
Abstract
Peptides have been reported to possess interesting biological properties. The present study was designed to evaluate neuroprotective and memory-enhancing effects of antioxidant peptide from walnut (Juglans regia L.) protein hydrolysates. The neuroprotective effect of walnut peptide (WP) against oxidative stress on PC12 cells was evaluated. And zebrafish was used as the model to corroborate the effect. Its effect on learning and memory of mice using the Morris water maze and the step-down passive avoidance tests were performed. Moreover, the acute toxicity of WP was carried out to assess its safety profile. It was found that WP was able to suppress H2O2-induced cell death in PC12 cells. In the zebrafish model, WP had an obvious neuroprotective effect, and the ratio reached 42% at 222 µg/mL. The mechanism study revealed that WP could inhibit the activity of caspases 3/7 and 8, reduce the mRNA expression levels of Bax and glial cell line-derived neurotrophic factor, and improve the mRNA expression level of brain-derived neurotrophic factor significantly. Besides, the treatment of mice with WP shortened the escape latency and exhibited much longer target time and more crossing times significantly, compared with untreated control groups in the Morris water maze test. Similarly, the step-down passive avoidance test showed that WP could ameliorate memory impairments. The administrated dose (20.1 g/kg body weight [BW]) did not produce mortality or treatment-related adverse effects with regard to BW, general behavior, or relative organ weights of the tested male and female mice. The current results indicated that WP could exert neuroprotective effect, and attenuated learning and memory impairments. These ameliorating effects of WP may be useful for treatment of memory impairment in Alzheimer’s and its related diseases.
Peptides are natural biological or artificially manufactured short chains of amino acid monomers linked by peptide (amide) bonds. 1 -3 Bioactive peptides always contain 2 to 20 amino acids with possible bioactivities, 4,5 including antihypertensive, 6,7 antioxidant, 8 -10 antimicrobial, 11 anticancer, 12 -14 and opioid activity. 15
Walnut (Juglans regia L.) belonging to the genus Juglans is the most widespread tree nut in the world, and it is one of the oldest tree nuts, which is native to Southeastern Europe, Eastern Asia, and Northern America. 16 -20 It has been cultivated in China for more than 2000 years for harvesting walnut fruit. 21 Walnut is a highly popular nut, because it possesses many important physical and functional activities such as antioxidant activity, 22 anti-inflammatory activity, 23 prevention of cancer, 24,25 lowering of cholesterol, 26 reduction of cardiovascular risk, 27 and promotion of cognition. 28,29 Walnut is rich in unsaturated fatty acids, 30 proteins, 31 polyphenols, 32 and minerals, 33 so it is perceived as a nutrient-dense “brain food.” The unsaturated fatty acids have nutritional advantages in lowering blood cholesterol and antioxidizing. 34,35 Walnut protein hydrolysates were reported to exhibit outstanding capacity in free radical scavenging and lipid peroxidation inhibiting. 36,37 Defatted walnut meal, as the main by-product obtained from the roasted kernels of walnut during oil extraction, is composed of more than 40% proteins with a high essential amino acid content. 38 Moreover, walnut proteins have demonstrated to be a notable source of bioactive peptides with antioxidant, angiotensin-converting enzyme inhibitory. and hypocholesterolemic activities. 37,39 Researchers had purified peptides from walnut protein hydrolysates using gel chromatography to obtain cancer cell growth inhibitory peptides, and these peptides could induce apoptosis in cancer cells. 40
As we all know, eating a handful of walnuts per day or even less may help boost memory, concentration, and the speed at which your brain processes information. In our previous work, we developed a facile and reproducible preparation of antioxidant peptides from walnut protein hydrolysates. 41 Recently, new research indicated that cognitive impairment had been correlated to the accumulation of oxidative damage to lipids, proteins, nucleic acids, and the vulnerability of various neurotransmitters/neurotrophin systems’ activity to oxidative stress. 42 To determine whether the antioxidant peptides prepared from walnut by the method could help improve brain function or support brain health, we used H2O2-induced injury model of PC12 cells to explore the protective effect. Zebrafish (Danio rerio) is not only a popular tropical fish pet but also an important model organism for vertebrate development, genetics, and human biology and disease. 43 And they have recently become a focus of neurobehavioral studies since larvae display neuropathological and behavioral phenotypes that are quantifiable and relate to those seen in man. 44 Thus, zebrafish was used as the model to study the neuroprotective effect of walnut peptide (WP).
Recently, some studies had showed that antioxidant peptides could improve learning and memory ability of mice. 45,46 Learning and memory is one of the most important functions of the brain, which is associated with complex neurophysiologic and neurochemical changes. 47,48 Usually memory processes are divided into 3 stages: memory acquisition, memory consolidation, and memory reproduction. 49,50 As far as we know, Morris water maze and step-down passive avoidance tests are commonly used laboratory tools in behavioral neuroscience. 51 -54 Chemical agents such as scopolamine, sodium nitrite, and ethanol impaired memory in mice trained on Morris water maze and step-down passive avoidance tests, which are used to measure the 3 stages of memory process depending on the drug-treated period. 55 -58 Herein, memory-enhancing effect of WP in mice was assessed by Morris water maze and step-down passive avoidance tests in the present work. At the same time, the acute toxicity was performed to assess its safety profile.
PC12 cells are neuronal cells derived from a cancerous lesion, pheochromocytoma, of the adrenal medulla of the rat. 59 When cultured in vitro, it could perform some growth characteristics similar to neurons, like cell aggregation and emergency of fibrous ridges. 60 Thus, the cells exhibit many characteristics useful for the study of the neurotoxicity, neuroprotection, and neurocognition. 61 H2O2, one of the main reactive oxygen species, is known to elevate oxidative stress, resulting in apoptosis or necrosis of PC12 cells, which was characterized by chromatin margination or condensation, DNA fragmentation, and nuclear collapse. 62,63 Our study was carried out to determine whether WP could protect against H2O2-induced oxidative injury in PC12 cells.
To investigate the WP effect of cell toxicity on PC12 cells, cells were exposed to 0, 1, 10, 30, 100, and 200 µg/mL of WP. Figure 1(a) shows the cell viability of cultured PC12 cells after 24 hours of incubation with different concentrations of WP using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. PC12 cells treated with WP did not exhibit any significant viability or proliferation alterations. As shown in Figure 1(b), cell viability of PC12 cells incubated with 100 µmol/L H2O2 for 12 hours showed significant decrease. The cell viability rate was 57.2% compared to the control. In contrast, pretreatment of the cells with WP (0, 1, 10, 30, 100, and 200 µg/mL) for 6 hours could remarkably restore cell survival to 76.9%. Moreover, the protection rates of WP, according to the respective calculations, are listed in Table 1. These results suggested that WP could be considered as a neuroprotective agent against H2O2-induced oxidative stress.

Cytotoxicity and cytoprotective activity of WP. (a) PC12 cells were pretreated with various concentrations of WP for 24 hours; (b) cell viability of PC12 cells pretreated with various concentrations of WP for 6 hours before exposure to H2O2 (100 µmol/L) for 12 hours. WP, walnut peptide.
The Protection Rates of WP in PC12 Cells.

WP, walnut peptide.
To further validate the neuroprotective effect of WP, zebrafish was used as the model to study the effect. The fluorescent intensity of zebrafish is presented in Figures 2 and 3, and the neuroprotection ratios are listed in Table 2.

The fluorescent intensity of zebrafish in different groups. GSH, glutathione; WP, walnut peptide.

The effect of WP on fluorescent intensity of zebrafish. ### P < 0.001 compared with normal control group, *** P < 0.001 compared with negative control group. GSH, glutathione; WP, walnut peptide.
Neuroprotective Effect of WP in Zebrafish.

GSH, glutathione; WP, walnut peptide.
P < 0.001 compared with the normal control group.
P < 0.001 compared with the negative control group.
As shown in Table 2, the fluorescence intensity of zebrafish in negative control group (565 783) was significantly higher than that of the normal control group (161976), indicating that the model was established successfully. The fluorescence intensity of glutathione (GSH) was 193900, compared with the negative control group (P < 0.001), and the neuroprotection ratio was 92%, which indicated that GSH had an obvious protection effect. The fluorescence intensities of different doses of WP (222, 667, and 2000 µg/mL) were 395 025, 451 259, and 636 837. Compared with the negative control group, their neuroprotection ratios were 42%, 28%, and −18%, proving that as dose increases, the neuroprotective effect became weaker and weaker.
The mechanism of neuroprotective effect was investigated by determining the inhibitory effects of caspases 3/7, 8, and 9. The results of inhibitory effects of caspases 3/7, 8, and 9 are shown in Table 3 and Figure 4. The luminescence intensity (caspase 3/7) of negative control group (12.42 × 106) compared to normal control group (7.17 × 106), P < 0.001 indicated that the model was highly significant and the establishment of model was successful. The luminescence intensity and caspase 3/7 inhibition rate of low-dose WP group were 11.54 × 106 and 17%, while medium-dose WP were 11.23% and 23%, and high-dose WP group were 12.76% and −6%, respectively. The luminescence intensity (caspase 8) of negative control group (5.21 × 106) was significantly higher than that of the normal control group (2.81 × 106), suggesting that the model was successfully established. The luminescence intensity and caspase 8 inhibition rate of medium-dose WP group were 3.87 × 106 and 56%, respectively, and P < 0.005 indicated that the caspase 8 activity was inhibited obviously. Interestingly, low- or high-dose WP group could not inhibit the activity of caspase 8. The luminescence intensity (caspase 9) of normal control group (6.74 × 106) was significantly lower than that of the negative control group (9.80 × 106), indicating that the model was successfully established. The intensity of all WP groups was not lower than that negative control group significantly, indicating that WP could not inhibit the activity of caspase 9. Based on these experimental results, medium dose of WP inhibited the activity of caspases 3/7 and 8, but had no significant effect on the activity of caspase 9.

The luminescence intensity levels of caspases 3/7, 8, and 9. (a) Caspase 3/7, (b) caspase 8, and (c) caspase 9. # P < 0.05 compared with the normal control group, ## P < 0.01 compared with the normal control group, ### P < 0.001 compared with the normal control group, *P < 0.05 compared with the negative control group, **P < 0.01 compared with the negative control group. WP, walnut peptide.
The Inhibitory Effects of WP on Caspases 3/7, 8, and 9 of Zebrafish.

WP, walnut peptide.
P < 0.001 compared with the normal control group, * P < 0.05 compared with the negative control group, ** P < 0.01 compared with the negative control group.
P < 0.05 compared with the normal control group,
P < 0.01 compared with the normal control group.
Meanwhile, the mRNA expression levels of Bax, brain-derived neurotrophic factor (BDNF), and glial cell line-derived neurotrophic factor (GDNF) were also studied. The results are listed in Table 4. As shown in Table 4, the mRNA expression of Bax in negative control group (1.01) was significantly higher than that of the normal control group (0.28), indicating that the model was established successfully. The mRNA expressions of Bax in WP groups (dose: 74, 222, and 667 µg/mL) were 0.23, 0.10, and 1.42, respectively. Compared with the negative control group, the low dose or medium dose of WP could reduce the expression of Bax. However, high dose of WP could increase the mRNA expression of Bax. The mRNA expression of BDNF in negative control group (1.00) was compared with that of the normal control group (1.28), indicating that the establishment of model was successful. The mRNA expressions of BDNF in WP groups (dose: 74, 222, and 667 µg/mL) were 1.14, 1.13, and 0.32, respectively. Compared with the negative group, the mRNA expression of BDNF was improved by the low or medium dose of WP. Instead, the high dose of WP could reduce the expression of BNDF. The mRNA expression of GDNF in negative group (1.00) was compared with that of the negative group (0.39), indicating that the model was successfully established. The mRNA expressions of GDNF in WP groups (dose: 74, 222, and 667 µg/mL) were 0.94, 0.68, and 0.46, respectively, proving that WP could reduce the mRNA expression of GDNF. These results were identical with the previous study. Based on the above results, WP could be considered as a neuroprotective agent.
The mRNA Expression Levels of Bax, BDNF, and GDNF in Zebrafish.

BDNF, brain-derived neurotrophic factor; GDNF, glial cell line-derived neurotrophic factor; WP, walnut peptide.
P < 0.001 compared with the normal control group, * P < 0.05 compared with the negative control group, *** P < 0.001 compared with the negative control group.
P < 0.01 compared with the normal control group.
Besides, the present study was to examine whether WP prepared by our method may influence brain function in mice. To do this, Morris water maze and step-down tests in mice were carried out.
Since scopolamine is known to show rapid antidepressant effects in humans, and impair short-term memory and learning ability, it has been used for the Alzheimer’s disease animal model. 64,65 Therefore, the behavioral memory improvement of WP in scopolamine-induced impairment in memory acquisition in mice was examined. The results are summarized in Table 5.
Effect of WP on the Escape Latency of Scopolamine-Induced Acquisition Impairment in the Morris Water Maze Test.

WP, walnut peptide.
P < 0.01 compared with the normal control group, * P < 0.05 compared with the negative control group, ** P < 0.01 compared with the negative control group.
As shown in Table 5, over 5 training days for place navigation, the escape latency decreased over the course of training trials in all groups, which indicated that learning and memory functions of mice improved with extension. The negative control group showed marked retardation in the escape, indicating that intraperitoneal injection of scopolamine would significantly induce learning and memory deficits in mice. The results of multivariate analysis of variance showed that there was significant difference in daily escape latency among groups. Nmodipine as the positive control could greatly reduce escape latency of scopolamine-treated amnesia mice. The treatment of mice with medium dose (100 mg/kg) or high dose (300 mg/kg) of WP shortened the latency, compared to the negative control group. However, there was no significant difference in latency between low dose (30 mg/kg) of WP and negative control groups.
In the spatial probe test, negative control group had a significantly lower target time and less crossing times, compared to the normal control group (Supplemental Table S1). It was interesting that administration of nmodipine and WP exhibited much longer target time and more crossing times. Thus, nmodipine and WP treatment could improve greatly memory acquisition impaired by scopolamine.
Sodium nitrite significantly impaired memory of mice, which is also supported by the earlier study. 49 Hypoxia induced with sodium nitrite reduces incorporation of choline into acetylcholine, thus decreasing the synthesis of acetylcholine and hence lead to cognitive deficits. 66 Therefore, sodium nitrite was used to induce impairment of memory consolidation in mice in the present work.
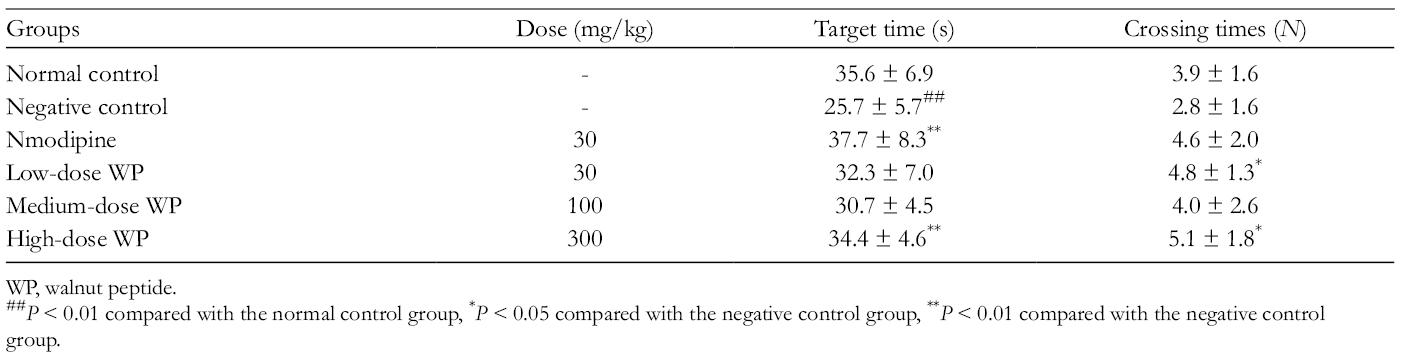
Over 4 consecutive training days for place navigation in the Morris water maze experiment, the mice were injected with 90 mg/kg sodium nitrite solution subcutaneously. After 24 hours, the escape latency was prolonged over the course of training trials in all groups (Supplemental Table S2). It suggested that the treatment of mice with sodium nitrite would lead to impairment in spatial learning and memory. Interestingly, the treatment of mice with high dose of WP exhibited shorter escape latency than that of negative control and positive groups, indicating that WP intervention would inhibit the impairment by sodium nitrite and promote active spontaneous motion. It can be seen from Table 6 that the negative control group had less crossing times and lower target time, compared to the normal control. In high dose of WP group, the target time and crossing times were similar to those in the nmodipine group and significantly more than those in the negative control group. Therefore, these results revealed that high dose of WP greatly reversed sodium nitrite-induced impairment of memory consolidation.
Effect of WP on the Target Time and Crossing Times of Sodium Nitrite-Induced Consolidation Impairment in the Morris Water Maze Test.
WP, walnut peptide.
P < 0.01 compared with the normal control group, * P < 0.05 compared with the negative control group, ** P < 0.01 compared with the negative control group.
Ethanol is able to alter cognitive and behavioral performance in both humans and laboratory animals. 67 The Morris water maze with ethanol-intoxicated mice is one of the most frequently employed methods for evaluating memory-enhancing effects in vivo. 68 The present study was undertaken to investigate the effect of WP on impairment of memory reproduction induced by ethanol in mice. The results obtained are presented in Table 7.
Effect of WP on the Escape Latency of Ethanol-Induced Reproduction Impairment in the Morris Water Maze Test.

WP, walnut peptide.
P < 0.01 compared with the normal control group, ** P < 0.01 compared with the negative control group.
As shown in Table 7, 40% ethanol administration notably prolonged the escape latency of the mice in all groups. Significant differences were observed between the high-dose WP group and others. The treatment of mice with high dose of WP obviously cut down the escape latency as compared with the other groups, which suggested that high dose of WP could reverse the ethanol-induced memory deficits in mice. The target time and crossing times were remarkably shorter and lower in the negative control mice than those in the normal control mice (Supplemental Table S3). By contrast, nmodipine and high dose of WP could prolong and increase the target time and crossing times markedly. Interestingly, no significant differences appeared on the escape latency, target time, and crossing times in the WP (30 and 100 mg/kg) groups as compared with the negative group. Therefore, by treatment with high dose of WP, learning and memory performances were manifestly improved.
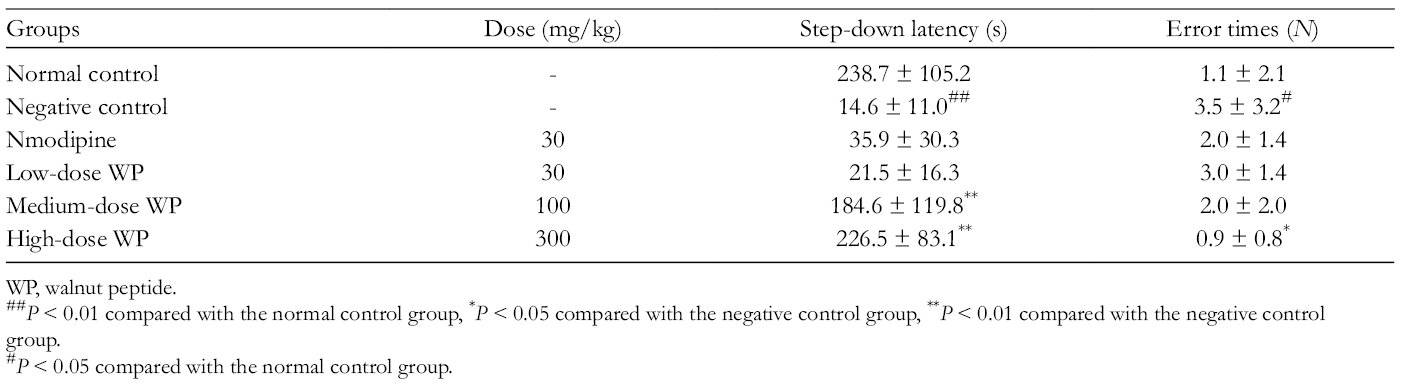
After 7 days of continuous administration, step-down type test was used to determine the effect of WP on impairment of memory acquisition induced by scopolamine. The results are summarized in Table 8. The present results showed that the step-down latency was decreased in scopolamine-treated mice as compared to control animals, which indicated that scopolamine caused impairments on memory retention. The mice treated with medium dose or high dose of WP prolonged the step-down latency markedly, compared to the other groups. In addition, much less error responses were found in mice treated with high dose of WP. Therefore, these findings revealed that preadministration of WP was effective in preventing this memory deficit in this task. It could be concluded that treatment with high dose of WP could attenuate the effect of scopolamine on memory acquisition significantly.
Effect of WP on Scopolamine-Induced Acquisition Impairment in the Step-Down Test.
WP, walnut peptide.
P < 0.01 compared with the normal control group, * P < 0.05 compared with the negative control group, ** P < 0.01 compared with the negative control group.
P < 0.05 compared with the normal control group.
Effect of WP on sodium nitrite-induced impairment in memory consolidation in mice was also investigated using the step-down type passive avoidance task. The mice treated with sodium nitrite exhibited shorter step-down latency and more error responses than those of the normal control group, which indicated that sodium nitrite caused impairments on memory retention (Supplemental Table S4). However, of greater significance was the finding that longer latency and less error responses were found in mice treated with WP and nmodipine, and the levels of latency and error times were similar to the ones of normal control group. Taken together, these data suggested that WP ameliorates the impairment effect of sodium nitrite on learning and memory deficits.
To further confirm the effect of WP on learning and memory functions, the behavioral memory improvement of WP in ethanol-induced impairment in memory reproduction in mice was examined. The step-down latency of negative control group was shorter than that of the normal control group, and the negative control group exhibited more error numbers (Supplemental Table S5). In contrast, the latency was significantly longer and the number of errors was significantly lower in the WP and nmodipine groups. These behavioral results revealed that preadministration of WP could offer protection against ethanol-induced memory reproduction impairment.
Toxicity tests are experimental screening methods that are used to confirm the safety of food or drugs in assays with animal models. Mortality is an important criterion on toxicological evaluation. The results obtained in our acute oral toxicity study showed no mortality at the dose evaluated (20.1 g/kg body weight [BW]). Thus, it is suggested that the oral lethal dose (LD50) of WP is greater than 20.1 g/kg BW. These animals were found to be normal in appearance with no changes in their skin, fur, eyes, and mucous membranes. They showed neither behavioral changes such as lethargy or altered sleep patterns nor any observable signs of toxicity such as convulsion, tremors, diarrhea, or coma.
The increase or decrease in BW can indicate significant physiological changes such as hormonal variations, liver disorders, and decreased absorption of proteins, amino acids, and other nutrients. The BW of mice (presented in Figure 5) in each group showed a steady increasing trend, while WP-treated groups had a higher body mass index than that of the control group. However, statistical analysis of BW gain did not reveal any significant difference between the treated and the control groups.
Body weight of mice treated orally with WP (20.1 g/kg body weight). WP, walnut peptide.
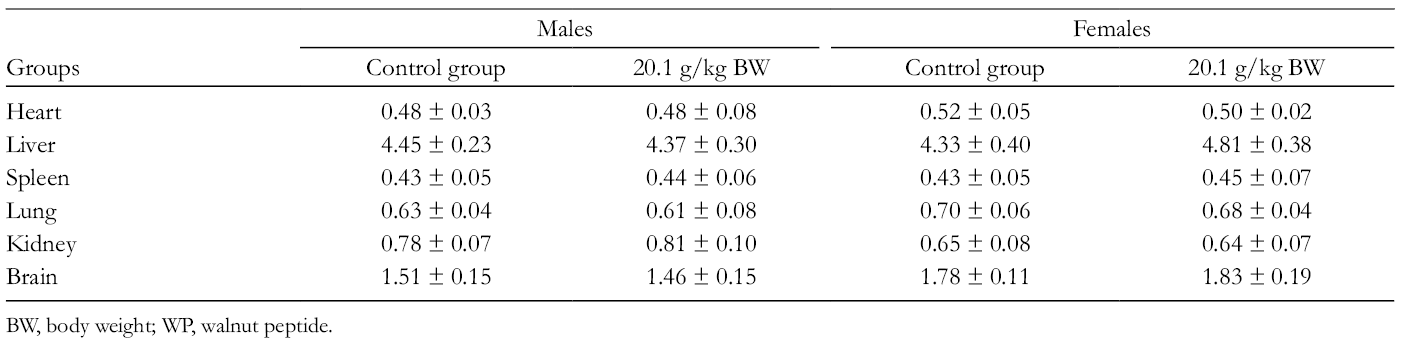
Relative organ weight is a more specific parameter than absolute weight in toxicity evaluation in animal models. 69 The effects of WP on the relative organ weights of mice are presented in Table 9. As shown in Table 9, the oral administration of WP caused no significant changes in relative weight of the organs (heart, liver, spleen, lung, kidney, and brain) in the treated ones compared to the control mice of both sexes. The results demonstrate that the essential organs were not adversely affected during the treatment.
Relative Organ Weights of Mice Receiving WP for 14 days.
BW, body weight; WP, walnut peptide.
In the present work, the neuroprotective and memory-enhancing effects of WP were studied in vitro and in vivo. Results revealed that WP was able to protect cultured PC12 cells against H2O2-induced oxidative stress. Zebrafish was used as the model to study the neuroprotective effect of WP. WP had an obvious effect, and the neuroprotection ratio reached 42% at 222 µg/mL. The mechanism study indicated that the medium dose of WP could inhibit the activity of caspases 3/7 and 8, reduce the mRNA expression levels of Bax and GDNF, and improve the mRNA expression level of BDNF significantly. Meanwhile, WP could ameliorate learning and memory impairments using the Morris water maze and the step-down passive avoidance tests in mice. In addition, the acute toxicity study indicated that administration of WP at the dose of 20.1 g/kg BW to mice induced no mortality or treatment-related adverse effects with regard to BW, general behavior, or relative organ weights. Thus, WP is nontoxic and safe by oral intake. Based on the results of these in vitro and in vivo studies, further studies can be carried out to explore the other possible mechanisms for memory-improving effect of WP and its usefulness in the management of cognitive disorders.
Experimental
Materials and Chemicals
Walnuts (Juglans regia L.) were collected from Aksu city, Xinjiang province, in China, in May 2015. Prof Huangching Lin, Institute of Pharmacy, Taiwan National Defense Medical Center, identified the plant material. A voucher specimen (No. 150 522) was deposited at R&D Center, Sinphar Tian-Li Pharmaceutical Co., Ltd. Food-grade protease (neutrase) (≥600 units/mg) were obtained from Guangxi Pangbo Biothech Co., Ltd. Reagents (sodium hydroxide and hydrochloric acid) were of food grade, and used without further purification unless otherwise noted. Dulbecco’s modified Eagle’s medium (DMEM), fetal bovine serum (FBS), and trypsin (1:250) were purchased from Gibco BRL, Life Technologies (New York, USA). MTT was purchased from Ameresco (Massachusetts, USA).
Preparation of WPs
WP was prepared according to a previously described method. 41
Analysis of Cell Viability by MTT Assay
The rat pheochromocytoma (PC12) cell line was obtained from the Cell Institute of Biochemistry and Cell Biology (Shanghai, China), and cultured in DMEM supplemented with 10% heat-inactivated FBS. The cells were maintained at 37°C in a humidified 5% carbon dioxide and 95% air incubator. The culture media were changed every 2 days. When the cells reached subconfluence, they were treated with trypsin, after which they were seeded into 96-well plates at a density 1 × 104 cells/well/100 µL for 24 hours.
WP was dissolved in dimethyl sulfoxide (DMSO), and then the peptides at different concentrations were added into wells. The cells were treated for 6 hours, followed by co-culture with 100 µmol/L H2O2. After treated for 12 hours, 10 µL of MTT (5 mg/mL, in PBS) was added into each well and cultured for another 4 hours. Supernatant was removed and the cells were lysed in 150 µL of DMSO. The plate was shaken for 10 minutes, and the optical density (OD) values were detected by a microplate reader using a detective wavelength of 570 nm, with a reference wavelength of 650 nm. The following formula was used: Cell viability (%) = absorption value experimental group/absorption value of control group × 100%. Cell viability was expressed as a percentage with the control cells treated with vehicle as 100%.
In Vivo Neuroprotective Effect of WP in Zebrafish
Fish cultivation
The wild-type AB strain of zebrafish was obtained from Hangzhou Hunter Biotechnology Co., Ltd., Hangzhou, China (Certification No. SYXK 2012-0171, Hangzhou, China). Zebrafish embryos were generated by natural pairwise mating. All fishes were raised and kept under standard conditions at 28°C in aerated water (200 mg instant ocean sea salt per liter; conductivity 480-510 μS/cm; pH 6.9-7.2; hardness 53.7-71.6 mg/L CaCO3), on a 14-hour light and 10-hour dark cycle. All experimental procedures involving fish conform to AAALAC standards and were approved by institutional IACUC committees.
Evaluation of neuroprotective effect
Zebrafishes at 24 hours postfertilization (hpf) were placed into a 6-well microplate at a density of 30 zebrafishes per well, and treated with mycophenolate mofetil, in order to induce nerve injury of zebrafishes. Zebrafishes were divided into 6 groups (n = 30 in each group): normal, control, GSH (154 µg/mL, positive control group), WP 222 (222 µg/mL), WP 667 (667 µg/mL), WP 2000 (2000 µg/mL). GSH and WP were dissolved in aerated water. The groups except normal group were treated with mycophenolate mofetil (0.25 μmol/L). After 24 hours, the fishes were stained by acridine orange. And then, 10 stained zebrafishes from each experimental group were selected randomly to photograph under fluorescence microscope, analyzing and counting the fluorescent intensity.
Caspases 3/7, 8, and 9 activation assays
Zebrafishes at 24 hpf were placed into a 6-well microplate at a density of 30 zebrafishes per well, and treated with mycophenolate mofetil. Zebrafishes were divided into 5 groups (n = 30 in each group): normal, control, WP 74 (74 µg/mL), WP 222 (222 µg/mL), WP 667 (667 µg/mL). WPs were dissolved in aerated water. The groups except normal group were treated with mycophenolate mofetil (0.25 μmol/L). After 24 hours, the percentage of caspases 3/7, 8, and 9 activation was quantified using the Caspase-Glo 3/7, 8, and 9 kit following the manufacturer’s instructions. This kit used a luminogenic substrate that was cleaved by caspases resulting in the generation of the luminescent signal. This signal was measured with a microplate reader.
RNA extraction and generation of cDNA
Zebrafishes at 24 hpf were placed into a 6-well microplate at a density of 30 zebrafishes per well, and treated with mycophenolate mofetil. Zebrafishes were divided into 5 groups (n = 30 in each group): normal, control, WP 74 (74 µg/mL), WP 222 (222 µg/mL), WP 667 (667 µg/mL). WPs were dissolved in aerated water. The groups except normal group were treated with mycophenolate mofetil (0.25 μmol/L). After 24 hours, total RNA was extracted using Trizol Reagent (Invitrogen, USA) following the manufacturer’s protocol. Both RNA purity and fidelity were checked by evaluation of OD (calculation of 260/280 nm ratio). The first-strand cDNA was generated from 2 µg of total RNA by using FastQuant RT Kit (Tiangen, China), and was stored at −20°C. β-Actin was used as the internal control. The mRNA expression was determined by using the 2-ΔΔCT method. The sequences of primers are demonstrated in Table 10.
Indicates the Sequences of the Employed Primers in This Study.

BDNF, brain-derived neurotrophic factor; GDNF, glial cell line-derived neurotrophic factor.
In Vivo Memory-Enhancing Effect of WP in Mice
Experimental animals
Male ICR mice (weighing 18-22 g) were originally obtained from Qinglongshan Animal Breeding Center (Certification No. SCXK 2017-0001, Nanjing, China). They were housed in a monitored environment (temperature 20∘C ± 2∘C, humidity 50% ± 10%, 12:12 hours light-dark cycle). The animals were euthanized by cervical dislocation. All procedures described were approved by the Animal Ethics Committee of School of Chinese Materia Medica, China Pharmaceutical University.
Morris water maze test
The Morris water maze test was used to evaluate the effect of WP on learning and memory according to the method described previously. 70 The Morris water maze, a large circular pool (90 cm in diameter and 30 cm in height), was filled with water (22-24∘C). The mice (male) were randomly divided into 6 groups (n = 10 in each group): normal, control, nmodipine (30 mg/kg, positive control group), WP 30 (30 mg/kg), WP 100 (100 mg/kg), WP 300 (300 mg/kg). The WP and nmodipine were dissolved in purified water, and administered orally once a day for 7 consecutive days. The normal and control groups were administered with an equal volume of purified water.
Scopolamine-induced impairment in memory acquisition in mice
Before the test trials, mice were pretrained over 5 consecutive days (2 trials per day) from the eighth day of administration of drugs. Thirty minutes after the drug administration, scopolamine (2 mg/kg) was injected intraperitoneally to control, nmodipine, and WP groups to induce memory impairment, while the same volume of saline was orally administered to normal group, 10 minutes prior to behavioral tests. The mice were given for 90 seconds to find the hidden platform. If a mouse failed to find the platform in 90 seconds, it was placed on it for 15 seconds. The mice of every group were trained for 4 consecutive days, and the escape latency (s) was recorded. On the fifth day of behavioral measurement, the platform was removed, and each mouse was placed into the water at the same incidence point. They were allowed to swim freely for 90 seconds. The crossing numbers over the position at which the platform had been located, and the target time (s, the swimming time spent in the target quadrant) were recorded as measure for spatial memory.
Sodium nitrite-induced impairment in memory consolidation in mice
Before the test trials, mice were pretrained over 5 consecutive days (2 trials per day) from the eighth day of administration of drugs. The mice were placed into the pool containing platform, and the escape latency (s, finding the platform) was recorded. The mice were given for 90 seconds to find the hidden platform. If a mouse failed to find the platform in 90 seconds, it was placed on it for 15 seconds. The mice of every group were trained for 4 consecutive days. Following the last training trial, the mice were injected with 120 mg/kg sodium nitrite solution subcutaneously with a view to induce memory impairment, with the exception of the saline control group, which was injected with the same volume of normal saline. After 24 hours, the crossing numbers over the position at which the platform had been located were recorded. At the same time, the escape latency (s) and the target time (s, the swimming time spent in the target quadrant) were recorded.
Ethanol-induced impairment in memory reproduction in mice
Before the test trials, mice were pretrained over 5 consecutive days (2 trials per day) from the eighth day of administration of drugs. Thirty minutes after the drug administration, the mice were placed into the pool containing platform, and the escape latency (s, finding the platform) was recorded. The mice were given 90 seconds to find the hidden platform. If a mouse failed to find the platform in 90 seconds, it was placed on it for 15 seconds. The mice of every group were trained for 4 consecutive days. On the fifth day of behavioral measurement, the mice were administered 0.1 mL/10 g 40% (v/v) ethanol with a view to induce memory impairment, with the exception of the saline control group, which was injected with the same volume of normal saline. The crossing numbers over the position at which the platform had been located were recorded. Moreover, the escape latency (s) and the target time (s, the swimming time spent in the target quadrant) were recorded.
Step-down test
Step-down latency in passive avoidance is a proper test for memory retention and retrieval in mice. 71 The test was performed according to a previously described method. 72 The floor of the chamber consisted of copper rods and a well-insulated platform in 1 corner of the chamber. Initially, the mice were placed on the insulated platform. When they stepped down with all the 4 paws onto the floor (error trial), they would receive an electric foot shock. Thus, the mice were trained not to step down from the insulated platform onto the floor to avoid the electric foot shock. The mice (male) were randomly divided into 6 groups (n = 10 in each group): normal, control, nmodipine (30 mg/kg, positive control group), WP 30 (30 mg/kg), WP 100 (100 mg/kg), WP 300 (300 mg/kg). The WP and nmodipine were dissolved in purified water, and administered orally once a day for 7 consecutive days. The normal and control groups were administered with an equal volume of purified water.
Scopolamine-induced impairment in memory acquisition in mice
The experiment was performed on the seventh day of administration of drugs. One hour after the drug administration, scopolamine hydrobromide (3 mg/kg) was injected intraperitoneally to control, nmodipine, and WP groups to induce memory impairment, while the same volume of saline was orally administered to normal group, 10 minutes prior to behavioral tests. Each mouse was kept in the box for 3 minutes to adapt to the environment. Then, the mouse was gently placed on the platform. When the electrical currents (36 V) were delivered, the mice would react by jumping onto the platform to avoid the electric shock, and the electric currents were maintained for 5 minutes. The retention test was carried out 24 hours after the training test specified above. The step-down latency (s, duration of staying on the platform) and the number of errors (the mouse steps down from the platform and gets a shock) within 5 minutes were recorded.
Sodium nitrite-induced impairment in memory consolidation in mice
The behavioral tests were carried out on the seventh day of administration of drugs. Each mouse was kept in the box for 3 minutes to adapt to the environment. Then, the mouse was gently placed on the platform. When the electrical currents (36 V) were delivered, the mice would react by jumping onto the platform to avoid the electric shock, and the electric currents were maintained for 5 minutes. Then, the mice were injected with 90 mg/kg sodium nitrite solution subcutaneously with a view to induce memory impairment, with the exception of the saline control group, which was injected with the same volume of normal saline. After 24 hours, the retention test was carried out. The step-down latency (s, duration of staying on the platform) and the number of errors (the mouse steps down from the platform and gets a shock) were recorded.
Ethanol-induced impairment in memory reproduction in mice
The experiment was performed on the seventh day of administration of drugs. We administered 0.1 mL/10 g 40% (v/v) ethanol to control, nmodipine, and WP groups to induce memory impairment, while the same volume of saline was orally administered to normal group, 30 minutes prior to behavioral tests. Each mouse was kept in the box for 3 minutes to adapt to the environment. Then, the mouse was gently placed on the platform. When the electrical currents (36 V) were delivered, the mice would react by jumping onto the platform to avoid the electric shock, and the electric currents were maintained for 5 minutes. The retention test was carried out 24 hours after the training test specified above. The step-down latency (s, duration of staying on the platform) and the number of errors (the mouse steps down from the platform and gets a shock) within 5 minutes were recorded.
Acute Toxicity Study
The experiment was conducted according to the protocols described by the guideline for testing of drugs (Technical Standards for Testing & Assessment of Health Food). Drinking water and food were provided ad libitum throughout the experiment, except for the short fasting period where the drinking water was still in free access but no food was provided 16 hours prior to treatment. Forty mice (half male and half female) were allocated into 2 groups of 20 animals each (10 males and 10 females). The treatment group was administered the WP at a single dose of 20.1 g/kg BW, whereas the control group received an equal volume of water. Both treatment were administered through oral gavage using a volume of 0.6 mL. Surviving animals were weighed and visual observations for mortality, behavioral pattern, changes in physical appearance, injury, pain, and signs of illness were conducted daily for a period of 14 days. At the end of the 14 days, all the mice were killed. Vital organs such as heart, liver, spleen, lung, kidney, and brain were quickly removed. All of the individual organs were weighed and their features were compared between both treated and control groups. Relative organ weight was calculated as (weight of organ/BW of mouse on the day of killing) ×100%.
Statistical Analysis
All statistical analyses were performed using SPSS 10.0, and the data were analyzed using one-way analysis of variance. Values are expressed as the mean ± standard error of mean (SEM). The mean separations were performed using the least significant difference method.
Supplemental Material
Supplementary data - Supplemental material for Neuroprotective and Memory-Enhancing Effects of Antioxidant Peptide From Walnut (Juglans regia L.) Protein Hydrolysates
Supplemental material, Supplementary data, for Neuroprotective and Memory-Enhancing Effects of Antioxidant Peptide From Walnut (Juglans regia L.) Protein Hydrolysates by Mingchuan Liu, Shengjie Yang, Jinping Yang, Yita Lee, Junping Kou, and Chaojih Wang in Natural Product Communications
Footnotes
*
The authors Mingchuan Liu and Shengjie Yang contributed equally to the work.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.