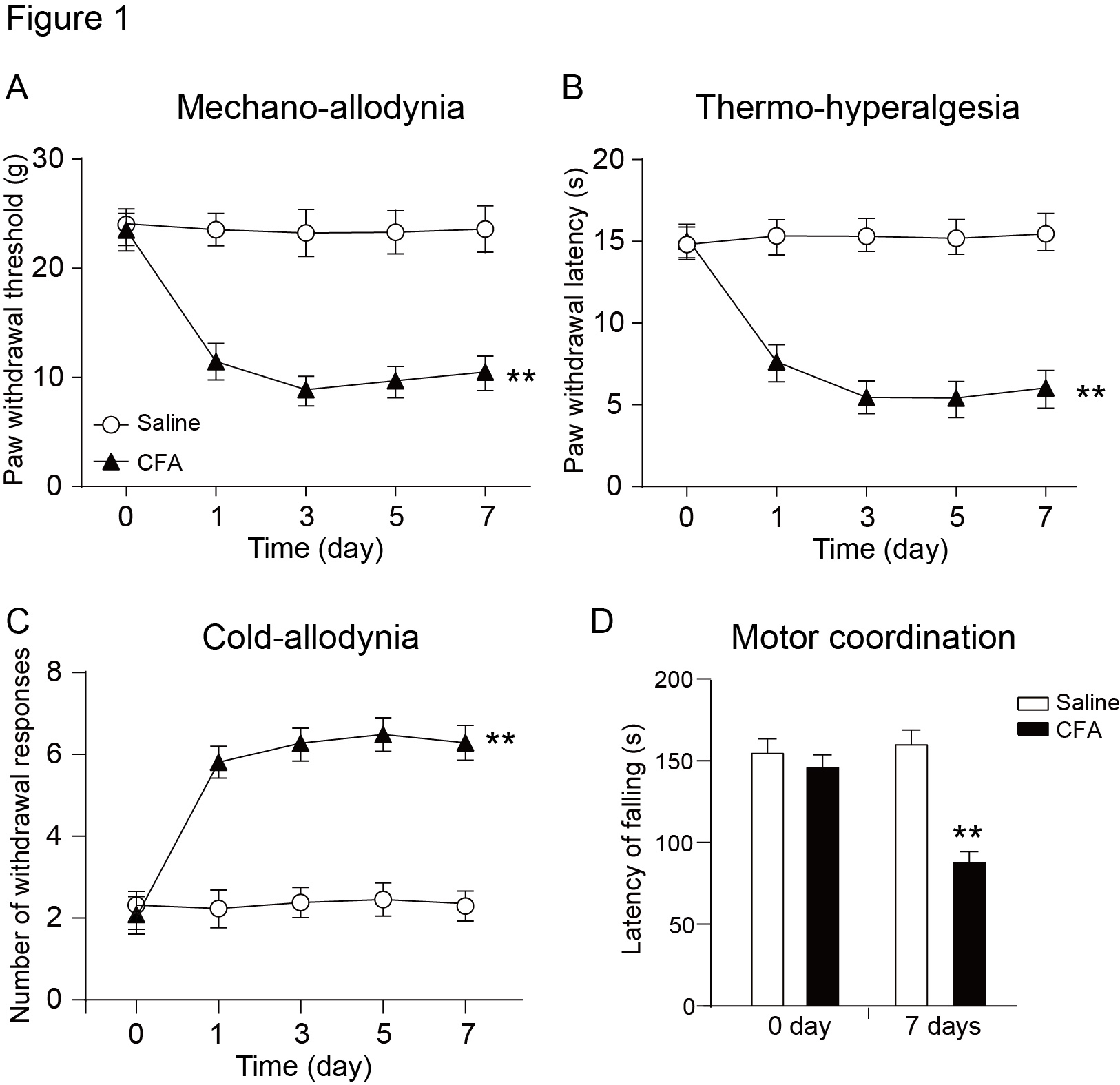
Behavioral assessments of the CFA-induced inflammatory pain model rats. Rats received either an intra-articular injection of CFA ( Sigma-Aldrich, USA) or saline, followed by sequential pain behavioral tests (n = 6). (
Emodin Reverses CFA-Induced Hyperalgesia and Abnormal Pain
To evaluate the effect of emodin on inflammatory pain, different doses of emodin (0, 25, 50, and 100 mg/kg) were given to the rats with CFA-induced inflammatory pain. The mechanical sensitivity test results showed that compared with the CFA group without emodin (0 mg/kg) injection, intraperitoneal injection of emodin could significantly alleviate CFA-induced hyperalgesia in a dose-dependent manner, and PWT increased by about 50% when emodin was injected at 50 mg/kg (Figure 2(A), P < .01). Similarly, in the hot plate test, compared with the CFA group without emodin treatment, 50 mg/kg emodin injection significantly increased the paw withdrawal latency by about 55% (Figure 2(B), P < .01). The results of the cold plate test showed that with the increase in emodin concentration, the cold pain sensitivity threshold of CFA rats decreased significantly (Figure 2(C), P < .01). The above results showed that emodin could significantly alleviate the hyperalgesia and abnormal pain induced by CFA.

Emodin relieves CFA-induced inflammatory pain in a dose-dependent manner. Emodin (0, 25, 50, and 100 mg/kg) or saline was injected intraperitoneally into CFA-induced inflammatory pain rats, and (
Emodin Can Effectively Prevent CFA-Induced Hyperalgesia and Abnormal Pain
To evaluate further the role of emodin in the occurrence and development of inflammatory pain, we injected CFA into the plantar of rats and 50 mg/kg emodin solution into the abdominal cavity and evaluated its effect on the occurrence and development of inflammatory pain through a series of pain behavior tests. As shown in Figure 3, compared with the saline and “saline + emodin” groups, the pain threshold of the CFA group in the mechanical sensitivity and thermal sensitivity tests decreased significantly after 1 day of treatment; this was maintained to the seventh day. Interestingly, intraperitoneal injection of emodin solution with CFA at the same time increased the CFA-induced pain threshold significantly on the seventh day of treatment (Figure 3(A) and (B), P < .05). Moreover, we also found that emodin treatment in the process of CFA-induced inflammatory pain can significantly reduce the CFA-induced cold pain threshold (Figure 3(C), P < .05).

Emodin prevents the progress of inflammatory pain caused by CFA. (
Emodin Inhibits the Upregulation of TRPA1 and TRPV1 Expression in Inflammatory Pain
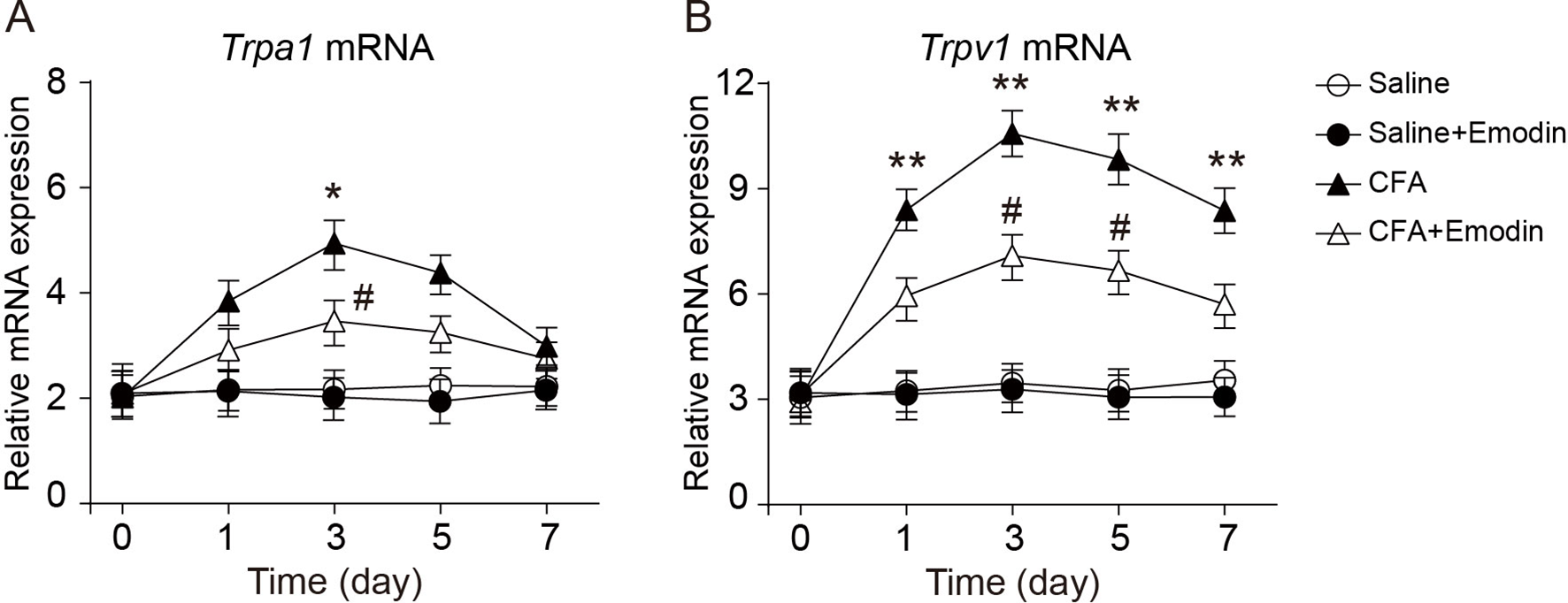
Trpv1 and Trpa1 are pain-related ion channels, which can be activated by physiological processes such as inflammation. 20,21 To determine whether alleviation and prevention of CFA-induced inflammatory pain by emodin is related to the regulation of Trpa1 and Trpv1 expression, we measured the expression of Trpa1 and Trpv1 mRNA in DRG of CFA rats after emodin injection. Compared with the saline and emodin groups, the expression level of Trpa1 and Trpv1 mRNA in DRG of rats was significantly upregulated 1 day after injection of CFA and reached a peak on the third day after injection. Interestingly, intraperitoneal injection of emodin with CFA at the same time significantly reduced the expression of Trpa1 and Trpv1 mRNA in CFA-induced DRG in rats.
Emodin Downregulates the Serum Levels of TNF-α, IL-1β, and IL-6 in CFA Rats
To clarify further the mechanism of emodin in relieving and preventing CFA-induced hyperalgesia and abnormal pain, we measured the levels of TNF-α, IL-1β, and IL-6 in the serum of each group by ELISA. As shown in Figure 4, the levels of TNF-α, IL-1β, and IL-6 in the CFA group were significantly higher than those in the control group. Injection of emodin into the CFA-induced inflammatory pain rats significantly reduced the levels of TNF-α, IL-1β, and IL-6 (P < .05).
Emodin inhibits the expression of TNF-α, IL-1β, and IL-6 in inflammatory pain rats. (
Discussion
In this study, a model of peripheral inflammatory pain was induced in rats by subcutaneous injection of CFA, and emodin was injected intraperitoneally to explore its effect on the induced inflammatory pain and its possible molecular mechanism. We observed that compared with the control group, CFA can cause peripheral inflammatory pain in rats, PWT and PWL values are significantly reduced, and the cold pain sensitivity is significantly increased, indicating that the rats are more sensitive to external stimuli (Figure 1). When we either pretreated or treated CFA-induced peripheral inflammatory pain rats with emodin, it could significantly alleviate the hyperalgesia and restore the significantly downregulated PWT and PWL values to near-normal levels (Figures 2 and 3).
TRPA1 and TRPV1 channels have been widely demonstrated to be expressed in peripheral sensory neurons and are closely related to the development of hyperalgesia. 11 Studies have shown that Trpv1 and Trpa1 can act as sensory sensors for nociceptive mechanical stimulation, or play an important role in mechanical hyperalgesia. 22 Similarly, studies have shown that injection of CFA can stimulate the expression of Trpa1 and Trpv1 in hyperalgesia-related ion channels. 23 Although our results show that Trpa1 and Trpv1 at the DRG level contribute to the maintenance of chronic inflammatory pain, they are not the only mechanism for the development of inflammatory pain. It has been shown that in young TRPA1-knockout mice with CFA-induced arthritis, peak development of mechanical hypersensitivity is delayed until 4 weeks. 4 Trpa1 and Trpv1 endogenous activators of DRG levels in inflammatory pain remain uncertain. Our results in this study showed that Trpa1 and Trpv1 expression levels were significantly upregulated in the spinal dorsal ganglia of rats after CFA injection (Figure 5). Interestingly, when rats were treated with emodin, we found that it could significantly downregulate the increase in Trpa1 and Trpv1 expression caused by CFA (Figure 5). We speculate that emodin relieves CFA-induced inflammatory pain by downregulating TRPA1 and TRPV1 expression in DRGs.

Emodin prevents the upregulation of Trpa1 and Trpv1 mRNA expression in dorsal root ganglions of inflammatory pain rats. (
The inflammatory process is a common pathophysiological process, which is the body’s stress response to external physical or chemical factors, and usually involves the accumulation of proinflammatory cytokines such as TNF-α, IL-1β, and IL-6. 24 -26 Numerous studies have shown that pain is often produced in the spinal cord, anterior cingulate cortex, and nervous tissues with elevated levels of TNF-α, IL-1β, and IL-6 mRNA and protein. 27,28 Inflammatory molecules such as TNF-α, IL-1β, and IL-6 are considered as important biomolecules involved in the development of pain. 29 At the same time, previous studies have shown that emodin has a bacteriostatic and anti-inflammatory effect. 14 Therefore, in this study we collected the serum of rats in each experimental group for the detection of TNF-α, IL-1β, and IL-6 expression levels. Consistent with previous reports, CFA significantly increased protein expression levels of TNF-α, IL-1β, and IL-6 compared with the control group and significantly reduced the upregulation of these factors when treated with emodin (Figure 4).
In summary, our results suggest that emodin may alleviate inflammatory pain by downregulating the expression levels of Trpa1 and Trpv1 in DRG neurons, and reducing the inflammatory factors TNF-α, IL-1β, and IL-6 expression in serum.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants from the Natural Science Foundation of Hubei Province (No. 2018CKB920).