
Abstract
Curcumin is a major component of turmeric with promising tumor-suppressive activity; however, its underlying molecular mechanisms need to be investigated further. We aimed to investigate the mechanisms of curcumin on the proliferation and migration of lung cancer cells. To confirm that, A549 lung cancer cells were cultured in vitro with or without curcumin; the effect of curcumin on cell proliferation was evaluated using the cell counting kit-8 assay, and its effect on cell migration was evaluated using a wound-healing assay. The results suggested that curcumin treatment inhibited the proliferation and migration of A549 cells, but these effects were alleviated when autophagy was inhibited by small interfering RNA that targets ATG5 or autophagy inhibitor 3-methyladenine. Similarly, inactivation of extracellular signal-regulated kinase 1/2 (ERK1/2) also attenuated the effects of curcumin on A549 cells. Collectively, our data suggested that curcumin induces autophagy via activating the ERK1/2 pathway and the autophagy is important for the inhibiting effect of curcumin in lung adenocarcinoma cells.
Lung cancer has gradually increased in incidence to become one of the most common human malignancies in both developed and developing countries. 1,2 Nonsmall cell lung cancer is the most common type of lung cancer, accounting for at least 80% of all lung cancer cases. Although there are currently many treatment options for lung cancer including surgery, radiotherapy, chemotherapy, and targeted therapy, half of all newly diagnosed cases are already at an advanced stage, when the effects of treatment are limited. Treatment in these cases may be limited to palliative care, resulting in a low 5-year survival rate.
Curcumin is a traditional Chinese medicine that has been used for dietary and medicinal purposes for thousands of years. Accumulating evidence suggested that curcumin can interact with multiple targets, and it has been used for centuries to treat numerous disease conditions, including different varieties of tumors. 3,4 It has also been found to be safe in human clinical trials; for example, oral curcumin improved the general health of colorectal cancer patients through enhanced p53 expression in tumor cells, leading to tumor cell apoptosis. 5 Garcea et al showed that oral administration of curcumin aided the pharmacologically effective levels in malignant colorectal tissues. 6 Many molecular mechanisms have been proposed to account for the tumor-suppressive effect of curcumin, including inhibition of extracellular signal-regulated kinase 1/2 (ERK1/2) activity, 7 inhibition of apoptosis signal-regulating kinase 1, 8 inhibition of the reactive oxygen species-dependent mitochondrial signaling pathway, 9 and upregulation of α1-antitrypsin. 10
Cell autophagy is a lysosome-dependent pathway of degradation that is specific to eukaryotic cells. Via autophagy, cells can degrade their own long-lived or misfolded proteins and damaged organelles, allowing the resulting biological macromolecules to be reused by the cells, and thus maintaining the stability of the cell environment. 11 The role of autophagy as a mechanism of cell survival or cell death is currently a topic of debate, though some evidence suggests that cancer cells may use autophagy as a survival mechanism to avoid death under certain conditions. 12
Curcumin has shown antitumor activity and has demonstrated a regulatory effect on autophagy. 13 Several studies have shown that curcumin can activate autophagy in lung cancer cells and inhibit proliferation of cancer cells. 3 However, the molecular mechanisms underlying the autophagy-induced effect in lung cancer cells need to be revealed further. To confirm the role of autophagy in the curcumin-induced effects on lung adenocarcinoma cells, we initially verified the effect of curcumin on the levels of autophagy in A549 cells. The autophagy-associated protein light chain 3 (LC3) is converted from LC3-I to LC3-II during autophagy and is translocated to the membrane of the autophagosome. We therefore transfected A549 cells with a green fluorescent protein (GFP)-LC3 plasmid, followed 24 hours later by treatment with 25 µM curcumin. After 24 hours, the distribution of LC3 was observed by fluorescence microscopy. To detect the LC3-II expression, A549 cells were treated with curcumin for 24 hours, and the expression of LC3-II in cell lysate was detected with western blot. The results suggested that curcumin treatment significantly induces LC3 puncta accumulation and enhances LC3-II expression (Figure 1a and b).
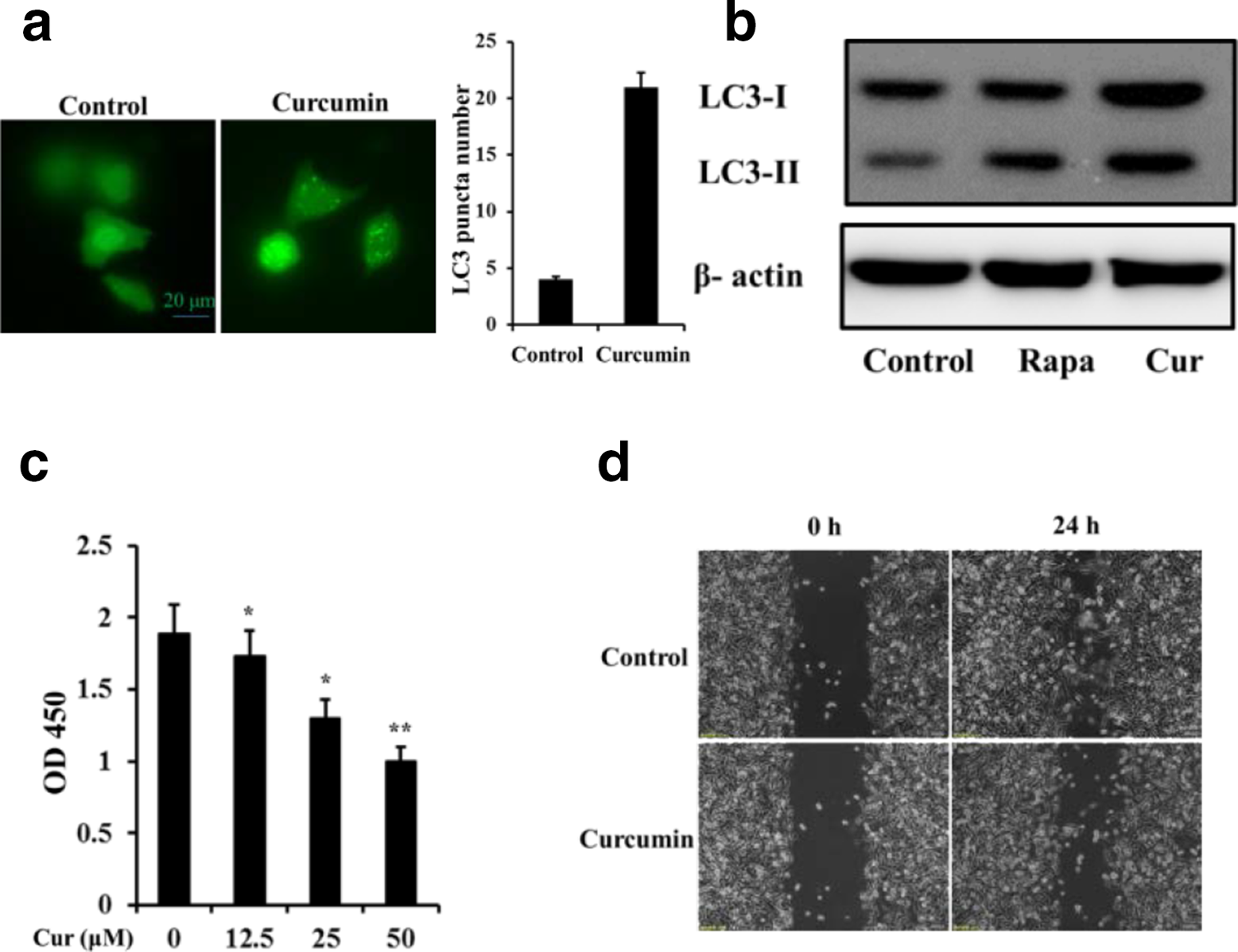
Curcumin induces autophagy and inhibits proliferation and migration in A549 lung adenocarcinoma cells. (a) Green fluorescent protein (GFP)-LC3 puncta formation. Representative images of GFP-LC3 puncta (left panel); quantitative analysis of numbers of dots in cells (right panel). (b) Western blot detects the protein expression of autophagy marker LC3. (c) Cell proliferation was evaluated using the cell counting kit-8 assay. (d) Cell migration was observed by phase contrast microscopy.*P < 0.05, **P < 0.01. Data are presented as mean±SD.
To verify the anticancer effects of curcumin in lung cancer cells, we first tested its impacts on lung cancer cell proliferation. A549 cells were incubated in growth medium with different concentrations of curcumin for 24 hours. The results are shown in Figure 1c. Curcumin treatment significantly inhibited the proliferation of A549 cells in a dose-dependent manner, as detected by cell counting kit-8 (CCK-8) assay. We next used a wound-healing assay to determine the impact of curcumin on cell migration. The results suggested that curcumin treatment markedly inhibited the migration ability of A549 cells (Figure 1d).
Next, we investigated the role of autophagy induction in the tumor-suppressive effect of curcumin by treating A549 cells with 3-methyladenine (3-MA), which can inhibit autophagy via its inhibitory effect on class III PI3K. We found that curcumin-induced cell autophagy was significantly inhibited by 3-MA (Figure 2a). In order to eliminate any impact from other possible effects of 3-MA, we treated the cells with a small interfering RNA (siRNA) targeting the autophagy-related gene ATG5. And we successfully knocked down the expression of ATG5 (Figure 2b). After that the siATG5 or control siRNA were transfected into A549 cells, 24 hours later; cells were treated with or without 25 μM curcumin for another 24 hours. As shown in Figure 2(c), knockdown of ATG5 also prevented curcumin-induced autophagy. Using a similar strategy, we found that the inhibitory effects of curcumin on cell proliferation and migration were alleviated in cells treated with 3-MA or ATG5 siRNA (Figure 2d-g).

Inhibition of autophagy reverses curcumin-induced inhibition of cell proliferation and migration. (a) Western blot was performed to detect the LC3-II expression of A549 cells under curcumin (25 µM) and 3-methyladenine (3-MA) (10 µM) treatment. (b) ATG5 expression was detected using western blot to evaluate the efficiency of ATG5 siRNA. (c) Accumulation of LC3-II upon autophagy activation was visualized by western blot. (d) and (e) Cell migration was observed by phase contrast microscopy. (f) and (g) Cell proliferation was evaluated by cell counting kit-8 assay. Results are expressed as means±SD for experiments performed in triplicate. **P < 0.01.
To evaluate whether curcumin can modulate the expression of ERK1/2, A549 cells were treated with 25 µM curcumin for different times, and protein levels of the phosphorylation status of ERK1/2 were detected by western blot. The results indicated that curcumin administration dramatically enhanced ERK1/2 phosphorylation and peeked at 24 hours, but total ERK1/2 expression levels were unchanged (Figure 3a). We hypothesized that inhibition of ERK1/2 activity might affect curcumin-induced autophagy. We therefore treated A549 cells, with or without 25 µM curcumin pretreatment, with the specific inhibitor U0126 for MEK, the upstream kinase of ERK1/2, to inhibit ERK1/2 phosphorylation. We found that treatment with U0126 for 24 hours notably inhibited the phosphorylation of ERK1/2 and suppressed the expression of LC3-II induced by curcumin (Figure 3b). In addition, pharmacological inhibition of the ERK1/2 signaling pathway also reversed the effects of curcumin on cell proliferation and migration (Figure 3b and c). These results suggested that ERK1/2 activation was involved in the curcumin-induced activation of autophagy and its consequent effects on cell proliferation and migration.

Extracellular signal-regulated kinase 1/2 (ERK1/2) signaling pathway activity was necessary for curcumin-induced autophagy in A549 cells. (a) A549 cells were treated with curcumin for indicated times and phosphorylation of ERK was detected by western blot. (b) ERK and LC3 expression levels were detected by western blot in A549 cells treated with curcumin with or without U0126. (c) Cell proliferation was evaluated by cell counting kit-8 assay. (d) Cell migration was observed by phase contrast microscopy. Results are expressed as means±SD for experiments performed in triplicate. **P < 0.01.
Autophagy is an evolutionarily conserved intracellular degradation system present in all eukaryotes, with important roles in many pathophysiological process, including development/differentiation, 14 immunity/inflammation, 15 metabolism, 16 obesity/diabetes, 17,18 and aging. 19 The role of autophagy in the regulation of cancer is complex. Autophagy has been shown to inhibit cancer development, such as in the study by Viry et al, who showed that autophagy improved the immune cell surveillance capacity, thereby inhibiting cancer development. 20 Similarly, other studies found that cell autophagy inhibited the expression of nuclear factor-κB or inflammatory factors to reduce the incidence of cancer. 21,22 In contrast, autophagy may play an important role in promoting the tumor development process. Cancer cell proliferation needs to overcome lack of nutrition and oxygen supply, as well as other unfavorable factors, and the cancer cells may use autophagy products to resynthesize their own substances, thereby promoting their proliferation. 23 Inhibition of cell autophagy significantly inhibited the formation of distant metastases by liver cancer cells, and melanoma cell cloning ability was also reduced. 24,25 The formation of tumor blood vessels has been associated with increased autophagy levels in some solid tumors. 26 Clinical treatment with chemotherapy drugs can often induce cell autophagy, which may act as a double-edged sword: Inhibition of autophagy can sometimes improve the effect of chemotherapy drugs, but chemotherapy-induced apoptosis may depend on cell autophagy.
Curcumin is a phenolic pigment derived from the roots or tubers of turmeric plants and is the main component of the traditional Chinese medicine, turmeric. Its potential for treating lung cancer has long been recognized, 27 and previous studies have shown that curcumin can increase autophagy in lung cancer cells. However, the role of curcumin-induced cell autophagy in its inhibition of proliferation and metastasis of lung cancer cells has not been confirmed. The effects of curcumin on protein localization and the expression of LC3-II in A549 cells in the current study supported previous reports of curcumin-induced cellular autophagy. 7 We further confirmed that curcumin could enhance the level of autophagy in A549 cells as well as significantly inhibit their proliferation and migration abilities. Blocking autophagy with an inhibitor or siRNA significantly reduced the inhibitory effects of curcumin on cell proliferation and migration.
ERK is a part the of mitogen-activated protein kinase superfamily and is well known for its ability to modulate many cellular functions, including proliferation and differentiation. 28 Several reports have found the complex roles of curcumin on ERK in cancer cells. Huang et al revealed that curcumin inhibits growth and induces apoptosis in human prostate cancer cells and was associated with decreased levels of phospho-ERK1/2. 29 However, curcumin induces apoptosis of human lung adenocarcinoma cells and was accompanied by sustained phosphorylation of ERK1/2. 9 Li et al revealed that curcumin activated the ERK1/2 signaling pathway in chondrocytes. 30 In the current study, we examined the essential role of the ERK pathway in lung cancer cell autophagy by treating A549 cells with the inhibitor U0126 and showed that inhibition of this pathway reduced the induction of autophagy by curcumin and attenuated the curcumin suppression effect on cell proliferation and migration. Altogether, our results indicate that ERK1/2 activation by curcumin induces autophagy and subsequently suppresses cell proliferation and migration. This provides a new evidence for a novel theoretical guidance whereby curcumin for clinical treatment of lung cancer.
Experimental
Reagents
The human lung adenocarcinoma A549 cell line was purchased from the Shanghai Cell Center of the Chinese Academy of Sciences (Shanghai, China). SiATG5 was designed and synthesized by Suzhou GenePharma Corporation (Suzhou, Jiangsu, China). LC3 antibody was purchased from Cell Signaling Technology (Danvers, MA, USA). β-Actin antibody and horseradish peroxidase-linked secondary antibody were from Beyotime Biotechnology (Shanghai, China). RPMI-1640 medium and fetal bovine serum were purchased from Gibco (USA). 3-Methyladenine and rapamycin were obtained from Sigma-Aldrich (St Louis, USA).
Cell Culture and CCK-8 Detection of Cell Growth Rate
A549 cells were cultured in RPMI-1640 medium containing 10% fetal bovine serum and routinely passaged at 37°C in a 5% CO2 incubator. The cells were seeded in 96-well plates and grown to 50% to 70% confluence, after a series of treatment, 10 µL of CCK-8 reagent (Beyotime, China) was added to each well, followed by incubation for 2 hours. The absorbance at 450 nm was then measured using a microplate reader.
Cell Migration Assay
Cell migration assays were performed using a cell culture insert (No.80209, ibidi, Munich, Germany). In brief, A549 cells were seeded at a density of 2 × 104 cells into a 3.5-cm Petri dish containing an insert and grown overnight. The cells were then washed with phosphate-buffered saline, the inserts were removed, and the cells were allowed to continue to grow for another 24 hours. Migrating cells were then examined and photographed under a light microscope.
Western Blot
Cells were lysed using RIPA lysis buffer (50 mM Tris pH 7.4, 150 mM NaCl, 2 mM ethlenediamine tetraacetic acid, 1% NP-40, 0.1% sodium dodecyl sulfate) at 4°C for 30 minutes. Total protein concentrations were quantified using a colorimetric detection assay (BCA Protein Assay, Pierce, Rockford, IL, USA). Equal amounts of protein were separated by 12.5% polyacrylamide gel electrophoresis and transferred to a nitrocellulose membrane. The membrane was blocked with 5% skimmed milk and incubated with primary antibodies against LC3, ERK1/2, p-ERK1/2, and β-actin overnight at 4°C, followed by washing 3 times with TBST for 5 minutes each. After that, the membrane was incubated with secondary antibody for 1 hour at room temperature and then washed 3 times. The membranes were then detected using the enhanced chemiluminescence system (Pierce).
Quantification of GFP-LC3 Puncta Formation
Cells were transfected with GFP-LC3 vector and treated with or without 25 µM curcumin for 24 hours, followed by fixing in 4% paraformaldehyde for 20 minutes. The accumulation of GFP-LC3 puncta was detected under a fluorescence microscope using the same fixed scan parameters and settings. Green fluorescent proteinGFP-LC3 puncta formation was quantified by counting at least 100 cells in each of 3 or more independent experiments.
Statistical Analysis
All experiments were conducted in triplicate to ensure that the results were reproducible. Data are presented as the mean±SD. A P-value < 0.05 was considered significant, and P < 0.01 was considered highly significant.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/orpublication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The research was financially supported by the Natural Science Foundation of Jiangsu province (BK20170251), the Nature Science Foundation of Jiangsu Higher Education Institutions of China (17KJB320022), Applied Basic Research Program of Xuzhou Jiangsu (KC18046), and Master’s Innovation Team Project of Xuzhou Central Hospital (XZS201640).