
Abstract
As the main by-product during the oil production of peony seeds, the episperm is traditionally used as a lead component in folk herbal formulas for the cancer treatment in China. However, the investigation of its phytochemical foundation underlying anticancer effects remains an ongoing challenge. The work therefore determined growth inhibition activities of 8 solvent extracts of peony episperms in the human liver cancer cell line. This activity was then mapped onto the secondary metabolite profile of extracts by principal components analysis (PCA). The top 3 principal components of High Performance Liquid Chromatograph (HPLC)-PCA map discriminated extract activities mainly based on the differential content of 5 stilbene compounds, which were then tested individually. The trans-ε-viniferin, gnetin-H, suffruticosol A, suffruticosol B, and suffruticosol C were thus determined as growth inhibitors and apoptosis inducers of human liver cancer cells with activities comparable to that of the antineoplastic cisplatin. A partial least squares regression-HPLC model was also constructed for the prediction of inhibitory effects of peony episperm extracts. These results expand the fundamental understanding of the peony episperms and support its current medicinal uses in China. Moreover, PCA-mediated secondary metabolite mapping was proved to be an efficient approach to qualify biomedical products required for pharmaceutical and medicinal uses.
The peony seed (the kernel of Paeonia ostii or Paeonia rockii T. Hong and J.J. Li), as a valuable healthy food, has attracted great attention since its oil authorized as a new food resource by the Ministry of Health of the People’s Republic of China in March 2011. 1 The peony episperm, the seed coat of tree peony, is one of the most important by-products during the preparation of peony seed oil. It is claimed that the peony episperm was used in many folk Traditional Chinese Medicine (TCM) formulas for the treatment of cancer-related symptoms in China. 2 However, the scientific evidence of this traditional medicinal use is sparse. A fundamental variable in evaluating such integrated therapy is the chemical profile of the plant material, a point usually represented through qualification and quantification of certain abundant compounds within the complex natural product and their clinical formulations. Generally, this could often be confounded by cultivation differences and conflicting results from published pharmacological activities of compounds that are not relevant to clinical use. 3
Concerning peony seeds, the polysaccharides, monoterpene glycosides, and stilbenes have been reported as bioactive compounds in the previous phytochemical studies. 4 -7 Especially, the polysaccharides and stilbene resveratrol oligomers isolated from peony seeds were found to behave with great anticancer activities either on cancer cell lines in vitro or in cancerous animal models in vivo. 7 -9 The peony seed polysaccharides presented remarkable inhibitory effects on the growth of prostate cancer (PC-3), colon cancer (HCT-116), breast cancer (MCF-7), and cervical cancer (Hela) cells. It arrested cell cycles at G0/G1 stage through regulating expression of cyclin A/B1/D1/E1 and CDK-1/2/4/6. Moreover, cancer cells were induced into apoptotic progress through the modulation of cytochrome c, Bax, Bcl-2, and caspase proteins. 7 Two resveratrol oligomers from the peony seed, cis- and trans-gnetin H, inhibited proliferation of lung cancer (A549), breast cancer (BT20 and MCF 7), and osteosarcoma (U2OS) through arresting cell cycles and then inducing apoptotic death. Moreover, cis-gnetin H could suppress the growth of xenograft lung tumors in mice models. 9 The cis- and trans-suffruticosol D were reported to selectively inhibit growth of human lung cancer cells A549, breast cancer cells BT20 and MCF-7, and osteosarcoma cells U2OS, but not normal human breast cells HMEC and lung cells HPL1A. In lung cancer cells, cis- and trans-suffruticosol D could provoke oxidative stress, induce apoptosis, decrease the mitochondrial membrane potential, inhibit cell motility, and block the NF-kappa B pathway. 8 Taken together, the reports above have partly provided scientific evidences for anticancer compounds of peony seeds. However, the peony episperm, a herb used in the anticancer formulae of TCM, has not been systematically investigated. The active compounds related to its anticancer uses are still unknown.
The chemical fingerprint, generated by either High Performance Liquid Chromatography (HPLC) or Nuclear Magnetic Resonance spectroscopic data from the permutation and combination of multiple chemical compounds, could confer the unique pattern to each herbal samples. 10 However, since the data involved are too numerous and complex to analyze manually, the statistical and software learning approaches are needed to extract useful information and biological insights. 11 Multivariate data analysis (MVA) has thus been employed for the metabolic profiling of herbal secondary metabolites in many studies. 12,13 The commonly employed approaches are principal components analysis (PCA) and partial least squares (PLS) analysis. 12 These approaches, applied recently in the analysis of the herbal preparation, provided more precise information about active components responsible for pharmaceutical effects via PCA loading plots and distribution analysis. 13,14 Nevertheless, the PLS-mediated approach constructed the chemical-pharmaceutical activities’ models which can predict the pharmacological activity of herbal preparations based on the chemical patterns alone. 13,15
In this study, to clarify the phytochemical foundation of peony episperm with regard to its anticancer usages in China, HPLC secondary metabolite maps were constructed to correlate the pure compounds with growth inhibitory activity of crude extracts on human hepatoma cell lines. It is thus expected to reveal the functional anticancer compounds of peony episperm.
Materials and Methods
Cell Lines and Sample Preparation
The human hepatoma cell line HepG2 was kindly provided by School of Medicine, Henan University of Science and Technology. Cells were grown in Dulbecco modified Eagle medium medium (Solarbio, Beijing, China) supplemented with 10% fetal bovine serum, 100 g/mL penicillin, and 100 g/mL streptomycin in a 5% carbon dioxide humidified incubator at 37°C. Experiments were performed when cells were approximately 80% confluent.
The peony seeds (Paeonia rockii) were purchased from the local peony Cultivation Base (Luoyang Tuqiao flower and seeding Co. Ltd., Luoyang, China), and the identification of original plant P. rockii was authenticated by Professor Houxiao Gai (Agricultural School, Henan University of Science and Technology). The seeds were then directly collected from the mature plants. The voucher specimens have been deposited in natural products research laboratory, School of Chemical Engineering and Pharmaceutics (access number: JGG2017-026).
The episperms were removed from the peony seed by hand and then minced. Portions of minced materials (10 g) were each extracted with 100 mL of different solvents: 45%, 70%, or 100% EtOH and extracted by shaking for 24 hours at room temperature. The solvents were completely removed and the residue was freeze-dried to produce 1.56, 1.36, and 1.25 g of 45%, 70%, and 100% EtOH extracts, respectively. Meanwhile, minced materials (200 g) were extracted with 2000 mL methanol twice. The extracts were filtered, and the solvent was completely removed to produce 41.4 g dry extracts. The dry residues were then redissolved in 100 mL 10% methanol and sequentially extracted twice with 200 mL of petroleum ether, hexane, ethyl acetate, and butanol. The remaining water phase was retained. The fractionation solvents were then completely removed and the residues were lyophilized to produce 29.72, 0.6, 7.44, 1.02, and 1.5 g dry petroleum ether, hexane, ethyl acetate, butanol, and water extracts, respectively. The stilbenes trans-ε-viniferin, gnetin-H, suffruticosol A, suffruticosol B, and suffruticosol C were isolated from the peony episperms as previously reported and kept in vacuum storage to avoid oxidation throughout the whole experiment. 16 The chemical structures were shown in Figure 1.

Chemical structures of trans-ε-viniferin, gnetin-H, suffruticosol A, suffruticosol B, and suffruticosol C.
Cytotoxicity Assay
The 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT, Solarbio, Beijing, China) assay was performed as described below. Briefly, 200 µL cell suspensions were seeded at a concentration of 1.5 × 104 cells/mL in a 96-well plate. After overnight incubation, peony episperm extracts, trans-ε-viniferin (100-25 μg/mL), gnetin-H (100-3.1 μg/mL), suffruticosol A (250-125 μg/mL), suffruticosol B (150-25 μg/mL), and suffruticosol C (150-25 μg/mL) were added in 10 µL aliquots. In this work, the systematic dilution for compounds was not used as different compounds presented greatly different levels of cytotoxicity against cancer cells. The dilution ranges for each compound were determined through several preliminary experiments. The current dilution could present the best cytotoxic tendency and thus gave the precise IC50 values. The negative control was 10 µL of 0.01% Dimethyl Sulphoxide (DMSO) in Phosphate Buffered Saline solution. After 24 hours incubation, 20 µL MTT solution was added and incubated at 37°C for 4 hours. The optical density (O.D.) was measured at 590 nm using a Multiskan Spectrophotometer. The growth inhibition was determined using the following formula: Growth inhibition percentage = ((control O.D. − sample O.D.)/control O.D) × 100%.
HPLC Instrumentation
The chemical profiles of herbal extracts and stilbenes standards were performed using a Waters series HPLC system comprising a Waters e2695 Separations Module with degasser, a variable wavelength 2489 UV/Vis detector, and an autosampler. The UV detector was set at 230 nm and the column used was a Thermo C18, 5 µm (250 × 4.6 mm). Gradient elution of samples and standards were performed using 0.1% formic acid (eluent A) and methanol (eluent B). The initial conditions of gradient elution were 5% of eluent B with linear gradient to 34% from 1 to 15 minutes, to 56% from 15 to 33 minutes, and to 60% from 33 to 45 minutes, this proportion was then returned to the initial condition at 50 minutes. The flow rate was 1 mL/min. Dry residues of the peony episperm extracts were dissolved in methanol at 5 mg/mL. The sample injection volume was 10 µL: 3 injections were performed for each sample and standard.
HPLC Quantitation of trans-ε-Viniferin, Gnetin-H, Suffruticosol A, Suffruticosol B, and Suffruticosol C
The calibration curves of trans-ε-viniferin, gnetin-H, suffruticosol A, suffruticosol B, and suffruticosol C were obtained by preparing stock solutions (500 µg/mL) in methanol. Subsequent dilutions were made to give 500, 400, 300, 200, 100, and 50 µg/mL. Ten microliters of each final concentration was injected in triplicate. The linearity of the calibration curves was determined (R2 >0.994). The intraday precision was calculated by comparing the area of standard obtained for 3 injections of standards within the day. For the interday precision, the area of standard in the calibration curve for 3 days was summed and compared.
Multivariate Data Analysis
Principal components analysis and PLS analysis were performed using SIMCA-P 16.0 MVA software (Umetrics, Sweden). The multiple linear regression models were constructed to predict values for IC50 values. HPLC chromatograms of the peony episperms extracts at 230 nm comprised 3001 discrete regions by data acquisition every second from 0.00 to 50.00 minutes. The resulting data were formatted into Microsoft Excel 2010 and then exported into SIMCA-P 16.0 for analysis. Three principal components were chosen for PLS analysis. The process of generating a PLS model on randomly chosen samples followed by validation was repeated in order to ensure all extracts were excluded once.
Apoptosis Assay
The HepG2 cell suspension (800 µL) was seeded at a concentration of 1.5 × 104 cells/mL in a 24-well plate with 30 mm cover slide plated in the bottom of the well. After overnight incubation, the trans-ε-viniferin, gnetin-H, suffruticosol A, suffruticosol B, and suffruticosol C (IC50 concentration) were added in 40 µL aliquots. The negative control was 40 µL 0.01% DMSO in Hank’s solution. Camptothecin (50 µg/mL) was used as positive control. The cell-seeded cover slides were removed from 24-well plates after 24 hours treatment. The cells were then stained using the Annexin-V-FLUOS staining kit (Roche Applied Science, Mannheim, Germany). The apoptotic cells were visualized by green fluorescence microscopy with excitation wavelength at 488 nm.
Statistical Analysis
The data were shown as mean ± standard deviation. Statistical comparison among treatments was carried out using one-way analysis of variance. The statistical significances between control and sample groups were calculated by the Student's t-test. Data were taken as significant where P < 0.05.
Results
HPLC-PCA Map and PLS Prediction
The PCA scores plot constructed from the HPLC data of crude extracts was presented in Figure 2. Each point represented 1 HPLC UV absorption dataset for a given extract and each extract was tested in triplicates. The samples with similar composition formed clusters, while distinct samples were separated in the scores plot. As illustrated by the ellipses superimposed in Figure 2, all 24 samples could be discriminated into 3 groups. This was consistent with experimental IC50 values of each sample’s inhibitory effects on human liver cancer cell HepG2 in vitro. Crude extracts with IC50 values (<72 µg/mL) were considered as strong inhibitors: 45%, 70%, and 100% aqueous EtOH, petroleum ether, and ethyl acetate extracts. Those of hexane and butanol showed intermediate inhibition (85 < IC50 < 90 µg/mL). The weak inhibition (IC50 > 272 µg/mL) was located in the crude extract from water. This map demonstrated good correlation between the composition and the inhibition. Furthermore, as the variance was mainly due to the top 3 principal components (PC1, 40.6%; PC2, 30.8%; and PC3, 16.1%), it could determine the contributing variables (HPLC peaks) using the PCA contribution scores plot (data not shown). Peaks with retention times of 19.7 minutes (suffruticosol A), 21.1 minutes (suffruticosol B), 24.8 minutes (suffruticosol C), 31.1 minutes (trans-ε-viniferin), 34.9 minutes (unknown), and 35.3 minutes (gnetin-H) contributed most to PC1-3, as confirmed by spiking samples with standards previously isolated. 16 This indicated that the variance effects of peony episperm extracts depended on the concentration of these compounds, and the higher concentration could lead to the higher growth inhibition on liver cancer cell line HepG2.
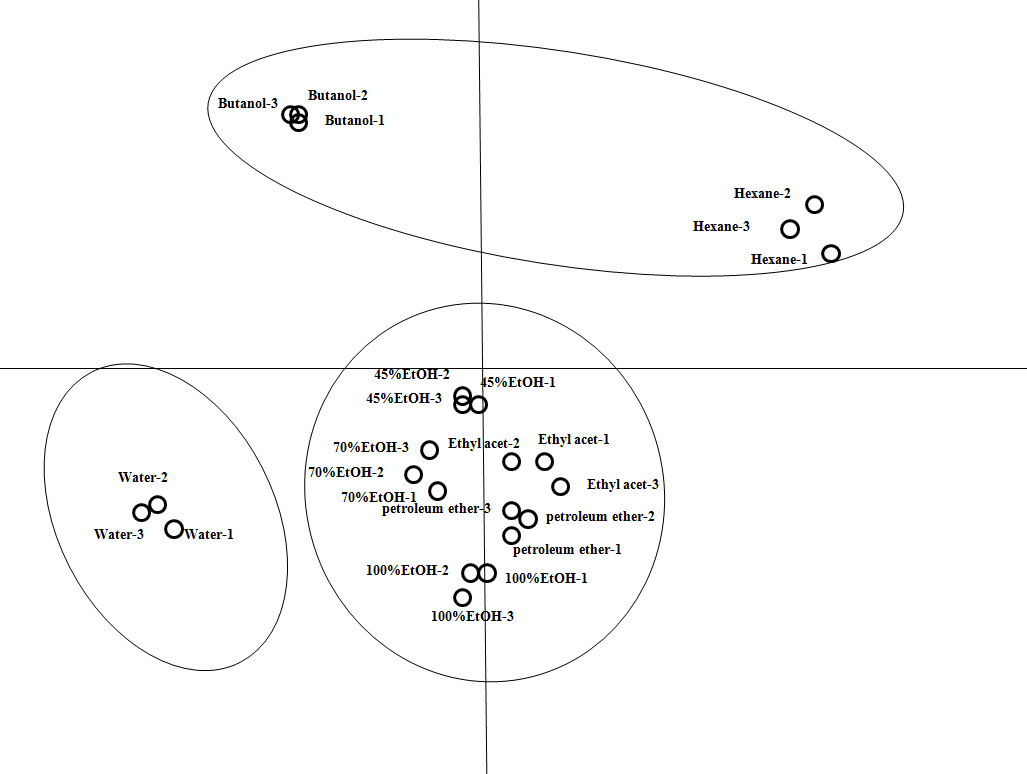
Principal component analysis scores plot for peony episperm crude extracts. Each point on the plot represents one HPLC dataset of an extract and points with the same name indicate replicates (n = 3).
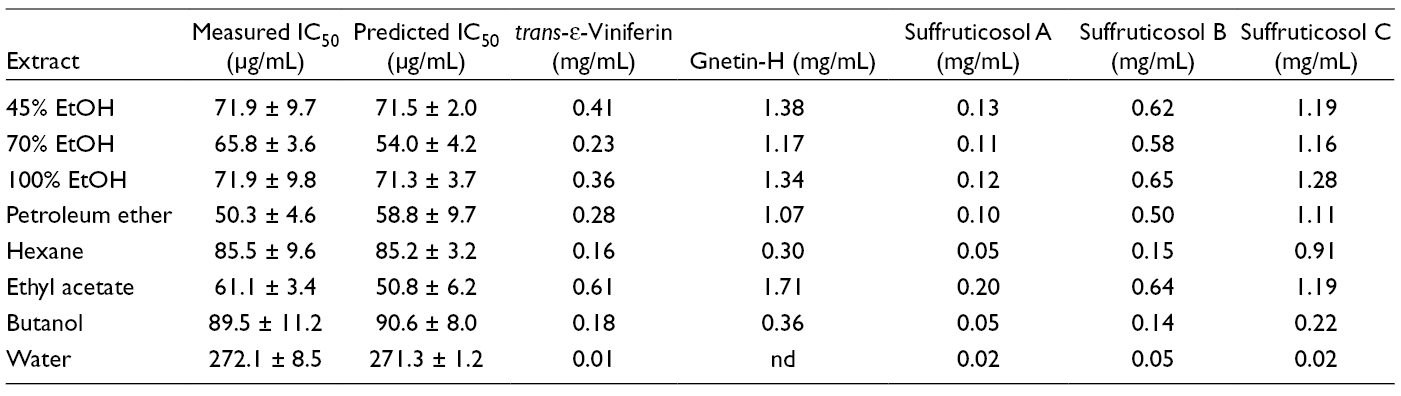
Partial least squares regression, the supervised classification and regression model, was further employed to provide a correlation between complex multivariate chemical datasets and the experimental IC50 of peony episperm extracts. The IC50 values of the crude extracts against liver cancer cells were imported as the y-matrix in the corresponding HPLC-PLS regression model. Three components were used for the model, with variance (R 2) values at 0.992 and cross-validated variance (Q 2) values at 0.925. The value of Q 2 above 0.5 was considered as the good prediction of the constructed model. 13 The overall predictions for each extract were summarized in Table 1. The correlation coefficient between predicted vs actual values was 0.996.
Experimental and Predicted IC50 Values of Peony Episperm Crude Extracts on HepG2 Liver Cancer Cells and Concentration of trans-ε-Viniferin, Gnetin-H, Suffruticosol A, Suffruticosol B, and Suffruticosol C in Each Extract.
Determination of Stilbene Compounds in Peony Episperm Extracts
The concentrations of trans-ε-viniferin, gnetin-H, suffruticosol A, suffruticosol B, and suffruticosol C in 8 extracts were determined by standard calibration curves and summarized in Table 1. Among them, gnetin-H and suffruticosol C presented relatively highest amounts in all of peony episperm extracts, while the concentration of suffuticosol A was lowest. Moreover, the water extract contained the lowest amount of 5 stilbenes tested, and the doses of 5 stilbenes were lower in hexane and butanol extracts than those of ethanol, petroleum ether, and ethyl acetate extracts of the peony episperm.
Validation of Peony Episperm Inhibitors of Human Liver Cancer Cells and Their Apoptosis Inducing Activity
To further validate the prediction above, the individual stilbene components were tested for inhibition on HepG2 cancer cells in vitro. All of 5 compounds presented the dose-dependent growth inhibitory effects against liver cancer cells. As shown in Table 2, the IC50 values for trans-ε-viniferin, gnetin-H, suffruticosol A, suffruticosol B, and suffruticosol C were found to be 64.71 ± 3.00 µg/mL (142.36 ± 6.60 µM), 74.60 ± 7.29 µg/mL (109.66 ± 10.72 µM), 208.66 ± 17.15 µg/mL (306.37 ± 24.21 µM), 98.19 ± 16.23 µg/mL (144.34 ± 23.86 µM), and 125.29 ± 13.12 µg/mL (184.18 ± 19.29 µM), respectively. These were comparable to those of the antineoplastic cisplatin (IC50: 132.33 ± 14.01 µM).

NR, no report.
Moreover, it was found that a proportion of HepG2 cells was stained by Annexin V-FLUOS (green) under 5 stilbene components treatment (Figure 3). Therefore, apoptosis was identified as one of the mechanisms for liver cancer cell death induced by trans-ε-viniferin, gnetin-H, suffruticosol A, suffruticosol B, and suffruticosol C.

Apoptosis induced by trans-ε-viniferin (1), gnetin-H (2), suffruticosol A (3), suffruticosol B (4), and suffruticosol C (5) on human liver cancer cells HepG2 using Annexin-V staining.
Discussion
In this work, HPLC-PCA secondary metabolite map was constructed to correlate the growth inhibitory activity of crude extracts on human liver cancer cell line HepG2. The stilbene compounds, trans-ε-viniferin, gnetin-H, suffruticosol A, suffruticosol B, and suffruticosol C, were thus first determined as the functional anticancer compounds of peony episperm. Nevertheless, a HPLC-PLS regression model was developed which could successfully make the prediction of inhibition by peony episperm crude extracts. The stilbene compounds were found to be apoptosis inducers on HepG2 cancer cells, and the IC50 values for trans-ε-viniferin, gnetin-H, suffruticosol A, suffruticosol B, and suffruticosol C were 64.71 ± 3.00, 74.60 ± 7.29, 208.66 ± 17.15, 98.19 ± 16.23, and 125.29 ± 13.12 µg/mL, respectively
The complex chemical profiles of HPLC analysis for crude plant extracts usually precluded the correlation between certain compounds with the growth inhibitory effects. In this work, PCA was employed to model HPLC data and to aid determination of discriminative contributors. As shown in Table 1, if the concentrations of trans-ε-viniferin, gnetin-H, suffruticosol A, suffruticosol B, and suffruticosol C were used to classify the extracts into the high, intermediate, and low dosage groups, then all extracts would be placed in the same group as that of the IC50 values model (Figure 2). The extracts with the most anticancer potency (IC50 values < 72 µg/mL) had the highest concentration of trans-ε-viniferin (0.23-0.61 mg/mL), gnetin-H (1.07-1.71 mg/mL), suffruticosol A (0.10-0.20 mg/mL), suffruticosol B (0.50-0.65 mg/mL), and suffruticosol C (1.16-1.28 mg/mL), respectively. The intermediate active extracts (85 < IC50 < 90 µg/mL) contained the medium amounts of trans-ε-viniferin (0.16-0.18 mg/mL), gnetin-H (0.30-0.36 mg/mL), suffruticosol A (0.05 mg/mL), suffruticosol B (0.14-0.15 mg/mL), and suffruticosol C (0.22-0.91 mg/mL). The least anticancer extract (IC50 >272 µg/mL) had the lowest concentration of trans-ε-viniferin (0.01 mg/mL), gnetin-H (not detected), suffruticosol A (0.02 mg/mL), suffruticosol B (0.05 mg/mL) and suffruticosol C (0.02 mg/mL). Therefore, increasing activities of extracts correlated with an increasing concentration of trans-ε-viniferin, gnetin-H, suffruticosol A, suffruticosol B, and suffruticosol C. Taken together, the stilbene compounds trans-ε-viniferin, gnetin-H, suffruticosol A, suffruticosol B, and suffruticosol C were predicted as the main inhibitors of the peony episperm against human liver cancer cells.
Moreover, if the predicted IC50 values of the corresponding PLS-HPLC model were used to classify the extracts again into the high, intermediate, and poor anticancer groups, then all extracts would be placed in the same group as that of the experimental data model (Figure 2). This clearly demonstrated the potential of using orthogonal chemical datasets to predict the pharmacological activity of herbal preparations based on the chemical composition and experimental data. It thus suggested that by using HPLC data in modeling IC50 values, it was possible to reasonably predict the cytotoxic effects of plant extracts which would normally require larger scale lab-based assays.
As shown in Table 2, the stilbene constituents of the peony episperm have been reported to inhibit the growth of a broad spectrum of cancer cells in vitro through numerous and competing apoptotic processes. 9,17-20 The trans-ε-viniferin inhibited a range of cancer cells, including A549, BT20, MCF7, MDA-MB-231, U2OS, HeLa, HT29, L1210, K562, HTC116, and HL-60, with IC50 values at 6.0 to 194.5 μM. 17 -20 The gnetin-H was reported to be cytotoxic to cancer cells HL-60, Hela, MCF-7, HT-29, L1210, K562, and HTC116 with IC50 values at 19.0 to 90.7 μM. 18 -20 The suffruticosol A inhibited cancer cells A549, BT20, MCF, MDA-MB-231, U2OS, HeLa and HT29 in vitro with IC50 values at 3.4 to 134.4 μM. 17,20 The suffruticosol B inhibited a range of cancer cells, including A549, BT20, MCF7, MDA-MB-231, U2OS, HeLa, HT29, and HL-60, in vitro with IC50 values at 4.8 to 137.4 μM. 17,20 The suffruticosol C showed the strongest cytotoxicity and inhibited cancer cells A549, BT20, MCF7, MDA-MB-231, U2OS, and HeLa in vitro with IC50 values at 2.4 to 18.4 μM. 17 The data of this work indicated similar growth inhibition effects on the human cancer cell line HepG2 and it had further related to the actual concentration of 5 anticancer compounds in crude extracts.
Moreover, this work confirmed the apoptosis inducing effects of stilbene constituents of the peony episperm, which were highly in agreement with the data reported in the literature. Suffruticosol B was found to suppress the expression of CYP1B1 gene and thus induced the apoptotic process in HL-60 cells. 19 Both trans-suffruticosol D and cis-suffruticosol D could induce A549 lung cancer cells into apoptosis at a concentration-dependent effect. Two apoptotic proteins, the inhibitor of apoptosis proteins family and X-linked inhibitor of apoptosis protein, were significantly downregulated by cis- and trans-suffruticosol D. 8 The cis- and trans-gnetin H could promote apoptosis in A549 and BT20 cancer cells by releasing mitochondria cytochrome c, activating caspase3/7, and inhibiting NF-κB activation. 9 In a recent research, 10 oligostilbenes, including resveratrol (E)-form, cis-ε-viniferin, trans-ε-viniferin, suffruticosol A, suffruticosol B, suffruticosol C, cis-suffruticosol D, trans-suffruticosol D, cis-gnetin H, and trans-gnetin H, were all reported to inhibit growth of human breast cancer cell MDA-MB-231 by inducing apoptosis. Increased levels of caspase3/7 were confirmed to be related to the apoptotic death of cancer cells in this work. 17 Taken together, the current results evidenced and confirmed the apoptosis-inducing abilities of trans-ε-viniferin, gnetin-H, suffruticosol A, suffruticosol B, and suffruticosol C on human liver cancer cells in vitro which expanded the fundamental understanding of the anticancer activity of peony episperm. However, the detailed anticancer mechanisms of peony stilbenes still are highly suggested to be further studied.
Conclusion
In this study, peony stilbene constituents trans-ε-viniferin, gnetin-H, suffruticosol A, suffruticosol B, and suffruticosol C were first identified as growth inhibitors and apoptosis inducers of human liver cancer cells with activities comparable to that of the antineoplastic cisplatin. The significance of this result was to systematically evident the phytochemical foundation of anticancer activities of the peony episperm. It expanded the fundamental understanding of the peony seeds, one of the most popular and promising traditional herbs, and supported its current medicinal uses in China. Moreover, PCA-mediated secondary metabolite mapping was employed to quickly identify the anticancer compounds from crude plant extract. This approach could be an important first step to qualify medicinal products required for industrialization and standardized production. The further work may include elucidation of detailed pharmacological mechanisms in molecular levels and structure modification of stilbenes, which is aiming to obtain potential anticancer agents or lead compounds from the peony episperm.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) declared no financial support for the research, authorship, and/or publication of this article.