
Abstract
Background:
Intestinal fibrosis-associated stricture can lead to bowel complications and subsequent surgeries in patients with Crohn’s disease (CD), but there are no widely accepted biomarkers for intestinal fibrostenosis.
Objectives:
This study aims to investigate the value of interleukins (IL) in detecting CD-related intestinal fibrostenosis.
Design:
This is an observational study.
Methods:
Transcriptomic profiling was performed from paired CD surgical resections containing non-involved and fibrostenotic segments. Data were integrated with a public dataset and a Luminex-based serum assay to identify fibrosis-related interleukins, which were further validated at mRNA and protein levels. Correlations with clinical indicators and surgical outcome were analyzed. Lastly, an in vitro assay was used to evaluate the pro-fibrotic effect of the candidate interleukin.
Results:
Bulk RNA-sequencing and public dataset revealed increased expression of IL11 in the fibrostenotic intestinal segments of CD patients, which was further validated by real-time polymerase chain reaction and immunohistochemistry. The serum Luminex assay showed that serum IL11 is significantly increased in stricturing CD patients compared to that in non-stricturing CD patients. Clinically, serum IL11 was correlated with disease behavior (r = 0.343, p = 0.006), and increased IL11 expression was linked to a higher risk of subsequent surgery (log-rank p = 0.0055). Furthermore, single-cell RNA sequencing revealed that IL11 and its receptor IL11RA were mainly expressed by fibroblasts. In vitro, IL11 functionally promoted intestinal fibrosis.
Conclusion:
IL11, mainly derived from fibroblasts, is enriched in fibrostenotic tissue of CD and promotes intestinal fibrosis. IL11 may serve as a potential biomarker for CD fibrostenosis.
Intestinal tissue from both non-involved and fibrostenotic areas of CD patients was analyzed using bulk RNA sequencing. Luminex assay was used to test the serum levels of various interleukins in CD patients with different types of disease behavior. This study indicated that IL11 was the only interleukin that was elevated in both fibrostenotic intestinal tissue and serum of CD patients with type B2. Single-cell RNA sequencing data and subsequent immunofluorescence staining revealed that IL11 was mainly expressed by fibroblasts. In vitro experiments showed the pro-fibrotic function of IL11.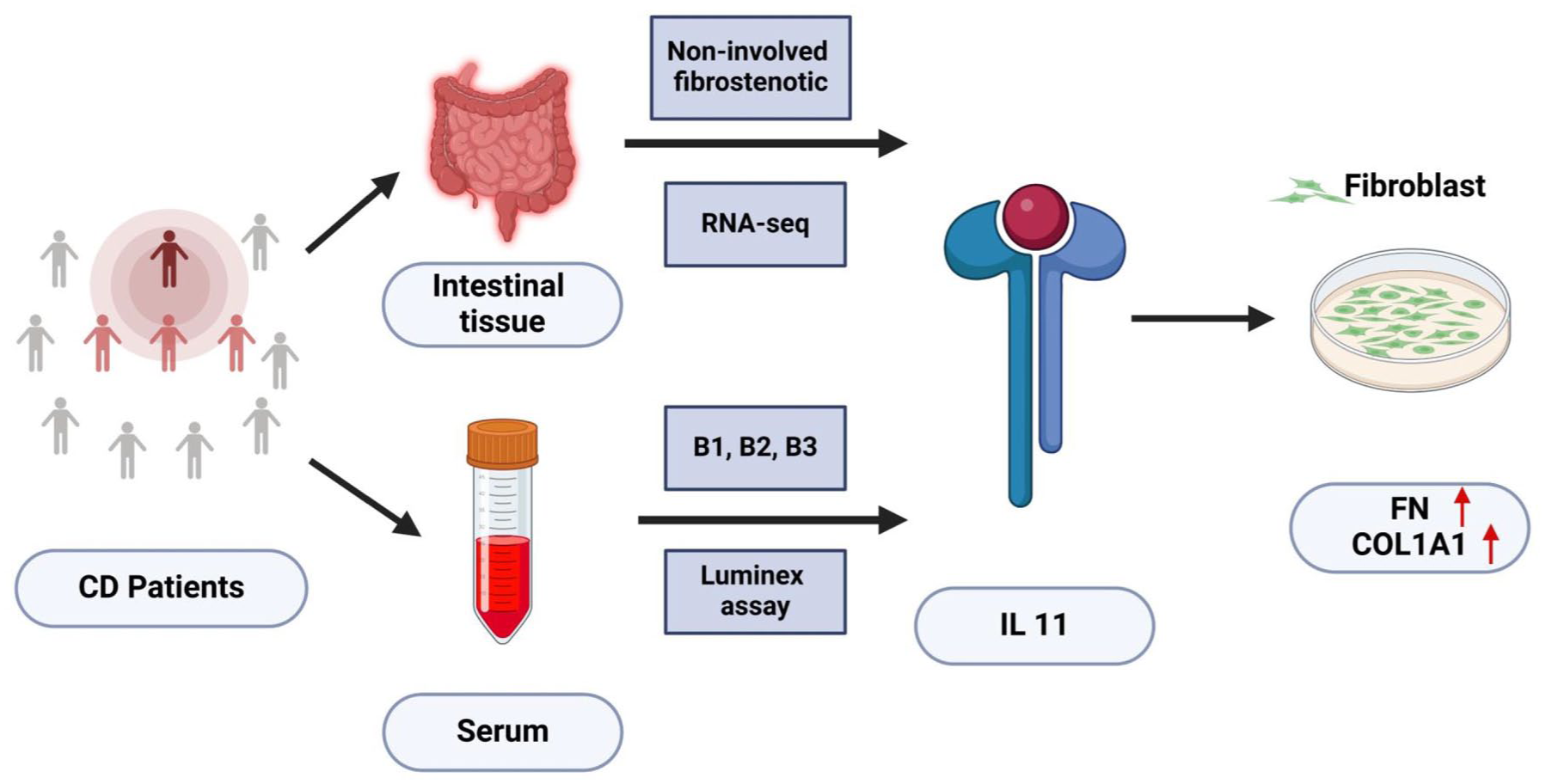
Introduction
Crohn’s disease (CD) can involve any part of the digestive tract with complex pathophysiological mechanisms.1,2 According to the Montreal classification, CD is categorized into three types: non-stricturing non-penetrating (type B1), stricturing (type B2), and penetrating (type B3). Despite the emergence of various biologics and small molecule agents, the prevalence of CD patients with strictures remains high. 3 Moreover, some patients with CD initially diagnosed with type B1 may progress into type B2 over time. 4 The stricturing CD can lead to intestinal obstruction, higher hospitalization rates, greater surgical risks, and increased healthcare costs, thus imposing a heavy burden on patients, families, and society. Therefore, early identification and timely intervention of these high-risk patients are crucial to reduce the occurrence of complications.
Some studies have already explored biomarkers for diagnosing and predicting stricturing CD, including genetic, histopathological, and serum biomarkers. 5 For instance, CD patients with variants of CARD15 (Caspase recruitment domain family, member 15) were more likely to have a stenosing pattern. 6 Low level of serum Cathelicidin suggested a higher risk of intestinal stricture of CD patients and showed better performance when combined with C-reactive protein (CRP) levels. 7 In addition, positive anti-Saccharomyces cerevisiae antibodies (ASCA) were more frequent in CD patients with type B2 or B3 than in those with type B1, while the presence of ASCA showed higher risk of disease progression. 8 However, due to limitations such as a lack of specificity, inconsistent conclusions, and incomplete understanding of the underlying mechanisms, these biomarkers have not been widely adopted in clinical practice. 5
CD is commonly accompanied by immune dysregulation, while cytokines play a key role in the immune response. 3 Interleukins (IL), a subset of cytokines, have been implicated to play roles in inflammation, and tumors.9,10 In recent years, a series of studies have successively suggested that IL also contributed to the occurrence and development of fibrosis.11–30 However, there has been no systematic study investigating the value of interleukins in detecting intestinal fibrostenosis in CD.
This study aims to explore the roles of various interleukins in the diagnosis of intestinal fibrostenosis in CD, with the purpose to identify potential biomarkers and therapeutic targets to improve early detection and clinical management.
Materials and methods
The study was conducted in the First Affiliated Hospital, Sun Yat-sen University, consecutively between 2016 and 2022.
Human subjects and specimens
A total of eight CD patients who underwent surgery in our cohort (The First Affiliated Hospital of Sun Yat-sen University (FAH-SYSU) cohort) were included in the study for RNA sequencing. During surgery, paired mucosa/submucosa specimens were collected from non-involved and fibrostenotic segments, and then immediately kept at −80℃ before transcriptomic profiling.
Besides, the serum of 63 CD patients was used to measure the levels of 20 interleukin molecules mentioned in the literature, and clinical correlation was analyzed among these patients. The inclusion criteria for these patients included (1) consecutive CD patients between 2016 and 2022; (2) the patients had not received treatment with immunosuppressants, biologics, or small molecule agents prior to their enrollment, while steroids were withdrawn for at least 3 months; (3) serum and intestinal specimens were obtained before the treatment; (4) the patients had more than 6 months of follow-up since the index evaluation. Patients were excluded if (1) they had any treatments such as steroids, immunosuppressants, biologics, or small molecule agents within 3 months prior to the enrollments; (2) they had a history of malignant tumors; (3) accompanied with infection (e.g., tuberculosis or cytomegalovirus infection); (4) pregnancy. All patient details have been de-identified. Baseline demographic and clinical characteristics were also collected, including gender, the time of diagnosis, Montreal classification, perianal disease, smoking history, white blood cell count, hemoglobin, platelet count, CRP, erythrocyte sedimentation rate (ESR), and serum albumin. Baseline simple endoscopic score for Crohn’s disease (SES-CD), a widely used tool for assessing disease activity in clinical trials,31,32 was also collected. A follow-up for at least 6 months was addressed to assess the surgical outcome. The reporting of this study conforms to the STROBE statement (Supplemental Table 4). 33
Murine model of intestinal fibrosis with dextran sodium sulfate
Male C57BL/6J mice (6–8 weeks old) were obtained from the Experimental Animal Center of Sun Yat-sen University and housed in a controlled environment with a temperature of 23 ± 3°C, relative humidity of 35% ± 5%, and a 12-h light/dark cycle. Following a 7-day acclimatization period, mice were randomly assigned to two groups (n = 6 per group). Intestinal fibrosis was induced in the experimental group by administering 2.5% dextran sulfate sodium (DSS; MP Biomedicals, Cat# 160110, Solon, Ohio, United States) dissolved in drinking water. The treatment protocol consisted of three cycles: each cycle included 7 days of DSS exposure followed by 14 days of recovery with normal drinking water. The control group received standard drinking water throughout the study. All animals were euthanized for tissue collection 2 weeks after completion of the final treatment cycle.
RNA isolation and sequencing
RNA isolation
The total RNA was extracted from the eight paired CD specimens using the method of Trizol. (1) Tissue preparation and homogenization: 50–100 mg of tissue were kept in liquid nitrogen and pulverized into a fine powder using a pre-cooled mortar and pestle. Transfer the powdered tissue directly into a tube containing 1 mL of TRIzol reagent and vortex immediately and vigorously. (2) Phase separation: add 0.2 mL of chloroform into 1 mL of TRIzol reagent and shake tubes vigorously for 15–30 s. Incubate the mixture at room temperature (RT) for 10 min, and centrifuge the samples at 12,000 rcf for 15 min at 4°C. (3) RNA precipitation: transfer the upper aqueous phase to a new tube, and add 0.5 mL of isopropyl alcohol. Mix the tube and incubate the samples at RT for 10 min to precipitate the RNA. Centrifuge at 12,000 relative centrifugal force (rcf) for 10 min at 4°C. (4) RNA wash: carefully discard the supernatant, and wash the RNA pellet by adding 1 mL of 75% ethanol. Vortex the samples and centrifuge at 7500 rcf for 5 min at 4°C. Wash RNA precipitate again with 1 mL ethanol and discard the supernatant. (5) Redissolving the RNA: air-dry the RNA pellet for 5–10 min at RT. Dissolve the RNA pellet in 30 µL of nuclease-free water by pipetting up and down gently. Incubate for 10 min on ice.
RNA sequencing
The extracted mRNA is enriched using mRNA Capture Beads. After purification with beads, the mRNA is fragmented using high temperature. The fragmented mRNA is then used as a template to synthesize the first strand of cDNA in a reverse transcription enzyme mixture system. While synthesizing the second strand of cDNA, end repair and A-tailing are completed. Next, adapters are ligated, and Hieff NGS® DNA Selection Beads (Yeasen, Shanghai, China) are used for purification to select target fragments. Polymerase chain reaction (PCR) library amplification is then performed, and finally, detection is carried out using the Illumina Novaseq X Plus by Gene Denovo Biotechnology Co., Ltd (Guangzhou, China).
Measurement of the interleukin molecules by Luminex
Serum samples were kept at −80℃ prior to measurement. The concentration of IL1beta/IL1F2, IL2, IL4, IL5, IL6, IL7, IL10, IL11, IL12/IL23 p40, IL13, IL15, IL17/IL17A, IL18/IL1F4, IL19, IL21, IL27, IL31, IL33, IL34, and IL36/IL1F8 in serum was quantified using the Luminex system following the manufacturer’s instructions (LabEx Co., Shanghai, China).
Immunohistochemistry
Immunohistochemistry was performed on formalin-fixed, paraffin-embedded full-thickness terminal ileum tissue obtained from six control (NL) individuals and six CD patients, including samples from both relatively normal and fibrostenotic areas. Tissue sections (4 µm thick) were deparaffinized in xylene and rehydrated through graded alcohols. Antigen retrieval was conducted by microwaving the sections in citrate buffer (pH 6.0) for 20 min at boiling temperature. To mitigate non-specific or high background staining due to endogenous peroxidase activity, the sections were treated with hydrogen peroxide (H2O2) for 15 min. Afterward, blocking was performed at RT for 1 h using a phosphate-buffered saline solution containing 10% normal goat serum and 1% bovine serum albumin. A rabbit monoclonal antibody against human IL11 (PA5-36544; Invitrogen, dilution 1:200, Rockford, Illinois, United States) was applied, and the sections were incubated overnight at 4°C. Detection was performed using a biotin-free horseradish peroxidase-polymer system, with 3,3′-diaminobenzidine (DAB) serving as the chromogen (DAB Substrate Kit, Vector, Newark, California, United States). Counterstaining was executed with hematoxylin to provide nuclear detail, followed by a sequential dehydration process and the application of a mounting medium for slide preservation.
Real-time PCR
Total RNA was extracted using TRIzol reagent (15596026; Thermo Scientific, Carlsbad, California, United States). A constant quantity of RNA (1 µg per sample) was reverse transcribed into complementary DNA (cDNA) utilizing the Evo M-MLV Reverse Transcription Kit II (AG11711; Accurate Biology). Subsequent quantitative PCR was conducted using SYBR Green Pro Taq HS (AG11701; Accurate Biology). Gene expression levels were quantified using the delta-delta Ct (ΔΔCt) method. The primer sequences used were shown in Supplemental Table 1.
Primary intestinal fibroblast culture
Intestinal mucosa and submucosa were obtained from the surgical specimens of CD patients, as previously reported. 34 These specimens were minced into pieces and then planted into culture plates. The tissues were cultured for 3 weeks with high-glucose DMEM medium containing 10% fetal bovine serum, 1% penicillin–streptomycin–amphotericin B antibiotics, 2.5% HEPES, and 2% L-Glutamine, to obtain primary human intestinal fibroblasts. Cells at passage 4–8 were used for subsequent experiments.
Single-cell RNA-seq data processing
Standard pipelines of Cell Ranger (10× Genomics) were used to do sequence processing, mapped to the reference genome (human, GRCh38; mouse, mm10) using cellranger v3.0.1.
Integration and clustering were performed using Seurat v5.0.1. Cells with mitochondrial gene percent ⩾25%, hemoglobin gene percent ⩾3%, fewer than 200 genes, or more than 6000 genes were filtered out. Doublets were identified and removed using DoubletFinder v2.0.4 based on the proportion of artificial k nearest neighbors. Through these quality control steps, low-quality cells or ambient RNA were removed. After filtering, gene counts were normalized using LogNormalize. R package Harmony v1.2.0 was used to remove batch effects. The integrated expression matrix was scaled and subjected to principal component analysis for dimensionality reduction. FindNeighbors and FindClusters functions were used for clustering, and RunUMAP was used for cluster visualization.
Bioinformatic and statistical analysis
For bioinformatic analysis, the DESeq2 package was used in bulk RNA-seq to identify differentially expressed genes (DEGs). 35 The cutoff criterion of a false discovery rate is <0.05 and fold change >1.5.
For analysis of clinical data, the baseline demographic and clinical characteristics were expressed in medians and interquartile range for continuous variables and frequency distributions for categorical variables. The comparison of serum interleukins between different disease behavior groups was based on Wilcoxon’s test with Benjamini–Hochberg adjustment due to the limited sample and the non-normal distribution of the data. The correlation between interleukin expression and clinical indicators was analyzed by Spearman’s correlation, while its association with surgical outcome was analyzed by Kaplan–Meier analysis with log-rank test.
All statistical analyses were performed using R v4.1.0. Statistical significance was defined as a two-sided p < 0.05.
The methods of immunofluorescence, western blot, RNA quality assessment, filtering of clean reads, and alignment with reference genome were detailed in the Supplemental Materials.
Results
IL11 was highly expressed in fibrotic intestinal tissue of CD patients
We firstly evaluated the expression of various interleukins in the intestinal tissue of CD patients using transcriptomic profiling. Baseline demographic and clinical characteristics of the eight CD patients who performed surgical resections in our FAH-SYSU cohort were detailed in the Supplemental Table 2. Masson staining showed that all fibrostenotic specimens of CD patients were severely fibrotic (Supplemental Figure 1). It should be noted that seven specimens from each of the two groups, which passed the quality control, were included in the final analysis. The global DEG list can be found in the Supplemental Materials.
The volcano plot (Figure 1(a)) showed that IL11, IL1A, IL1B, IL6, and IL33 were up-regulated, while IL32 was down-regulated in CD fibrostenotic tissue when compared with paired CD non-involved segments. Interestingly, IL11 was one of the most dominant increased interleukins in our FAH-SYSU cohort (Figure 1(b)). Gene set enrichment analysis also indicated that interleukin signaling was among the significantly enriched pathways, and IL11 emerged as one of the core enrichment genes within this pathway (Supplemental Figure 2(A)). We further collected more surgical samples (N = 15) using paired mucosa/submucosa specimens, obtained from both non-involved and fibrostenotic segments of the same CD patient, and performed real-time PCR (RT-PCR) analysis. The result revealed that IL11 expression was significantly higher in the fibrotic tissue group compared to the non-involved tissue group (Supplemental Figure 2(B)). To validate our findings, we explored the public transcriptome dataset GSE192786, which investigated the fibrotic signatures in CD patients. Bioinformatic analysis showed that IL11 and IL24 were up-regulated in the group of fibrotic biopsies when compared with non-fibrotic biopsies (Figure 1(c)). A Venn diagram indicated that IL11 is the only interleukin correlated with CD fibrostenosis (Figure 1(d)). Interestingly, correlation analysis showed that IL11 was also positively correlated with fibrosis-associated genes, such as COL1A1, PDGF, MMP1, and MMP7 (Figure 1(e)–1(h)).

IL11 was highly expressed in fibrotic intestinal tissue of CD patients. (a) Volcano plot shows up-regulated (red dots) and down-regulated (blue dots) IL family genes in mucosa/submucosa of fibrostenotic tissue compared with non-involved tissue in the FAH-SYSU cohort (n = 7). (b) Box plot shows the expression level of IL11. The y-axis represents the FPKM level of IL11 gene expression. Data are presented as median + IQ. Statistical differences were determined by the Mann–Whitney U test. (c) Volcano plot shows up-regulated (red dots) and down-regulated (blue dots) IL family genes in fibrotic tissue compared with non-fibrotic tissue in the GSE192786 dataset. (d) Venn diagram shows the interleukins that are commonly up-regulated at the fibrostenotic sites in both the FAH-SYSU cohort and the GSE192786 dataset. (e) Spearman correlation analysis shows the correlation between IL11 and COL1A1. (f) Spearman correlation analysis shows the correlation between IL11 and PDGFB. (g) Spearman correlation analysis shows the correlation between IL11 and MMP1. (h) Spearman correlation analysis shows the correlation between IL11 and MMP7.
Furthermore, we tested the RNA and protein levels of IL11 in additional subjects as an internal validation. Full-thickness terminal ileum tissues were obtained from six control individuals (NL) and six CD patients, which included both relatively uninvolved and fibrostenotic areas. RT-PCR showed IL11 has significantly higher expression in the fibrostenotic intestinal tissue of CD patients (CD-S) group than that in the relatively uninvolved intestinal tissue of CD patients (CD-N) and control subjects (NL) group (Figure 2(a)), which confirmed the result of the transcriptomic profiling in our FAH-SYSU cohort and GSE192786 dataset (Figure 1). Besides, we found that the IL11-positive cells were significantly increased in the CD-S group when compared with those in the CD-N group and NL group (Figure 2(b) and (c)). We also investigated the expression of IL11 in the murine model of intestinal fibrosis (chronic DSS model). After three cycles of DSS treatment, the chronic DSS group showed significantly higher IL11 expression than the control group, both at the RNA and protein levels (Figure 2(d)–(f)).

Both RNA and protein levels of IL11 were increased in stenotic tissue of CD patients. (a) RT-PCR shows IL11 expression in intestinal wall of NL, CD-N, and CD-S groups (N = 6). Data are presented as median + IQ. Statistical differences were determined by the Kruskal–Wallis test. Gene expression values were calculated using the 2− ΔΔCT method and normalized to β-actin. (b, c) IHC showed the number of IL11-positive cells in NL, CD-N, and CD-S groups (N = 6). Data are presented as median + IQ. Statistical differences were determined by the Kruskal–Wallis test. (d) RT-PCR shows IL11 expression in intestinal wall of the control and chronic DSS group. Data are presented as median + IQ. Statistical differences were determined by the Mann–Whitney U test. Gene expression values were calculated using the 2− ΔΔCT method and normalized to β-actin. (e, f) IHC shows the number of IL11-positive cells in the control and chronic DSS group. Data are presented as median + IQ. Statistical differences were determined by the Mann–Whitney U test.
CD patients with type B2 or type B3 have higher serum levels of IL11
Next, we investigated the serum levels of 20 interleukins (IL1beta, IL2, IL4, IL5, IL6, IL7, IL10, IL11, IL12, IL13, IL15, IL17, IL18, IL19, IL21, IL27, IL31, IL33, IL34, and IL36), which were related to fibrosis based on the literature.11–30 Serum samples were collected from 63 CD patients, and the baseline demographic and clinical characteristics of these patients were detailed in Table 1. Among them, 27 patients were classified as type B1, 18 as type B2, and 18 as type B3.
Baseline demographic and clinical characteristics of the CD patients who underwent serum interleukin level tests (n = 63).
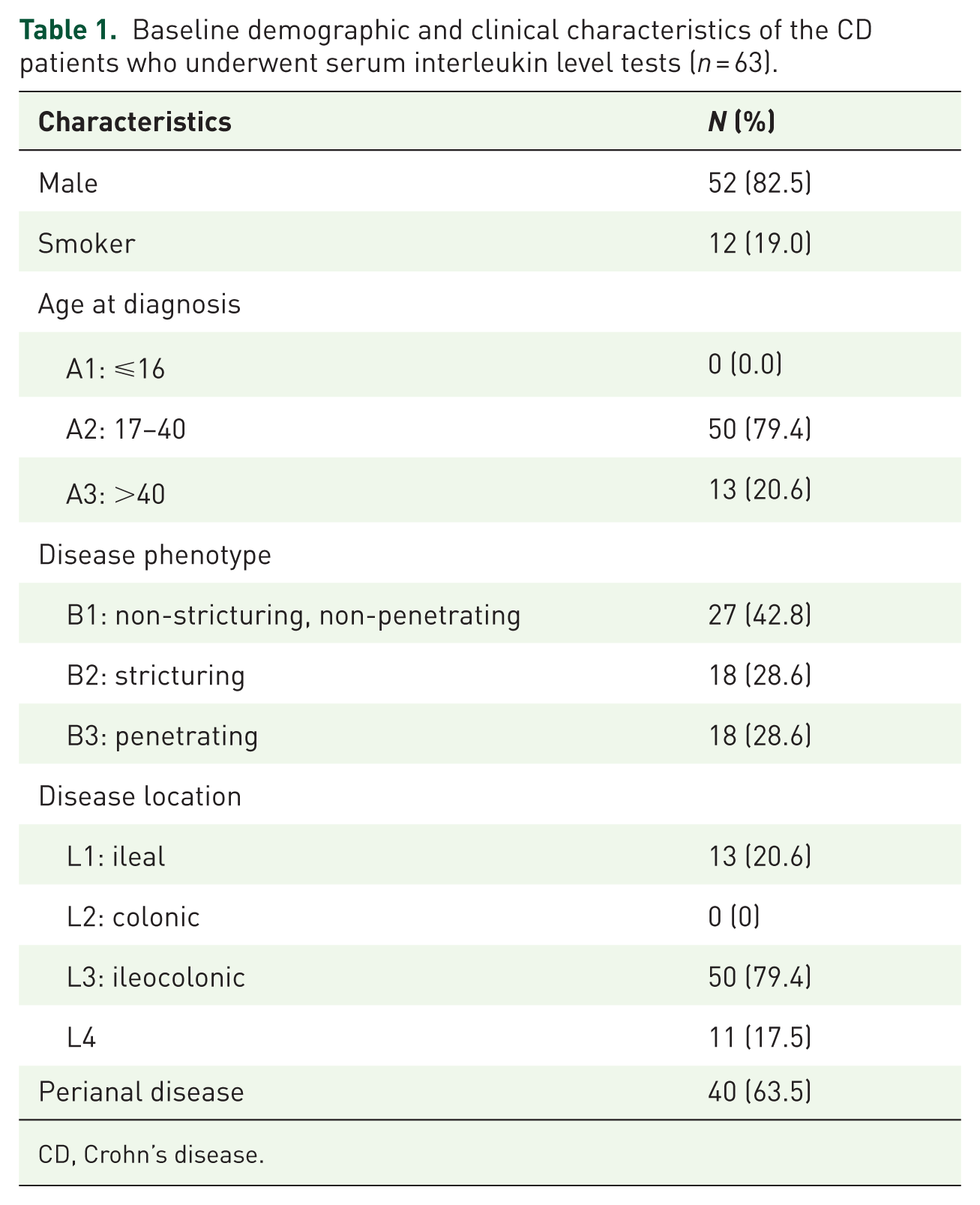
CD, Crohn’s disease.
Luminex assay was used to measure the serum concentrations of the interleukins. The result showed that IL11 was significantly elevated in CD patients with type B2 (p < 0.05) or type B3 (p < 0.05) than that in CD patients with type B1 (Figure 3(a)). As for IL17 and IL27, the serum levels were significantly higher in the type B3 group, but not in the type B2 group, when compared to the type B1 group (Figure 3(b) and (c)). However, there was no statistically significant difference in the comparison of the other interleukin molecules (IL1beta, IL2, IL4, IL5, IL6, IL7, IL10, IL12, IL13, IL15, IL18, IL19, IL21, IL31, IL33, IL34, and IL36) between any of the groups (Supplemental Figure 3). Thus, the serum assay suggested that IL11 might be a potential marker to distinguish the CD patients with fibrostenosis.

IL11 was correlated with clinical parameters, disease progression, and anti-TNF treatment response. (a) Luminex assay shows the serum level of IL11 in CD patients from type B1 (n = 27), type B2 (n = 18), and type B3 (n = 18) groups. Wilcoxon’s test with BH adjustment. (b) Luminex assay shows the serum level of IL17 in CD patients from type B1 (n = 27), type B2 (n = 18), and type B3 (n = 18) groups. Wilcoxon’s test with BH adjustment. (c) Luminex assay shows the serum level of IL27 in CD patients from type B1 (n = 27), type B2 (n = 18), and type B3 (n = 18) groups. Wilcoxon’s test with BH adjustment. (d) Dot plot heatmap shows the relationship between serum IL11 level and clinical parameters. The size of the dots represents the p-value, and the blue-red gradient indicates the correlation coefficient (r-value). (e) Spearman correlation analysis shows the correlation between IL11 and disease behavior. (f) Spearman correlation analysis shows the correlation between IL11 and CRP. (g) Spearman correlation analysis shows the correlation between IL11 and ESR. (h) Spearman correlation analysis shows the correlation between IL11 and SES-CD. (i) Kaplan–Meier curve shows the probability of subsequent surgery in the IL11high and IL11low groups in the CD patients.
IL11 was correlated with clinical parameters, disease progression, and anti-TNF treatment response
We further evaluated the associations between serum IL11 and clinical characteristics, laboratory examinations, endoscopic activity index, and surgical outcome in the 63 CD patients. Correlation analysis showed that serum IL11 was positively correlated with progressive disease behavior (type B2 or type B3; r = 0.343, p < 0.05), and CRP (r = 0.249, p < 0.05; Figure 3(d)–(f), Supplemental Table 3). IL11 was also positively correlated, but not statistically significant, with ESR (r = 0.179, p = 0.162) and SES-CD (r = 0.187, p = 0.176; Figure 3(g) and (h), Supplemental Table 3). In a subgroup with surgical outcome (n = 19), a median level of serum IL11 (42.02 pg/mL) divided patients into two groups (11 patients in IL11low group and 8 patients in IL11high group). Survival analysis showed that IL11high group encountered significantly earlier surgical outcome than that in the IL11low group (log-rank p = 0.0055; Figure 3(i)).
Next, we wonder whether the expression of IL11 in intestinal mucosa from CD patients was associated with anti-TNF treatment response. Using the public dataset GSE16879, we noticed that mRNA expression of IL11, at 4–6 weeks after first infliximab treatment, was significantly increased in the intestinal mucosa from patients with non-response to infliximab than those with good response (p < 0.05, Supplemental Figure 4(B)). Before the infliximab treatment, the mRNA expression of IL11 in the intestinal mucosa from patients with non-response to infliximab was also increased than those in the controls, but not statistically significant (p = 0.0733, Supplemental Figure 4(A)). Using another CD dataset GSE134881, we observed a similar result (Supplemental Figure 4(C)), indicating that intestinal mucosal IL11 expression may predict anti-TNF treatment response and guide clinical decision making.
IL11 and its receptor IL11RA were primarily expressed by fibroblasts
To determine the specific cell type expressing IL11, human single-cell RNA sequencing data generated previously (HRA006083) were used 36 . After quality control, we retained a total of 91,316 high-quality cells. We identified 11 main cell types through unsupervised clustering and classical marker gene annotation (Supplemental Figure 5(A) and (B)). Then we analyze the expression of IL11 in human single-cell RNA data. We found that IL11 was mainly expressed by fibroblasts (Figure 4(a) and (b), Supplemental Figure 5(C)), and fibroblasts obtained from severely fibrotic tissue exhibited higher IL11 expression (Figure 4(c)). Immunofluorescence analysis of full-thickness intestinal sections from stenotic regions in CD patients revealed the colocalization of IL11 with COL1A1, indicating its expression in intestinal fibroblasts (Figure 4(d)). Additional murine single-cell RNA sequencing data generated previously by our team using chronic DSS treatment were also investigated. After quality control, we retained a total of 56,470 high-quality cells. We identified nine main cell types through unsupervised clustering and classical marker gene annotation (Supplemental Figure 5(D) and (E)). UMAP plot and Dot plot showed that fibroblast was one of the main cell types expressing Il11 (Figure 4(e) and (f)). Besides, fibroblasts in the chronic DSS group exhibited higher Il11 expression than those in the control group (Figure 4(g)). Interestingly, Il11 was also highly expressed in murine endothelial cells and myocytes (Figure 4(f)). However, fibroblasts remained the largest population among the IL11-positive cells (Supplemental Figure 5(F)). Next, immunofluorescence of chronic DSS murine sections revealed that IL11 was colocalized with vimentin and podoplanin, indicating expression in the intestinal fibroblasts (Figure 4(h)).

IL11 was mainly expressed by fibroblasts. (a) UMAP plot shows the cells that express IL11 in the CD surgical specimens. The bigger black round dots represent the cells that express IL11 and the red dots represent the fibroblasts. (b) Dot plot shows the expression of IL11 in normal, and fibrotic tissue of CD surgical specimens. The average gene expression and percentage of cells expressing IL11 are shown by dot color and size, respectively. (c) Histogram shows the average expression of IL11 in fibroblast from normal and fibrotic tissue of CD surgical specimens. Statistical differences were determined by the Mann–Whitney U test. (d) IF shows the expression of IL11 (red color), and COL1A1 (green color) in full-thickness intestinal sections from stenotic regions in CD patients. (e) UMAP plot shows the cells that express Il11 in the chronic DSS murine model. The bigger black round dots represent the cells that express Il11 and the red dots represent the fibroblasts. (f) Dot plot shows the expression of Il11 in the control and chronic DSS group. The average gene expression and percentage of cells expressing Il11 are shown by dot color and size, respectively. (g) Histogram shows the average expression of Il11 in fibroblast from the control and chronic DSS group. Statistical differences were determined by the Mann–Whitney U test. (h) IF shows the expression of IL11 (red color), vimentin (blue color), and Podoplanin (green color) in chronic DSS murine sections.
Furthermore, we analyzed the expression of IL11 receptor alpha (IL11RA; the specific receptor for IL11) in human and murine single-cell RNA sequencing data. We found that fibroblasts constituted the largest population among IL11RA-positive cells, suggesting that IL11 likely plays a mainly functional role in fibroblast (Figure 5(a)–(d)). Primary intestinal fibroblasts were then isolated from CD patients. Without stimulation, we noticed that the cells isolated from stricture tissue of CD patients expressed more IL11 than those isolated from relatively normal and inflammatory areas of CD patients, indicating an autocrine loop (Figure 5(e)).

IL11 receptor IL11RA was expressed by fibroblasts. (a) Feature plot shows the expression of IL11RA in CD surgical specimens. (b) Pie chart shows the proportion of major cell types among IL11RA-positive cells in CD surgical specimens. (c) Feature plot shows the expression of Il11ra1 in the chronic DSS murine model. (d) Pie chart shows the proportion of major cell types among IL11ra1-positive cells in the chronic DSS murine model. (e) IF shows the IL11 expression in CDni-HIF, CDi-HIF, and CDs-HIF.
IL11 promoted the production of extracellular matrix
Lastly, we want to test the pro-fibrotic function of IL11 in human primary intestinal fibroblast. After 24 h stimulation with IL11, the production of extracellular matrix, such as fibronectin and collagen I, was increased (Figure 6(a)–(d)). Statistical analysis of protein expression by western blot demonstrated that the increase in fibronectin and collagen I expression was statistically significant (Figure 6(b) and (c)). Besides, we tested the cell proliferation ability using immunofluorescence of Ki67 and found that the number of Ki67-positive cells increased after IL11 stimulation (Figure 6(e) and (f)).

IL11 promoted the production of extracellular matrix. (a–c) Western blot shows the expression of pro-fibrotic proteins (FN and COL1A1) after 24 h stimulation with human IL11 (10 ng/mL). Results from four biological replicates. The protein expression of IL11-treated group was normalized by the control group from each primary fibroblast. Statistical differences were determined by paired t-test. (d) IF shows the expression of pro-fibrotic proteins after human IL11 stimulation. (e, f) IF of Ki67 shows the cell proliferation ability after human IL11 stimulation. Results from three biological replicates. Data are mean ± SEM. Statistical differences were determined by one-way ANOVA.
Discussion
In our study, IL11 was found to be increased in the fibrostenotic intestinal tissue and also in the serum of CD patients with type B2 or type B3. Clinical analysis showed that IL11 was associated with progressive disease behavior and CRP, and could predict the risk of surgery afterward and anti-TNF response. Furthermore, IL11 and its receptor IL11RA were mainly expressed in fibroblasts and could promote fibrosis in an autocrine loop.
Intestinal fibrosis and stricture are common complications of CD patients. According to an American population-based cohort study (n = 306), the cumulative probability of stricturing disease was 21.6% at 20 years after diagnosis, while that of penetrating disease was 37.1% at 20 years after diagnosis. 37 In addition, another study showed that half of the CD patients required surgery within 10 years after diagnosis, which imposes a significant burden on patients and their families. 38 Interestingly, penetrating CD often accompanies intestinal stenosis. 39 Thus, it is crucial to accurately identify and predict which patients will develop intestinal stricture. This helps to stratify higher risk patients and obtain timely treatment for a better prognosis. However, no ideal or reliable biomarkers can be used to detect intestinal fibrostenosis in CD.1,5
IL11 is a member of the interleukin-6 family. It forms a dimer by combining with its specific transmembrane receptor, IL11Rα, and then interacts with glycoprotein 130 to activate the downstream classical JAK-STAT3 or non-classical ERK and PI3K-AKT signaling pathways, thereby playing important pathophysiological roles in regulating cell proliferation, differentiation, migration, and angiogenesis.40–42 Early studies of IL11 mostly focused on the field of oncology.40,41 It has been shown that IL11 is mainly expressed and secreted by fibroblasts and is hard to detect in most healthy tissues and cells.18,41 However, its expression is upregulated in various cancer tissues, such as gastric cancer, colorectal cancer, and breast cancer, and it participates in the occurrence and development of these tumors. Therefore, some studies have regarded IL11 as a potential tumor therapeutic target.41,43 In 2017, a study published in Nature pointed out that IL11 promotes cardiac and renal fibrosis, and blocking IL11 can reverse the occurrence of fibrosis. 18 Subsequently, IL11 was also reported to promote the development of pulmonary fibrosis and liver fibrosis.44–47 To investigate the effect of IL11 on the gut, the same group from Singapore crossed the heterozygous Rosa26-Il11 mice with Col1a2-CreER mice to generate fibroblast-specific Il11 transgenic model (Il11Fib). The Il11Fib mice exhibited reduced colon length, upregulated expression of Il6, and increased inflammation. However, the colonic fibrosis was not significantly different when compared to controls. 48 In addition, the phenotype of Il11 conditional knock out mice is unclear, which needs further investigation in the future.
Since IL11 plays a role in fibrosis in the literature, we wonder if IL11 can be a potential biomarker for CD fibrostenosis. We focused on both the IL11 expression level in intestinal tissue and the serum of CD patients. Our data showed that IL11 increased in tissue from fibrotic/stenotic tissue and serum from stricturing/penetrating CD patients. These data indicated that IL11 could be a potential biomarker to recognize fibrostenosis in CD.
We noticed that the serum levels of IL17 and IL27 were increased in the group of type B3. IL17 was reported to play a role in the pathogenesis of fibrosis in several organs, such as lung, skin, heart, and liver.22,49–51 In terms of intestinal fibrosis, Biancheri et al. 52 found IL17A was significantly elevated in strictured CD tissues when compared with non-strictured CD tissues. Besides, Li et al. 53 confirmed that IL17 was elevated both in the serum and gut tissue in 2,4,6-trinitrobenzenesulfonic acid-induced intestinal fibrosis mouse model, while anti-IL17 antibody treatment significantly alleviated intestinal fibrosis. However, in clinical practice, IL17 blockers, which are widely used to treat psoriasis and ankylosing spondylitis, have paradoxical intestinal effects and have not been approved in CD patients. 54 IL27 belongs to the superfamily of IL6 and IL12, mainly produced by antigen-presenting cells. In 2011, IL27 was reported to be elevated in the serum of patients with systemic sclerosis, and IL27 stimulation increased proliferation and extracellular matrix synthesis of fibroblasts in patients with systemic sclerosis. 55 However, recent studies showed that IL27 could actually alleviate lung fibrosis.26,56 The role of IL27 in intestinal fibrosis of CD has not been reported yet, although IL27 interacts with different types of cells and plays a role in innate immunity, adaptive immunity, and ultimately intestinal homeostasis. 57 More researches are needed to clarify the role of IL27 in CD and particularly in intestinal fibrosis.
Our single cell RNA sequencing analysis showed that fibroblasts were the main cell type that expressed IL11 both in the intestinal fibrosis murine model and CD surgical sections, which was consistent with another team. 18 However, we could not exclude the effect of other cell types. In 2020, Lim et al. 48 showed that the transgenic murine model with smooth muscle cell-specific, conditional expression of IL11 exhibited an inflamed, fibrotic intestinal phenotype, which mimics inflammatory bowel disease. IL11 was also expressed in endothelial cells. In vitro and in vivo studies using conditional knock out mice are needed to test the function of IL11 in these cell types.
This study has some limitations. Firstly, this study is retrospective in design with relatively small sample size. We are unable to answer the question of whether the expression level of serum IL11 can predict progressive disease behavior (type B2 or type B3) or not, which has an important value in clinical practice. Besides, we could not determine whether a combined prediction model with IL11 (targeting both inflammation and fibrosis) can stably improve the overall predictive performance for surgical outcomes. Prospective larger cohorts with extended follow-up time and external validation will be critical to test these hypotheses and to further investigate the value of IL11 in the future. Secondly, we tested the fibrotic function of IL11 using primary fibroblast cell culture system. To confirm this finding, rescue experiments are needed, and in vivo model using conditional knock out mice or IL11-neutralizing antibody would be helpful to realize clinical translation. Lastly, this study mainly focuses on the role of various fibrosis-associated interleukins in CD intestinal fibrostenosis. Conventional pro-inflammatory cytokines, such as TNF, were not included in the analysis, which needs further investigation in future research.
Conclusion
In summary, our data indicated that IL11, mainly expressed by fibroblasts, could promote intestinal fibrosis and may be a potential biomarker for CD fibrostenosis.
Supplemental Material
sj-docx-1-tag-10.1177_17562848251391093 – Supplemental material for Fibroblast-derived interleukin-11 as a potential biomarker for intestinal fibrostenosis in Crohn’s disease
Supplemental material, sj-docx-1-tag-10.1177_17562848251391093 for Fibroblast-derived interleukin-11 as a potential biomarker for intestinal fibrostenosis in Crohn’s disease by Jinyu Tan, Xiaomin Wu, Weidong Huang, Jinshen He, Jing Nie, Longyuan Zhou, Zishan Liu, Yu Wang, Caiguang Liu, Jie Li, Yao Zhang, Minhu Chen, Ren Mao and Shanshan Xiong in Therapeutic Advances in Gastroenterology
Footnotes
Acknowledgements
We used the artificial intelligence tools such as Doubao and Deepseek for natural language processing on the Abstract section, which enhanced the clarity of our manuscript. The Associate Editor of Therapeutic Advances in Gastroenterology (Ren Mao) is an author of this paper; therefore, the peer-review process was managed by alternative members of the board, and the submitting editor had no involvement in the decision-making process.
Declarations
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.