
Abstract
Otitis media (OM) is the most common disease among young children and one of the most frequent reasons to visit the pediatrician. Development of OM requires nasopharyngeal colonization by a pathogen which must gain access to the tympanic cavity through the eustachian tube (ET) along with being able to overcome the defense mechanisms of the immune system and middle ear mucosa. OM can be caused by viral or bacterial infection. The three main bacterial pathogens are Streptococcus pneumoniae, nontypeable Haemophilus influenzae (NTHi), and Moraxella catarrhalis. Innate immunity is important in OM resolution as the disease occurs in very young children before the development of specific immunity. Elements of innate immunity include natural barriers and pattern recognition receptors such as Toll like receptors (TLRs), and Nod like receptors (NLRs). Surfactant proteins A (SP-A) and D (SP-D) act as pattern recognition receptors and are found in the lung and many other tissues including the ET and the middle ear where they probably function in host defense. Surfactant has a potential for use in the treatment of OM due to surface tension lowering function in the ET, and the possible immune functions of SP-D and SP-A in the middle ear and ET.
Introduction
Otitis media (OM) is the most common disease in young children and one of the most frequent reasons to visit the pediatrician.1, 2 It is estimated that about 60%-80% of infants will have at least one episode of OM. 3 Acute otitis media (AOM) is inflammation of the mucosa of the middle ear cleft which includes the mastoid process, tympanic cavity and, the Eustachian tube (ET) . 4 There is a significant global health burden of AOM; Thirty one million children of an estimated 709 million cases per year progress to develop chronic suppurative OM (OM with chronic ear discharge), and complications such as meningitis or brain abscess which result in about 21,000 deaths each year. 5 The World Health Organization estimates that 51,000 deaths/year are attributable to AOM in children younger than 5 years old and that chronic otitis media is the major cause of hearing loss in developing countries. 6 Studies show that before their third birthday, 80% of children will have suffered a minimum of one attack of OM and 40% will have at least six recurrent attacks by the age of 7 years. OM is also the primary indication for prescribing antibiotics among children. 7 In the first two years of life, children spend a mean of 90 days on antibiotics for OM. 8 Additionally, OM is the primary indication for ventilation tube insertion, which is the most commonly performed operation in children. 7
Pathogenesis
There are three requirements for developing OM: 1) nasopharyngeal colonization by the pathogen ; 9 2) ascending infection through the ET to the tympanic cavity; 3) pathogens must be able to overcome the defense mechanisms of the immune system and middle ear mucosa .10, 11 A multitude of host and environmental factors significantly influence the risk of developing OM. Host factors that can increase the risk of developing OM include, sex, young age, genetic susceptibility, adenoid hypertrophy, laryngopharyngeal reflux, race and ethnicity, and craniofacial malformations such as cleft palate, atopy, immunodeficiency, and viral upper respiratory tract infections (URTIs). 12 Family history of OM may be an especially important factor. 13 Environmental factors that increase the risk of OM include: low socioeconomic status, exposure to tobacco smoke, having older siblings, day-care attendance and the use of a pacifier. 14
Viral and bacterial pathogens induced OM
Acute otitis media can result from viral or bacterial infections. Viral infection cause around 20% of AOM. Respiratory syncytial virus (RSV), influenza viruses, adenoviruses, rhinoviruses, and enteroviruses, are the viruses that most commonly cause AOM.15, 16 A recent study by Heikkinen et al., showed that among children younger than three years, the average RSV infection incidence rate was 275 per 1000 children per year and 58% of these children with RSV developed AOM . 17 Bacterial infection is the predominant cause of acute and recurrent otitis media. It leads to hyperplasia of the tympanic epithelium, middle ear effusion and leukocytic infiltration of the tympanic cavity.18, 19 There are three main bacterial pathogens, Streptococcus pneumoniae, nontypeable Haemophilus influenzae (NTHi), and Moraxella catarrhalis.10, 20 Following the introduction of vaccines against S. pneumonia, NTHi has become the most common pathogen in OM.19, 21 The risk for developing AOM after upper respiratory viral infection depends on the otopathogenic bacteria colonizing the nasopharynx; the risk is highest if the nasopharynx is colonized by all three pathogenic bacteria and lowest with no colonized bacteria. 22 Viral infection negatively impacts the nasopharyngeal mucosa and the ET functions. Viral infection changes the nasopharyngeal mucosa by modifying host immune function, inducing cytokine activity and inflammatory mediators and upregulates host cell surface antigens that act as bacterial receptor sites and increases bacterial colonization and adherence. Viral infection impair the ET through altering the properties of mucus and diminishing the normal mucociliary clearance by mucosal cells of the ET. Transient impairment of ET functions allows bacteria colonizing the nasopharynx to ascend into the middle ear and cause AOM.14, 23 Using chinchilla otitis media model, studies shown that inoculation with S. pneumoniae alone through the nasopharynx resulted in the development of OM in 20% of the animals, whereas adding adenovirus to the bacteria led to the development of OM in nearly 80% of them. 24 Initially, M. Catarrhalis adheres to mucosal surfaces through a dozen or more adhesins that it expresses. However, in AOM, adherence to the mucosal surface alone is not sufficient to cause disease. A cofactor, such as a viral infection, is thought to be needed to precipitate migration to the ME through the Eustachian tube. M. catarrhalis is often isolated with S. pneumoniae and H. influenzae in respiratory tract cultures and may facilitate polymicrobial infection by sheltering these organisms from complement-mediated immune destruction, promoting biofilm formation, and releasing beta-lactamase into the local environment.25, 26 M. catarrhalis has shown the ability to overcome host innate immune response and to evade complement-mediated lysis.
Structure and function of the middle ear and ET
The middle ear cleft is comprised of the tympanic cavity, the mastoid air cell system and ET. The middle ear cleft is an irregular shaped gas filled chamber that uses the ET to equalize middle ear pressure with the ambient pressure through the nasopharynx 27 (Figure 1). The middle ear mucosa is derived from two origins, neural crest which give origin to the dorsal region of the middle ear mucosa which is a non-ciliated epithelium and endoderm of the first pharyngeal pouch which give rise to the ventral part of the mucosa forming the ciliated epithelium near the ET orifice. 28 Embryologically, the ET develops from the first pharyngeal pouch and connects the nasopharynx to the tympanic cavity. The nose, palate, nasopharynx, ET, middle ear and mastoid air cells form a system of contiguous organs. The ET is not a tube in reality, but an organ consisting of a lumen with its mucosa, cartilage, surrounding soft tissue, and peritubal muscles. 29 The ET functions to aerate and protect the middle ear from excessive changes in the atmospheric pressure, also as a mucus drainage pathway, and to protect the middle ear from otopathogens and other inhaled noxious agents. 30 The ET is lined by respiratory epithelium, which is pseudostratified ciliated columnar epithelium with interspersed goblet cells, that produce both mucoid and serous mucus. The cilia propel mucus in combination with epithelial secretion of antimicrobial protein, through the ET, from the middle ear to the nasopharynx which help to protect against bacterial colonization of the middle ear. 14 The ET epithelium is the frontline defense against the passage and colonization of otopathogens from the nasopharynx.14, 31 ET dysfunction is the primary cause of middle ear infection and effusion. 32 Poor or impaired function of the mucociliary system of the ET can result in bacterial entry into the tympanic cavity. OM is more common in infants which can be explained anatomically. The ET is shorter, wider and more horizontal in infants and young children (<1 year of age) than in adults, which facilitates otopathogen transmission to the middle ear increasing the opportunity for antigen exposure or pathogen colonization while reducing the opportunity for mucosal immune responses thus increasing the risk of OM, compared to the adult morphology14, 33
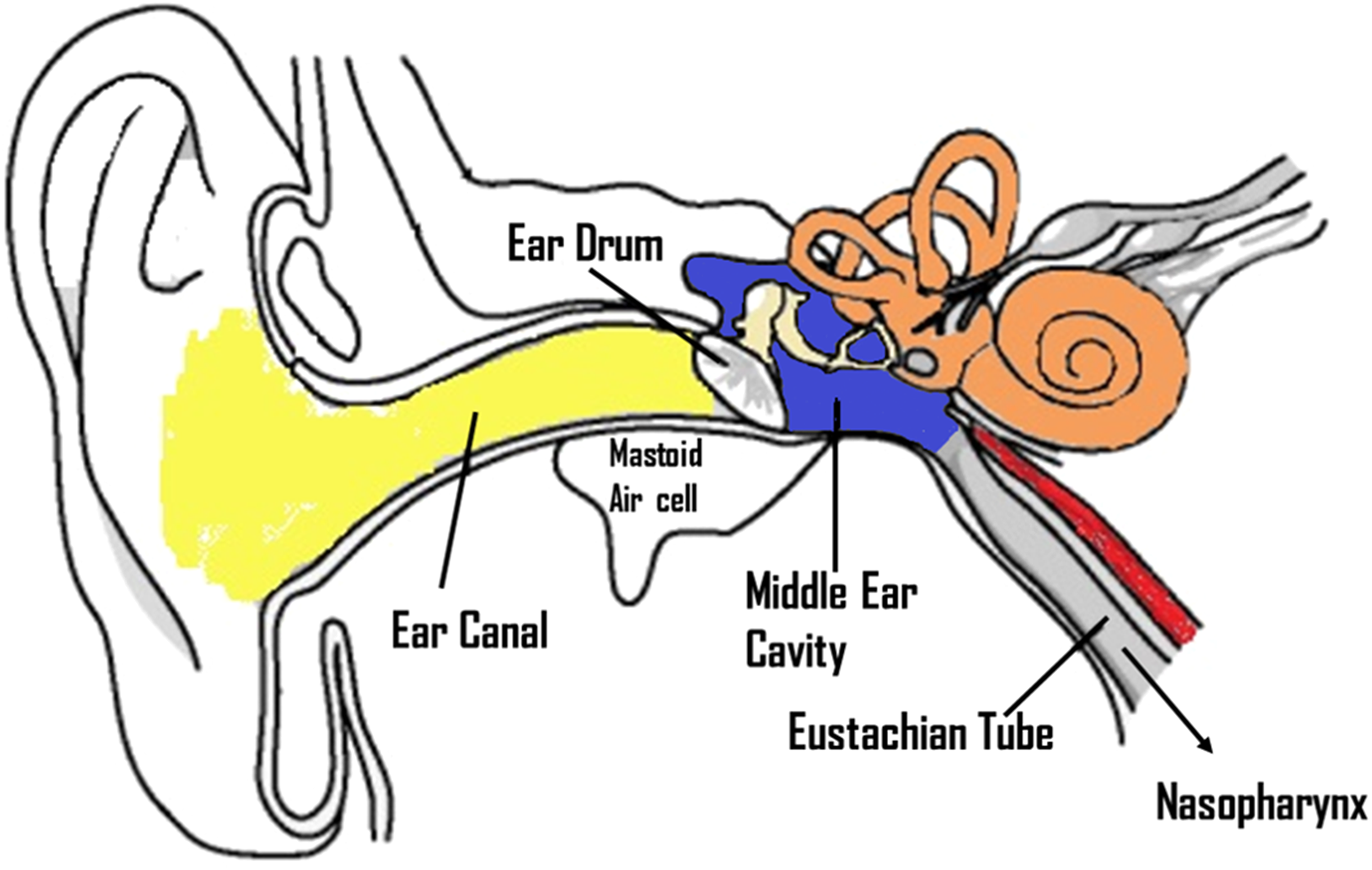
Schematic representation of ear structure showing the external ear (Yellow), the middle ear (blue) and the inner ear (orange). The middle ear space connects to eustachian tube and the mastoid air cell system.
Innate immune response in OM
The innate immune response is a nonspecific, first line defense against pathogens that does not require prior sensitization (see Table 1). There are diverse players that contribute to the innate immunity (Figure 2) in the middle ear and ET. These innate immune elements can be classified as natural barriers, pathogen recognition receptors, antimicrobial proteins, and peptides, mucociliary system and various cellular components. Natural barriers function through mucous membranes and mucociliary system that employs a bilayer stream of mucus blanket with a thick mucus layer secreted by goblet cells and submucosal glands traveling on top of a thin serous layer secreted by the epithelial cells. Together these elements constitute the periciliary fluid. Both layers cover the middle ear and ET mucosa and are important for the proper functioning of the mucociliary system. 34
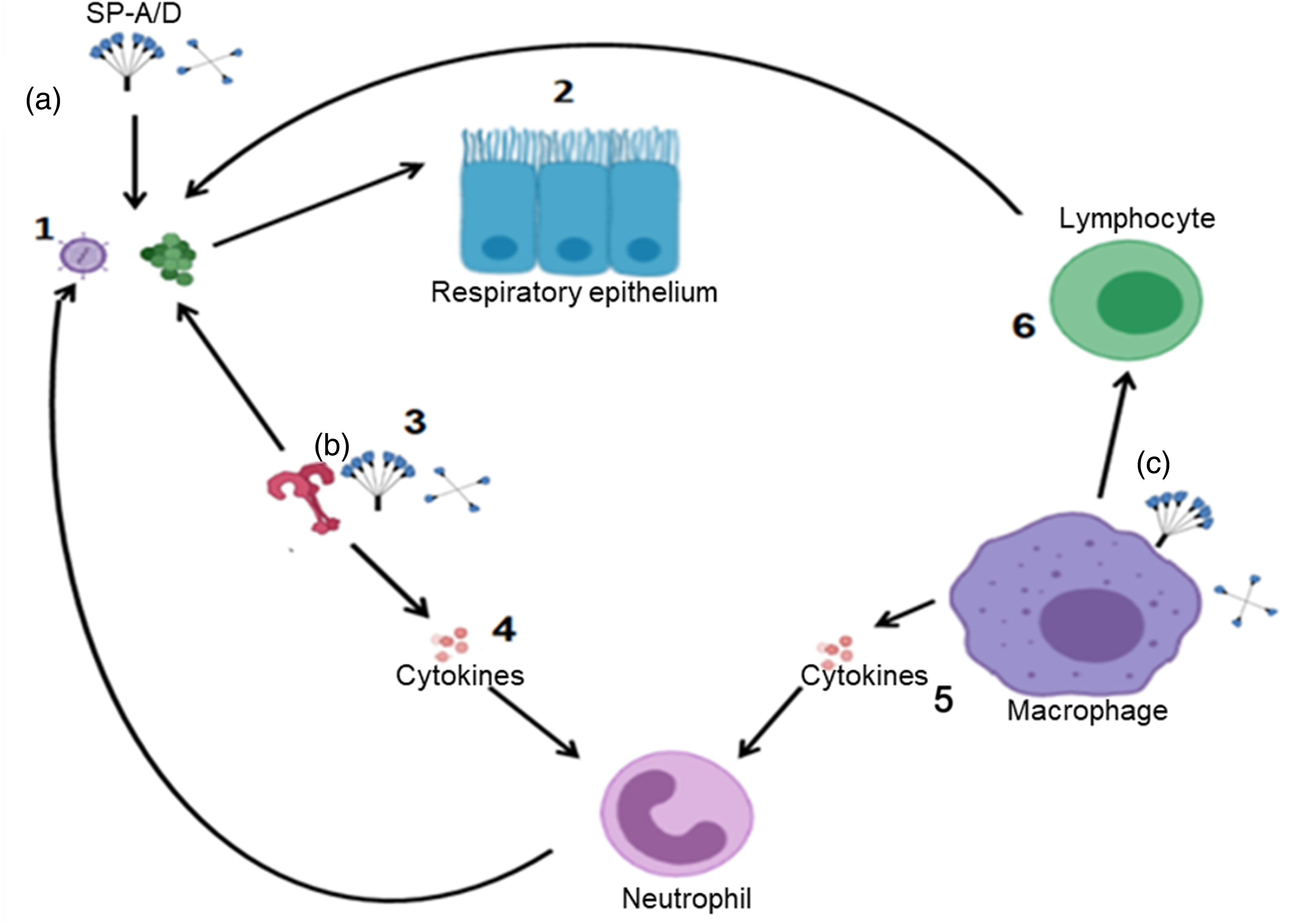
Schematic representation of the major players in the innate immune system. (1) pathogens attack the host; (2) Physical and chemical barriers as epithelium with the mucociliary system; (3) Pathogen recognition receptors as TLRs; (4) activation of cytokines which attract innate immune cells; (5) to attack the pathogens. Macrophage help to (6) activate adaptive immunity. SP-A and SP-D can attack pathogens directly (a) act as PRR (b) or enhance phagocytosis (c).
The innate immune system recognizes the presence of microbial infection by using pattern recognition receptors (PRRs) to identify pathogen-associated molecular patterns (PAMPs), which are the molecular signature of pathogens . 35 Toll-Like receptors (TLRs) play a key role in innate immunity. They are a class of PRRs recognizing molecules associated with microbial pathogens.36, 37 Recently, TLRs have emerged as key regulators of innate immune responses to infection in mammals. TLRs are found in the cytoplasm or on the cell membranes of the epithelial cells lining the middle ear, and also on mast cells and dendritic cells, which are abundant at middle ear mucosal surfaces. Contact with PAMPs leads to activation of the TLRs, which activate a signaling cascades that regulate innate immune response to mobilize cytokines, chemokines and interferons which attract neutrophils and macrophages to clear the site of infection from the invading bacteria and can then stimulate both local and systemic adaptive immune responses (Figure 2).37, 38 There have been thirteen different members of the mammalian TLR family identified (10 in humans and 12 in mice) . 39 Among the known 10 members of human TLRs, TLR2 is activated by pneumococcal cell wall components, such as lipoteichoic acid and lipoproteins and likely plays an important role in the pathogenesis of pneumococcal OM.38, 40 TLRs activate an intracellular signaling cascade through employing adapter proteins. Except for TLR3 all TLRs employ myeloid differentiation factor-88 (MyD88) as an adapter protein to induce nuclear factor-kappa B (NF-κB) or mitogen-activated protein (MAP) kinase–dependent proinflammatory gene expression to the production of proinflammatory cytokines such as tumor necrosis factor (TNF-α) and the interleukins (ILs).39, 40 Recovery from OM is impaired by the absence of key elements of TLR signaling, particularly TLR2, MyD88, and TNF- α. 37 In children with recurrent otitis media, the levels of IL-1β, IL-6, and TNF-α in nasopharyngeal secretions were found to be lower than in healthy children.6, 41 TLRs work by recognizing pathogen associated molecular patterns. The interactions between TLRs and these molecular patterns can activate intracellular signaling pathways, such as the NF-κB pathway. The activation of NF-κB pathway in turn upregulates the expression of proinflammatory genes involved in the production of cytokines and chemokines and activation of the adaptive immune system. TLR2, a member of TLR family, reported to regulate the pathogenesis of NTHi induced AOM as it recognizes pathogenic patterns of NTHi and regulates the inflammatory responses of the host 42 Lipoprotein P6 of the NTHi uses TLR2 signaling to activate NF-κB in human epithelial cells. Similarly, TLR4 mediates lipopolysaccharide (LPS) and lipooligosaccharide (LOS) responsiveness and recognizes gram-negative bacteria via the LPS/LOS moiety present on the surfaces of these pathogens. Although the structure of LOS and LPS is different, lipid A, which is a common component to both, is thought to be responsible for TLR4 signaling. 43 LPS-TLR4 signaling and neutrophil reduced nicotinamide adenine dinucleotide phosphate oxidase activate NF-κB signaling and upregulate TLR2 expression in endothelial cells, and this increased TLR2 expression via NF-κB signaling results in increased intercellular adhesion molecule 1 expression and enhanced neutrophil migration. TLR4 signaling appears to induce TLR2 expression, and TLR2 activation is critical for bacterial clearance and rapid resolution of OM. 44 A recent study demonstrated that the TLR4 gene locus, regulating the innate immune response, influences the genetic predisposition to childhood OM in a subpopulation of patients. 45 TLR4 Polymorphisms have been found to associate with the risk of recurrent OM, but the data are partly conflicting. 46 Additionally, TLR9 is localized to endosomes and when exposed to bacterial DNA it recognizes a common motif present in bacterial but not mammalian DNA, unmethylated cytidine-phosphate-guanosine (CpG). This results in signaling molecules recruitment, that eventually result in the production of pro-inflammatory cytokines and other target genes expression. TLR9 facilitates the development of the inflammasome, which might contribute in the development of effective adaptive immune response. 31 Leichtle et. al. recently showed that deletion of TLR9 significantly prolonged the inflammatory response induced by NTHi in the ME and delayed bacterial clearance. 31
Other PRRs including cytoplasmic (NOD)-like receptors (NLRs), and C-type lectin receptors (CLRs) are found to be involved in the innate immunity of OM . 7 The NOD-like receptors (NLRs) are cytoplasmic proteins that regulate inflammatory and apoptotic responses. The NOD proteins NOD1 (which is encoded by the caspase-recruitment domain 4 gene, CARD4) recognizes a molecule called meso-diaminopimelic acid, a peptidoglycan constituent only of Gram negative bacteria. NOD2 (which is encoded by CARD15) recognize intracellular muramyl dipeptide (MDP), a peptidoglycan constituent of both Gram positive and Gram negative bacteria.47, 48
Defensins are cationic proteins released by the middle ear epithelium and function as antimicrobial agent with a wide range of activity against viruses, bacteria, fungi and protozoa. Defensins can also upregulate cytokines to attract neutrophils, mast cells, and T-lymphocytes. 49 Lysozyme is an antimicrobial molecule secreted by epithelial cells in the airway and act by degrading the peptidoglycan in the bacterial cell wall. 50 Special inflammatory and immune-response-relevant cells such as mucosal dendritic cells (DCs) and mast cells have been found to play a role in the pathogenesis of OM.
Surfactant proteins and innate immunity in OM
Surfactant proteins (SPs) represent a small percentage of surfactant composition but they are essential for surfactant homeostasis. 51 Four surfactant proteins, named as SP-A, SP-B, SP-C, and SP-D, have been discovered and characterized in lung biology. SP-B and SP-C are hydrophobic proteins, and play a key role in lowering surface tension, 52 as well as surfactant metabolism and recycling in the lung. 51 SP-A and SP-D belongs to the family of C-type lectins, or collectins, they are hydrophilic proteins and mainly contribute to the innate immunity.53–55 At initial studies, all surfactant proteins are synthesized and secreted by alveolar type II cells and SP-A and SP-D were also found to be secreted by sub-mucosal and Clara cells in the lung. SPs are present at the luminal surface of pulmonary epithelial cells, and are secreted into alveolar spaces.56–58 Several recent studies have demonstrated that SP-A and SP-D are expressed in multiple tissues other than the lung, including the tongue and oral epithelium; the digestive, urinary and reproductive tracts; synovial and pericardial fluid; the spleen, thymus, pancreas, kidney, the middle ear and Eustachian tube.30, 59 Although SP-B and SP-C were thought to be expressed solely in the pulmonary epithelium,60–62 SP- B and various phospholipids were found also expressed in the eustachian tube. Therefore, all main components of pulmonary surfactant are produced in this tissue by fully competent cells having a complete surfactant machinery. 63
The innate immune response is very important for susceptibility to OM in young age before acquiring specific immunity. 3 Collectins protect the host through the identification of PAMPs on pathogens and allergens and respond by activation of multiple processes of innate immunity, including phagocytosis, cytokine secretion, and complement activation . 53 SP-A and SP-D play an important role in host defense through opsonization and complement activation (Figure 3). 64 In a recent study by our group we found that SP-A mediated NTHi aggregation and killing along with enhanced bacterial phagocytosis by macrophages in vitro and modulated inflammation of the ME in otitis media in vivo. 59 In another study (results not published yet) we found that mice lacking SP-D gene had prolonged inflammation and slow resolution of the ME inflammation in response to NTHi induced otitis media.
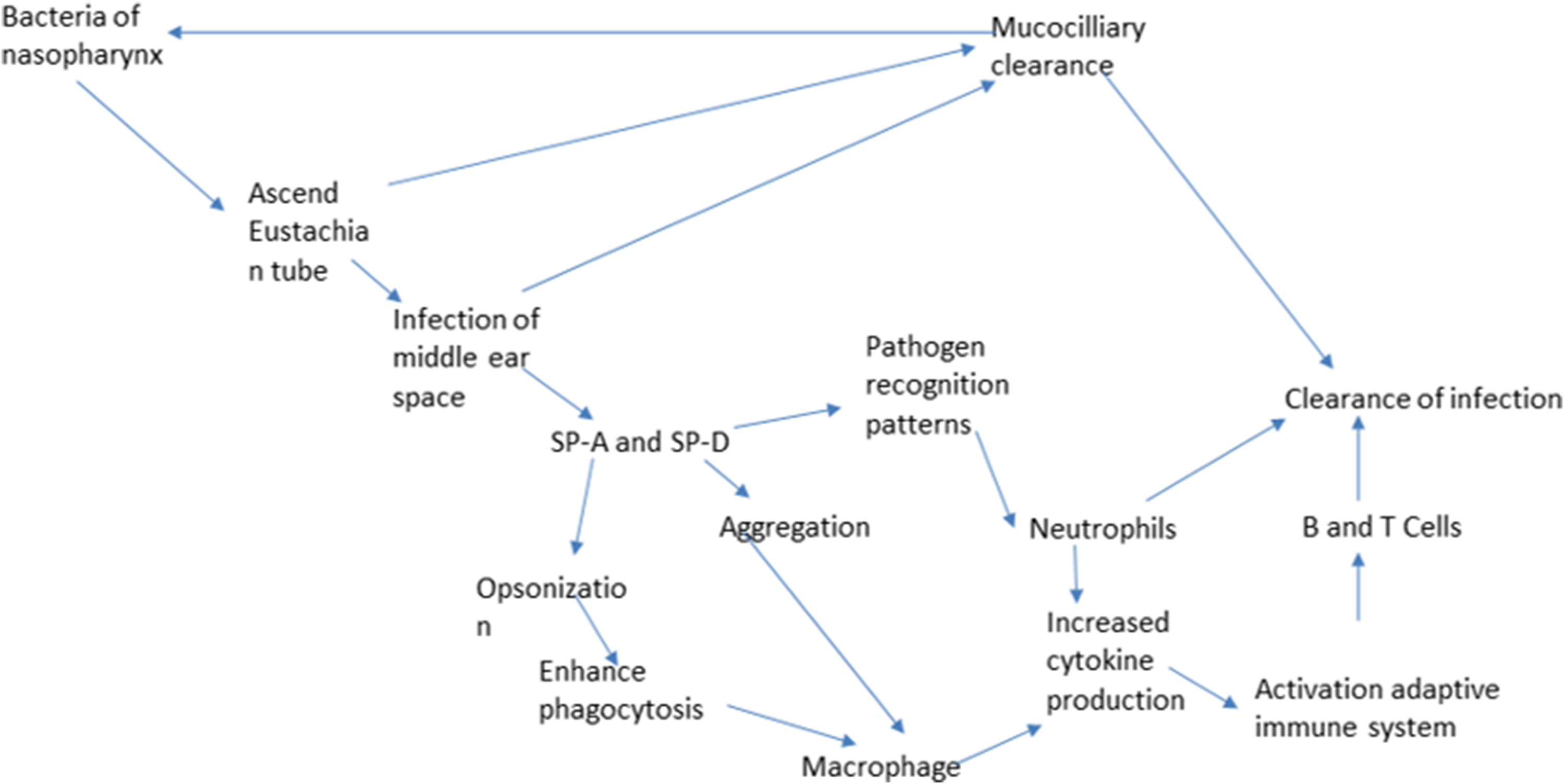
Schematic representation of the pathogenesis of AOM and the roles of SP-A and SP-D in the innate immune response of OM.
Collectins are structurally and functionally related to the first component of the classical complement pathway C1q, with the exception that collectins are exclusively present in the extracellular matrix.65–67 Collectins are formed of collagen like regions associated with non-collagen domains. The carbohydrate recognition domain (CRD) is the part that recognizes carbohydrate epitopes from multiple microorganisms. CRD is attached to the neck and collagen domain which is attached to N-terminal domain. 68 CRD of collectins can differentiate between self and non-self carbohydrates. For example, all collectins have affinity to mannose, SP-A has affinity to L-fructose and N-acetylmannosamine, and SP-D to glucose, mannose and inositol. On the other hand, they poorly recognize galactose and sialic acid which make up clusters of oligosaccharides in most vertebral animals. 58 Direct agglutination or neutralization of microorganisms can occur when they bind to collectins. This agglutination provides a first line of defense, which can be augmented by killing and clearance mechanisms mediated by phagocytic cells that carry receptors for SP-A and SP-D. 11 Collectins also help in opsonization and the presentation of bound microbes directly to phagocytes, or activate complement cascade via the lectin pathway. The lectin domains recognize and bind to carbohydrate structures found on a variety of microorganisms such as viruses, bacteria, yeast and fungi. The collagenous regions are ligands for the collectin receptor on phagocytes and also mediate C1q-independent activation of the classical complement pathway.69, 70 SP-A enhances production of secretory leukoprotease inhibitor (SLPI). SLPI is a 12 kda protein that is a constitutively expressed, up-regulatable inhibitor of serine proteases. It is found in airway surface epithelial cells, Clara cells, and associated with elastin fibers of the lung interstitium and monocytes, alveolar macrophages and neutrophils. SLPI targets neutrophil elastase, cathepsin G, chymotrypsin and trypsin. It has antimicrobial properties against bacteria and it regulates the production of TNF-α by inhibiting the LPS-induced NF-kB activation and inhibiting IkB- α degradation. 71
Because SP-A works as a pattern recognition receptor, it functions as one of the first lines of defense before the development of specific antimicrobial antibodies. SP-A binds to and increases the phagocytosis of S. pneumoniae and H. influenzae, the most common otopathogens.3, 61 SP-A opsonizes gram negative bacteria and modifies LPS for macrophage binding. It also modulates proinflammatory cytokines IL-1B, IL-6, and TNF-α which play an important role in fighting infections. SP-A and SP-D are expressed in the normal human and porcine eustachian tube. 54 SP-A is the most abundant surfactant associated protein in the body and also in the ET, where it is synthesized most profusely at the mucosal folds of the tubal floor, where ciliated cells predominate. The role played by surfactant in the pathobiology of OM is complex and entails both protein and lipid fractions. 63 Phosphatidylcholine (PC) is the same primary phospholipid which is the found in lung as well as ET surfactant, but the difference is in the secondary phospholipid content. As in the lung, phosphatidylcholine (PC) is the main component of ear surfactants, however, phosphatidylethanolamine was higher and phosphatidylglycerol was undetectable in the middle ear .63, 72
Regarding PC to sphingomyelin ratio, ET lavage had a 2:1 ratio versus a 67:1 ratio found in pulmonary lavage. Variations in phospholipid content may be related to middle ear pathology. Svane-Knudsen et al. examined the concentrations of PC in lung and ET aspirates of normal subjects versus patients with secretory OM. They found that PC lung concentrations are the same in both groups, but they found a significant decrease in PC concentration in ET in patients with OM. Surfactant phospholipids did not significantly reduce surface tension in the air/ME mucosa interface, but they have a cleansing function, facilitating the opening of the ET and mucociliary transport, thus reducing bacterial invasion63, 72
The ET surfactant appears to be an anti-adherent agent, rather than a surface tension lowering agent. As such, it may be perfectly suited to the structure and function of the eustachian tube. 60 Although the roles of SP-A and SP-D in the defense of the tubotympanum remains to be proven, it is likely that a deficiency in these host defense molecules may contribute to the pathogenesis of OM. 73
Adaptive immunity of OM
Immune system of mucosal surfaces can be divided into inductive site and effector site. In inductive sites, macrophages and dendritic cells process antigens and present it to naïve B or T lymphocytes, whereas the transformation of B cells to antibody producing plasma cells takes place in effector sites. Inductive sites are organized in mucosal-associated lymphoid tissue (MALT), which is known more specifically in the nasal area as nasal associated lymphoid tissue (NALT). NALT consists of the adenoids and palatine tonsils which form Waldeyer’s ring in humans.74, 75 Recently, respiratory microfold (M) cells were identified in the murine airways. 76 Respiratory M cells were capable of sampling inhaled bacterial antigens to initiate an Ag-specific immune response. It is widely accepted that NALT M cells are key players in the uptake of nasally delivered antigens for the subsequent induction of antigen-specific IgA immune responses.77, 78 If these cells are present in the middle ear, they may result in the induction of a specific antibody response within the middle ear, particularly in patients who have chronic OM where germinal centers can be identified in the middle ear and ET epithelium.79, 80 The main antibody that protect the mucosa of the middle ear is the secretory IgA and its presence highlights that the ME mucosa is an effector site of the mucosal immune system. 79
Genetic susceptibility of OM
Due to the high prevalence of OM, and the consequences of recurrence and chronicity of the disease, it is very important to understand its genetic background and susceptibility. Genetic differences could result in differences in anatomy of structures such as ET or the nasopharynx or differences in immunological factors such as cytokines and mucins. 13 Genetic studies of OM are based on twin studies, genome-wide linkage studies and genome-wide association studies (GWAS). In a monozygotic twins and triplets study, heritability was estimated to be 0.73 in both males and females (p < 0.001).13, 81 Genome-wide linkage studies (GWLS) identify regions of the genome that harbor disease susceptibility loci by typing microsatellite markers or single nucleotide polymorphisms (SNPs) spaced across the genome in sets of affected relatives. 81 An evaluation of 588 patients undergoing tympanostomy tube insertion for chronic or recurrent OM with DNA analysis, three important chromosomal regions were identified as important influencers, 10q, 19q, and 3p. 13 Casselbrant et al. used the Genetic Relationships across Implicated Loci tool to identify possible candidate genes within their linkage regions. This identified a cluster of chemokine genes on 17q12 and several surfactant protein genes near 10q22.3. Possible candidate genes at these sites are surfactant protein gene SFTPA2 in the 10q22.3 region. 82 The human SP-A locus in chromosome 10q22-q23 consists of 2 very similar genes, SP-A1 and SP-A2. Both gene products are required for fully functional SP-A protein. Several alleles that differ by single amino acid have been identified for each gene. The frequency of specific surfactant protein-A haplotypes and genotypes differs between children with recurrent otitis media compared with a control population. 61 Using a candidate gene approach, Ramet et al. reported an over-representation of the 6A4–1A5 haplotype in children with recurrent otitis media and in children diagnosed with their first episode of AOM before 6 months; there was also an underrepresentation of the 6A2–1A0 haplotype in the latter subgroup.61, 83 A study by Wiertsema et al. on Polymorphisms of Mannose-binding Lectin (MBL) in relation to AOM found that MBL variant type was associated with an increased number of otitis media at 12 to 24 months of age. 84
Potential use of surfactant and surfactant proteins in the treatment of otitis media
The intranasal instillation or local nebulization of natural surfactants has a strong background and preclinical evidence showing positive effects on OM. 63 It is conceivable to suggest the administration of exogenous surfactant will improve the ET function by reducing the surface tension, which helps in opening the ET. Many studies have investigated a possible role for pulmonary surfactant in the treatment of OM with effusion (OME). Some of these studies have shown that using surfactant reduced the resolution time for OME.85–87 A recent study also showed that surfactant treatment for OME restored ET function by reducing the opening pressure and increasing ET compliance through the reduction of the mucosa-air surface tension. 88 The study also showed improvement of hearing threshold as well as improvement in the ET mucosa and ciliary morphology compared with saline treated guinea pigs. 88 Another study showed that the administration of intranasal surfactant in OM chinchilla model lead to decrease in the severity and duration of infection. 32
Current treatment modalities for otitis media
Watchful waiting (WW), and withholding antibiotic therapy in mild to moderate AOM episodes is advocated by most guidelines. Antibiotic treatment is advised to be reserved for more severe cases.14, 89 Children should be carefully monitored by caregivers with clear instructions to return to the doctor if symptoms persists or the child’s condition worsened .
90
Usually, by the end of the first day after diagnosis, 61% of children with AOM have decreased symptoms, increasing to 80% after 2 to 3 days with or without antibiotics treatment, and approximately 75% have resolution of their symptoms by the end of the first week.
91
Assessment and management of pain is an essential part of AOM treatment and for this, anti-inflammatory analgesics may be warranted. Topical and oral antibiotics are not recommend in all cases, immediate antibiotic treatment of AOM is recommended in very young children <6 months, children with craniofacial anomalies or who are immunocompromised, as well as those with severe illness due to AOM.
92
Suppurative complications tend to occur whether initial antibiotics are provided
Future directions
Vaccination efforts constitute some of the important future directions in the prevention of OM and include targeting viruses associated with OM 23 and targeting pneumococci through the development of new vaccines using pneumococcal proteins, targeting M. catarrhalis, 95 and development of noninvasive methods to immunize against96, 97 Some of the future directions in surgery include The Acclarent, an automated ventilation tube insertion system which is a safe and effective device for placement of tympanostomy tubes in patients with chronic OME and recurrent AOM. 98 There is also a novel biodegradable drug-eluting ofloxacin-loaded ventilation tube with sustained drug release technology that has been tested to potentially treat chronic OME patients. 94 Balloon dilation of the ET is another treatment modality that improves the management of OME patients. This technology hopefully will be applied in the future for the treatment of children with chronic ET dysfunction. 99
Innate immune components and their acting mechanisms.

Footnotes
Author contributions
O.A. reviewed publications and drafted the manuscript. J.A. edited and made one figure. D.S.P edited the manuscript. G.W. reviewed publications and wrote the manuscript.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported in part by NIH R01HL136706, NSF research award (1722630), and one Award from Clark Endowment for Pediatric Research Fund (G.W.).