
Abstract
Background:
Testicular toxicity following chemotherapy is of increasing importance with the continuous improvement of survival rates. Gonadotropin-releasing hormone (GnRH) was suggested to protect testis against such toxicity; however, its suppressive quality and mechanism of action are still unclear. We examined whether and how pretreatment with GnRH antagonist protects against the testicular damage caused by chemotherapy.
Methods:
Mature male mice were injected subcutaneously eight times in 2-day intervals with either saline or GnRH antagonist (Cetrotide; 1 g/mg), followed by an intraperitoneal injection with either saline or cyclophosphamide (CTX;100 mg/kg BW) and sacrificed 2 weeks or 3 months later. Testicular weight, epididymis weight, epididymal sperm count and sperm motility were measured. Serum anti-Müllerian hormone (AMH) was measured by enzyme-linked immunosorbent assay. Immunohistochemistry (Ki-67), immunofluorescence (PCNA, CD34), terminal transferase-mediated deoxyuridine 5-triphosphate nick-end labeling (TUNEL) and computerized analysis were performed to examine testicular proliferation, apoptosis and vascularization. Quantitative real-time PCR was used to assess the amount of spermatogonial reserve (Id4 and Gfra1 mRNAs).
Results:
Pretreatment with GnRH antagonist transiently reduced testicular weight, epididymal weight, germinal proliferation and sperm count; it also abolished the permanent long-term effect of CTX on these parameters and prevented cyclophosphamide-induced testicular toxicity characterized by apoptosis and serum AMH increase and irreversible loss of spermatogonial reserve.
Conclusions:
Our findings imply that pretreatment with GnRH antagonist temporarily reduces spermatogenesis and may be used as pretreatment for reducing chemotherapeutic testicular toxicity.
Introduction
With the increasing success of anti-cancer treatments in prolonging the life expectancy of patients, came the realization that the induced permanent damage to self-renewing cell populations may also result in a variety of harmful side effects to fertility potential, ranging from temporary oligospermia to permanent azoospermia and sterility. 1 Gonadal injury following radiation or chemotherapy is of increasing importance with the continuous improvement of survival rates. Thus, attempts to prevent the negative effect of chemotherapy on fertility are of great importance. A variety of biochemical and biological approaches has been tested in experimental animal model systems to protect the testes against radiation or chemotherapy. A great research effort, which involves hormonal modulation, is conducted in an attempt to prevent or reverse the damage caused to germ cells by radiation or chemotherapy. 2
The hypothalamic decapeptide gonadotropin-releasing hormone (GnRH) binds to specific G protein-coupled receptors on pituitary gonadotrophs, leading to its activation; namely, phosphoinositide breakdown with generation of inositol trisphosphate and diacylglycerol as second messengers. Calcium release from intracellular stores is initiated, as well as activation of protein kinase C, which is important for gonadotropins synthesis and secretion. 3 The two pituitary gonadotropins, follicle-stimulating hormone (FSH) and luteinizing hormone (LH), induce spermatogenesis mainly through regulation of Sertoli cells and Leydig cells, respectively, at the seminiferous tubules. 4 GnRH analogues interact with the GnRH receptor, regulate GnRH activity and modify the release of FSH and LH. There are two groups of GnRH analogues, agonists and antagonists. Prolonged activation of GnRH receptors by GnRH agonists leads to cell desensitization and consequently to suppressed gonadotropin secretion. By contrast, GnRH antagonists compete with GnRH for receptors on gonadotroph cell membranes, inhibit GnRH-induced signal transduction and consequently gonadotropin secretion. 3
Presenting an immediate onset of action, GnRH antagonists may be more suitable for use just before chemotherapy treatment. 5 Chronic treatment with GnRH analogues reduces testicular function and spermatogenesis in mice. 6 It is not clear yet whether suppression of GnRH and gonadotropins by GnRH analogues protects testes against chemotherapeutic effect. Although several studies support the assumption that gonadotropin suppression protects mice, rats or monkeys testes against irradiation, chemotherapy, toxicants and heating damage thus enhances future fertility in face of the damage they cause,7,8 other reports suggest that hormonal suppression in mice does not protect spermatogenesis from damage.9–11 Moreover, only few studies in humans imply a possible protection of spermatogenesis in male cancer patients treated with GnRH analogue, 2 but the mechanism of action is still unknown. Glode et al. 12 showed that treatment of male mice with GnRH analogue produces prominent protection against histologically acute damage, 2 weeks after cyclophosphamide (CTX) administration. Their findings suggest that the reversible and temporary interruption of the pituitary–gonadal axis may ameliorate the toxic effect of systematic chemotherapy on the gonads. However, this study did not investigate long-term effect as no permanent long-term effect of CTX was observed in the experimental setting and the long-term effect of GnRH analogue could not be determined. Delic et al. 13 showed in rats that protection of spermatogenesis during chemotherapy might be achieved by gonadotropin suppression. Histomorphometric evaluation showed that mice that received GnRH antagonist and cetrorelix, together with cisplatin chemotherapy, displayed better recovery of spermatogenic epithelium and larger amount of spermatogonia compared to mice treated with chemotherapy-only. 14 Moreover, Udagawa et al. 15 showed that treatment with GnRH analogues improved spermatogenesis recovery following chemotherapy in mice, whereas Meistrich et al. 16 showed that the administration of GnRH agonists and antagonists has a protective effect against radiotherapy in rats.
CTX is a cytotoxic alkylating agent, widely used as a chemotherapeutic anticancer and immunosuppressive agent; yet it may cause long-term or permanent gonadal damage in young male patients. 5 The testicular cytotoxic effect of CTX targets Sertoli cells, Leydig cells and germ cells and reduces the population of germ cells lineage. Although several groups have tried in the past to find a method or mechanism that will protect the testicle from the peripheral damage of chemotherapy, the above important issue has not yet been proven and is of great value in helping cancer survivors. Our system in mice of the present study induced not only partial but also long-term damage induced by chemotherapy and aimed to evaluate whether pretreatment with GnRH antagonist can protect against the long-term permanent testicular damage caused by CTX chemotherapy and to elucidate the mechanism of action of such protection.
Materials and methods
Experimental design in mice
Mature ICR male mice (2 months old; Envigo, Jerusalem, Israel) were housed in the air-conditioned, light-controlled animal facilities of the Sackler Faculty of Medicine in Tel-Aviv University. Animal care and all experiments were in accordance with the institutional guidelines and were approved by the Institutional Animal Care and Use Committee, Sackler Faculty of Medicine, Tel-Aviv University, ID TAU-R 100106. Mice were weighed and randomly divided into four groups. Animals were injected eight subcutaneous injections of saline or GnRH antagonist every 2 days (1 mg/kg BW; Cetrotide, cetrorelix acetate; MERCK KGaA, Germany; Cet), and an intraperitoneal injection of saline or CTX (100 mg/kg BW; Endoxan; Baxter Oncology GmbH, Halle, Germany), thus forming four groups: Control (nine injections of saline), Cet (eight injections of Cetrotide and one of saline), CTX (eight injections of saline and one of CTX) and GnRH antagonist and CTX (eight injections of Cetrotide and one of CTX). The dose and administration method of CTX were according to Xie et al. 5 and Mohammadi et al., 17 indicating a mild, though significant, long-term testicular damage. The dose and administration method of Cetrotide were according to Pareek et al., 18 showing that multiple injections of Cet resulted in a complete disappearance of germ cells, except the spermatogonial cells. Treatment discontinuation resulted in full recovery of spermatogenesis after 3 months. Mice were sacrificed with Isoflurane (Pharmal Healthcare, India) after either 2 weeks (for assessment of short-term effects) or 3 months (for assessment of long-term effects); testes were excised, weighed (indicator of testicular function) and further processed. Epididymides were also excised and weighed (indicator of spermatogenesis). Cauda epididymides were punctured and sperm were allowed to swim into M2 medium (M-7167; Sigma Chemical, St. Louis, MO, USA) at 37°C in 35 mm Petri dishes. Makler counting chamber (Sefi Medical Instruments, Haifa, Israel) was used to assess sperm concentration (indicator of spermatogenesis) and motility.
Enzyme-linked immunosorbent assay for AMH
Samples of blood, drawn from the inferior vena cava of sacrificed mice, were centrifuged (6000 rpm, 10 min, 4°C) and sera were stored at −80°C. Measurements of AMH by a designated Enzyme-linked immunosorbent assay (ELISA) kit (Beckman Coulter, Chaska, MN, USA) were according to the manufacturer’s instructions. 19 We have previously shown that chemotherapeutic treatment induces increase of serum AMH. Testicular and serum AMH levels may serve as diagnostic indicator for the severity of testicular damage, as determined in both preclinical and clinical settings. 19
Immunohistochemistry, immunofluorescence and terminal transferase-mediated deoxyuridine 5-triphosphate nick-end labelling
Sections of paraffin-embedded testes were randomly selected and processed as previously described for immunohistochemistry (IHC; the observer blinded to treatment 20 ), using the following primary antibodies: rabbit anti-Ki-67 (1:300; Spring Bioscience, CA, USA), rabbit anti-proliferating cell nuclear antigen (PCNA; 1:100; Santa Cruz Biotechnology, Santa Cruz, CA, USA) and rat anti-cluster of differentiation (CD34; 1:100; Cedarlane, Ontario, Canada). We used Hoechst 33,280 (1 µg/ml; Sigma Chemical) for DNA staining, mixed with the following secondary antibodies: HRP-conjugated donkey anti-rabbit (1:200; Abcam, Cambridge, MA, USA), Alexa-488-conjugated donkey anti-rabbit (1:200; Abcam) and Alexa-555-conjugated donkey anti-rat (1:200; Abcam). DNA fragmentation was examined by terminal transferase-mediated deoxyuridine 5-triphosphate nick-end labelling (TUNEL) according to the manufacturer’s instructions (Dead End Fluorometric TUNEL system; Promega, Madison, WI, USA). Positive control sections were exposed for 10 min to DNase I (6 units/ml; Invitrogen, Carlsbad, CA, USA). Bright-field images were recorded by a digital-camera (Canon pc1089 CCD, Tokyo, Japan) connected to an Axiovert 200 M inverted microscope (Carl Zeiss MicroImaging; Oberkochen, Germany) equipped with an Apochromat 20× objective. Florescence images were photographed by an LSM-510 confocal laser-scanning microscope (Carl Zeiss MicroImaging) equipped with Plan-Neofluar 25× objective. Offset calibration of the photomultiplier was performed using sections stained with secondary antibodies only. Ki-67 staining of tonsil tissue served as a positive control for immunoperoxidase staining. Randomly selected images of 50 transverse testes sections of three mice from each experimental group and from each staining method were analyzed. The average number of Ki-67 positive cells, PCNA positive cells, TUNEL positive cells or CD34 blood vessels was automatically analyzed by Fiji software (National Institutes of Health, Bethesda, USA). The fully automated quantitative analysis does not require subjective qualitative support of human experts. The Fiji software is designed to be used in image analysis studies. We followed the developer’s instructions in the particle tracking section. We have validated this method in previous studies.21–23 The use of TUNEL and its range as apoptotic index is familiar to us from a number of previous studies. In our extensive experience, even with very testicular-toxic agents (e.g. busulfan and doxorubicin) almost only germ cells are ever positive to TUNEL. Because Sertoli cells are positive only one in millions, a larger sample size should be used with possible automation analysis of thousands of images/sections.
Because CD34 also stains large seminiferous tubules, we have added a restriction in Fiji software to count only particles that have perimeter smaller than 100 µm, corresponding to testicular blood vessels.
Quantitative real-time PCR
Mice testicular RNAs were isolated and quantified 24 ; first-strand cDNA was created by 35 cycles of reverse transcriptase (Catalog Number 4368814, Applied Biosystems, Foster City, CA, USA) with 0.4 µM gene-specific primers, using ready-mix mixture (Sigma Chemical). The amount of mRNA was assessed by SYBR Green Reagent (SYBR Green PCR Master Mix, ABI, Carlsbad, CA, USA) on an ABI Prism 7900 Sequence PCR machine. In each run, we used 20 ng of cDNA per reaction as an amplification template. The primers used were as follows: mouse inhibitor of differentiation four (Id4) forward 5′ AGGGTGACAGCATTCTCTGC 3′; mouse Id4 reverse 5′ CCGGTGGCTTGTTTCTCTTA 3′; mouse family receptor alpha-1 (Gfra1) forward 5′ GCGTGTGAAGCACTGAAGTC 3′; mouse Gfra1 reverse 5′ GGTTCAGTTCCGACCCAAC 3′. The house-keeping gene selected for the Quantitative real-time PCR (qPCR) calibration was hypoxanthine-guanine phosphoribosyl transferase (HPRT1) and the primers used were as follows: HPRT1 forward 5′ CTCATGGACTGATTATGGACAGGAC 3′; mouse HPRT1 reverse 5′ GCAGGTCAGCAAAGAACTTATAGCC 3′. Data were recorded and analyzed by the StepOne 2.1 software (Applied Biosystems, Thermo Fisher Scientific, Waltham, Massachusetts, USA). Undifferentiated spermatogonia cells are constitutively dividing and are prone to chemotherapy-induced apoptosis, like neoplastic cells. 25 It was previously shown that qPCR may serve as precise tool to measure mRNA of transcription factors that are expressed exclusively in undifferentiated spermatogonia, namely ID4 or GFRA1.23,24,26
Statistical analysis
Quantitative measurements are presented as mean ± standard error (SEM). Data were evaluated by independent, two-sample t-test for unequal sample sizes and unequal variances with significance of p < 0.05. A correlated one-way ANOVA statistical analysis showed similar results.
Results
We examined chemotherapy-induced testicular toxicity in four groups of mature mice: control, Cet, CTX and Cet + CTX. Mice were sacrificed 2 weeks or 3 months later for assessment of short- and long-term gonadal effects of chemotherapy, respectively. Firstly, we examined several general markers of testicular function and spermatogenesis, which are affected by chemotherapeutic insults. Our findings demonstrated that Cet by itself, transiently reduced testicular weight (Figure 1(a)), epididymal weight (Figure 1(b)) and sperm count (Figure 1(c)) 2 weeks after drug administration, but the values returned to normal at 3 months. More importantly, pretreatment with Cet significantly reduced the long-term adverse effects of chemotherapy, as all three parameters were significantly higher in the Cyclo + GnRH antagonist group than in the Cyclo-only group (Figure 1(a)–(c); p < 0.05). Sperm motility was not affected by either Cet or chemotherapy in our experimental setup (data not shown). In accordance with the damage caused by chemotherapy, increased level of serum AMH, an indicator for chemotherapy-induced testicular toxicity, 19 was also prevented by pretreatment with Cet (Figure 1(d)).
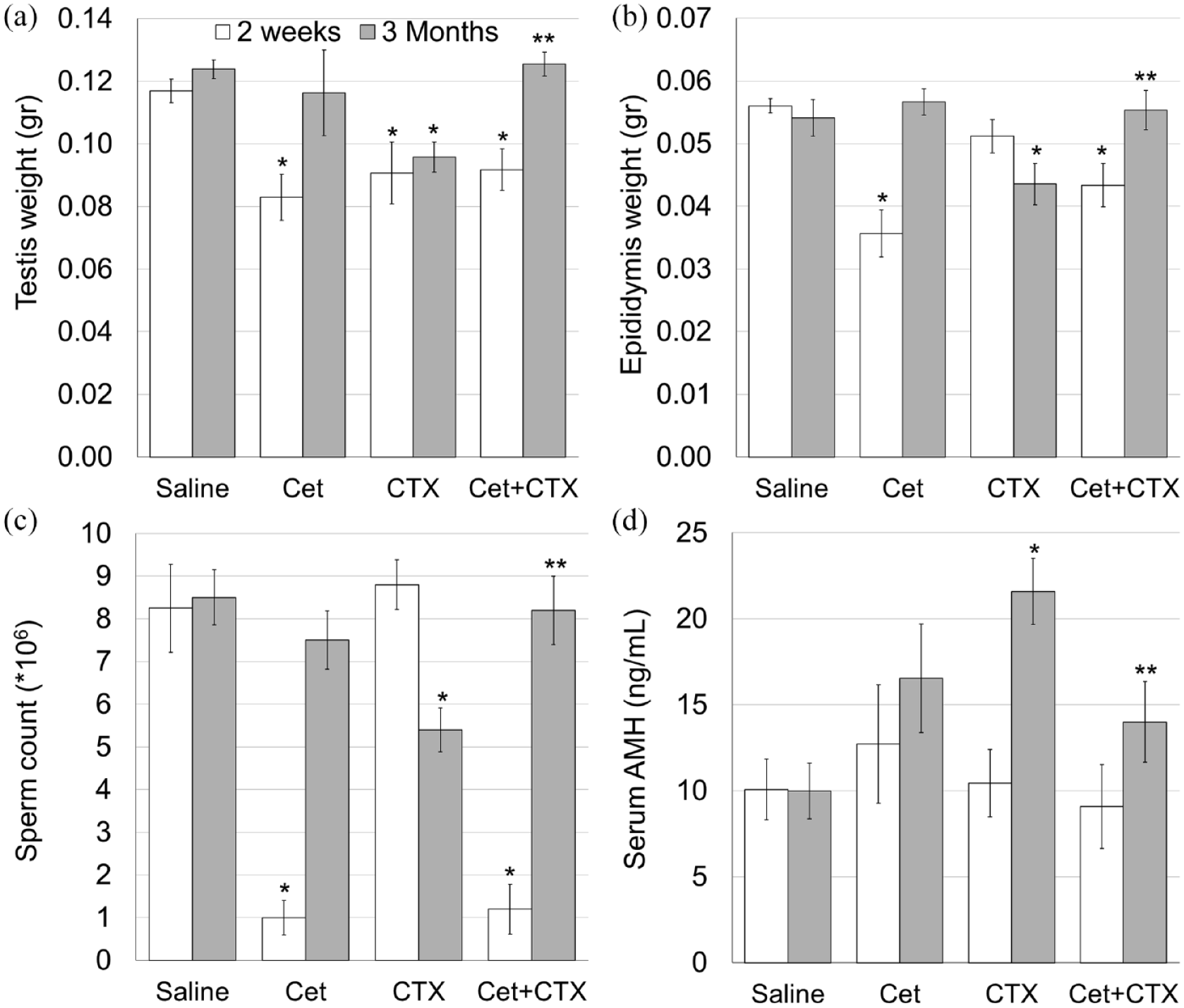
GnRH antagonist prevents the decrease in testicular and epididymal weights, sperm count and testicular toxicity caused by cyclophosphamide.
To examine the effect of treatment with Cet on testicular morphology, proliferation, apoptosis and vascularity we used immunohistochemistry, immunofluorescence and TUNEL assay on randomly selected images of transverse sections of mice testes from each experimental group, each staining and each time point (see additional details in Materials and methods). Our findings indicated a transient decrease of both proliferation markers (Ki67 and PCNA) in the testis, 2 weeks after administration of Cet and an irreversible decrease of these markers after CTX administration (Figures 2(a), 3(a), (b) and (b)–(h)), along with atrophy in part of the seminiferous tubules, 3 months posttreatment (Figure 2(f)). Pretreatment with Cet did not show long-term additive or synergic effect; it abolished the decreased proliferation caused by CTX (Figures 2(h), 3(a) and (b)). In addition, Cet inhibited the acute increase in apoptotic cells caused by CTX (Figures 2(a)–(h) and 3(c)). Testicular CD34 -positive vessels, was not affected by either Cet or CTX in our experimental setup (Figures 2(a)–(h) and 3(d)), implying that Cet exerts its testicular protection via a different mechanism.

Testicular characterization after exposure to cyclophosphamide with or without GnRH antagonist.
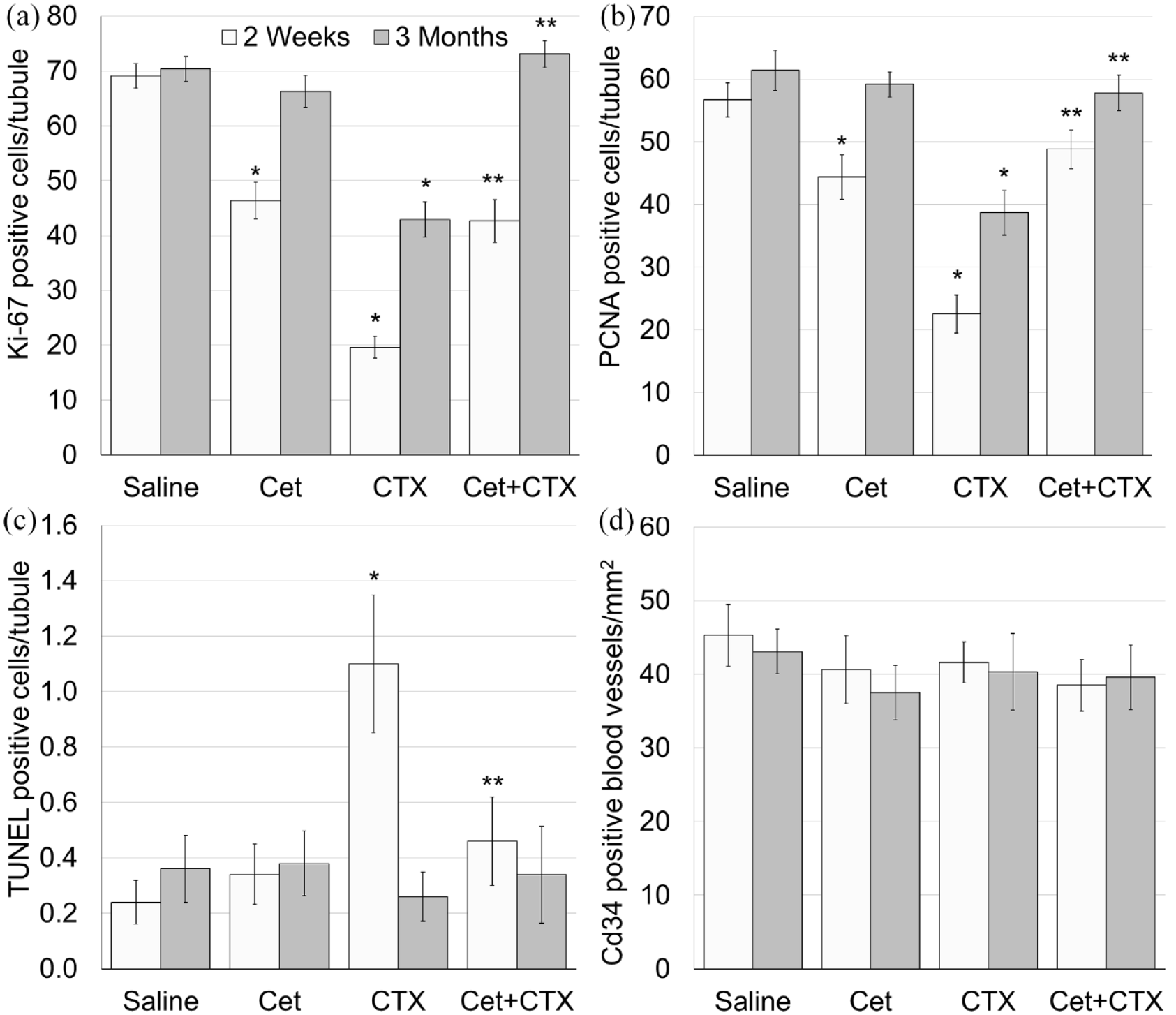
Testicular proliferation, apoptosis and blood vessels vascularity after exposure to cyclophosphamide with or without GnRH antagonist.
We used qPCR to measure mRNA of transcription factors that are expressed exclusively in undifferentiated spermatogonia, namely Id4 or Gfra1. 24 Pretreatment with Cet alone did not affect the amount of both indicators Id4 (Figure 4(a)) and Gfra1(Figure 4(b)), though it abrogated the acute and irreversible decline of both indicators caused by CTX.
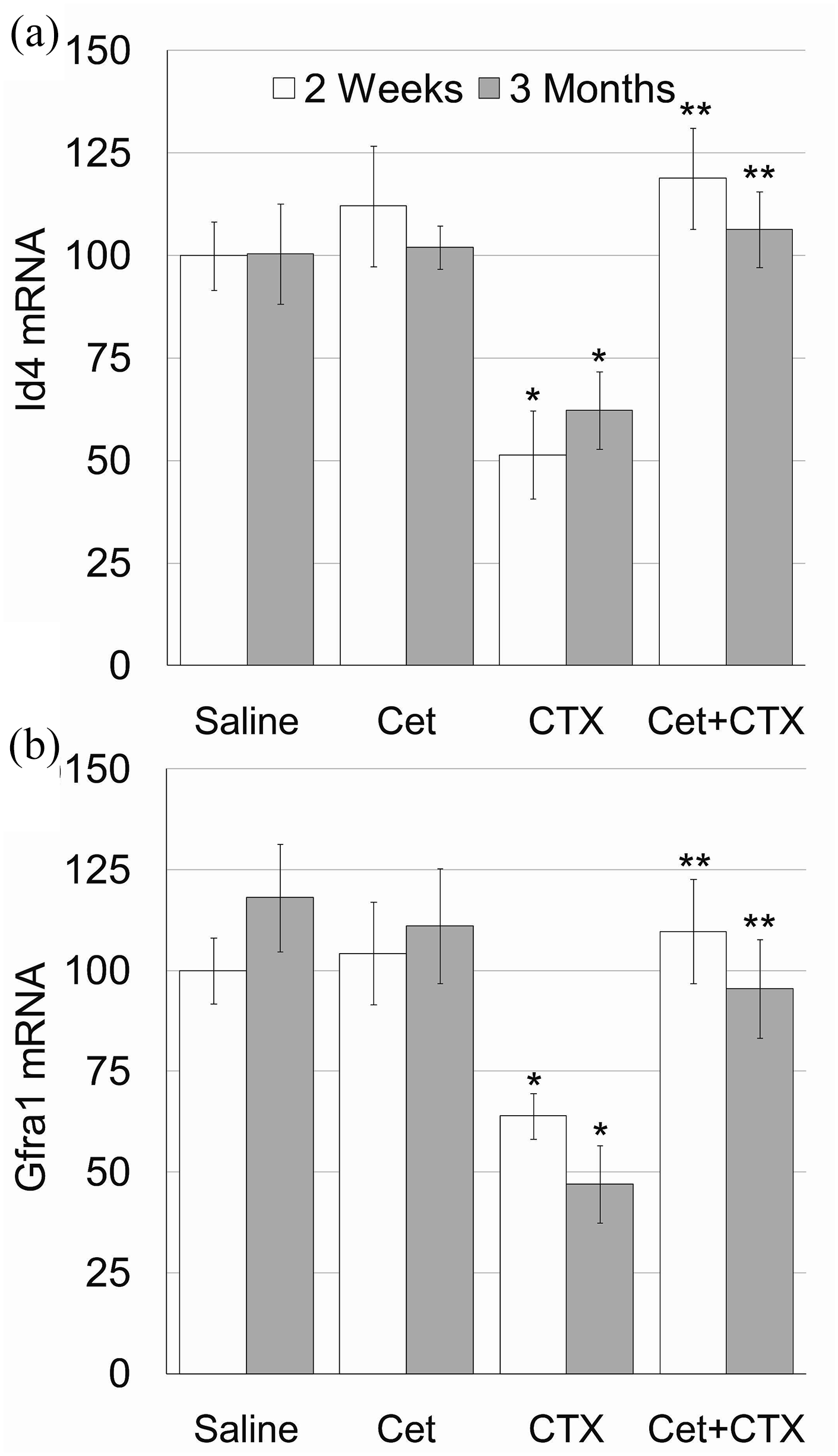
Testicular spermatogonial reserve after exposure to cyclophosphamide with or without GnRH antagonist.
Discussion
A series of Cet injections caused a complete disappearance of germ cells, except spermatogonial cells; but treatment discontinuation allowed a full spermatogenesis recovery. 18 Our findings indicate that testes of mice treated with Cet and CTX had a significantly higher mRNA expressed in spermatogonia compared to testes of mice treated with CTX alone. Spermatogonia constitute the testicular reserve and play an important part in testicular recovery and post-damage repopulation. Several studies also showed that mice and rats spermatogonia are affected by hormonal suppression.2,27–29 Our findings indicated that pretreatment with Cet transiently reduced testicular and epididymal weights, testicular germinal proliferation and sperm count. It also abolished the permanent long-term effects inflicted by CTX upon these parameters and prevented CTX-induced testicular toxicity, characterized by apoptosis, serum AMH increase and an irreversible loss of spermatogonial mRNA. It is possible that the small number of TUNEL-positive cells reflects an existing state of cells in the process of death. It is possible that over a longer period of time, more cells would have been observed as TUNEL-positive in our experimental setup; implying that vascular toxicity is not part of the mechanism of action. GnRH antagonists competitively block pituitary GnRH receptors and cause an immediate suppression of LH, FSH and gonadal hormones secretion 30 ; suggesting that pretreatment with GnRH antagonists suppresses gonadotropin secretion, thus reducing spermatogonial proliferation. Since non-dividing cells are less prone to toxic effects of chemotherapeutic agents, spermatogenic cells, at their non-proliferating period, are also not affected and repopulate the testis with surviving spermatogonia. Our results imply that Cet alone did not affect the amount of mRNA expressed exclusively undifferentiated spermatogonia cells, and it is possible that it protects these cells against the long-term effect of chemotherapy through a mechanism that does not directly include proliferative or apoptotic processes. The expression of GnRH and its receptors in mouse and human testes suggests that GnRH can be part of a paracrine regulation system.31–34 GnRH plays an important role in the control of mammalian reproduction. In addition to its well-documented classic hypophysiotropic action, GnRH might also play a role as a modulator of cell growth and metastasis in a number of human malignant tumours, including breast, ovary, endometrium and prostate cancers. In addition, GnRH receptors, expressed in many tumour types, provide suitable targets for GnRH analogues therapy. 35 It has been claimed that GnRH inhibition protects against anti-cancer treatments for also females. However, the mechanism of action is yet to be determined. 36 Both GnRH agonists and antagonists have been widely used as therapeutic agents in treating sex steroid-dependent tumours. In accordance with our findings, several studies in mice, rats or monkeys suggest that GnRH suppression may protect against testicular damage.7,8,13,37–44 showed that GnRH analogue did not protect against testicular toxicity caused by CTX. However, their experimental setting was different from ours: they used different protocol, including X6 of the total dose of CTX, we used and did not assess the effect of the same GnRH analogue alone and showed that the effect of CTX was absolute (complete azoospermia and sterility), whereas we showed that the effect of CTX was mild and partially reversible.
The protection exerted by gonadotropin hormone antagonist in long-term adult male testicular cancer survivors has not yet been evaluated. Only several studies imply a possible short-term protection of spermatogenesis in human cancer patients treated with GnRH analogue. 2 This might be attributed to small sample size, heterogeneity of both cancer types and anti-cancer treatments, as well as difficulty in assessing fertility in male patients who hold large physiological fluctuations in sperm counts. 2 Our findings suggest that pretreatment with GnRH analogues may be beneficial in ameliorating chemotherapy-induced testicular damage.
Our findings suggest that spermatogenesis arrest caused by GnRH antagonist (Cetrotide) protects against the damage caused by chemotherapy (cyclophosphamide) in mice. Further research in clinical settings is needed to evaluate testicular toxicity and protection in mature male cancer patients going through treatments such as radiotherapy and chemotherapy or in autoimmune treatments with anti-mitotic drugs, both known to have negative effect on fertility. The clinical implication in patients that received neo-adjuvant GnRH-antagonist protecting treatment prior to the anti-mitotic therapy may lead to improvement in patient care by reducing one of the major negative side effects of long-term decreased fertility or even total sterility. The CTX used in our study was representative of chemotherapeutic treatment, which causes not only partial but also irreversible damage as part of an experimental system for examining a possible protective effect. In addition, the protection protocol by GNRH inhibition was also in line with our previous studies in animal models and professional literature. We assume that the treatment and protection protocol is critical for examining any possible protective treatment in humans. The GnRH analogues protection mechanism needs to be further studied in other experimental models as well as in a clinical setting.