
Abstract
To investigate the effects of surfactant proteins A and D (SP-A and SP-D, respectively) in urinary tract infection (UTI), SP-A and SP-D double knockout (SP-A/D KO) and wild type (WT) C57BL/6 female mice were infected with uropathogenic Escherichia coli by intravesical inoculation. Compared with WT mice SP-A/D KO mice showed increased susceptibility to UTI, as evidenced by higher bacterial CFU, more infiltrating neutrophils and severe pathological changes. Keratinocyte-derived chemokine increased in the kidney of WT mice but not in SP-A/D KO mice 24 h post-infection. Compared with control, the level of IL-17 was elevated in the kidney of infected WT and SP-A/D KO mice and the level of IL-17 was higher in the infected SP-A/D KO mice than in infected WT mice 24 and 48 h post-infection. The basal level of p38 MAPK phosphorylation in SP-A/D KO mice was higher than in WT mice. The phosphorylated p38 level was elevated in the kidney of WT mice post infection but not in SP-A/D KO mice. Furthermore, in vitro growth of uropathogenic E. coli was inhibited by SP-A and SP-D. We conclude that SP-A and SP-D function as mediators of innate immunity by inhibiting bacterial growth and modulating renal inflammation in part by regulating p38 MAPK-related pathway in murine UTI.
Introduction
Urinary tract infection (UTI) is one of the most common infectious diseases, causing significant morbidity and mortality. 1 Approximately 40% of women and 12% of men experience at least one symptomatic UTI in their lifetime. As many as 25% of women experience recurrent UTI within 6 to 12 mo. 2 UTI causes more than 11 million physician visits and almost half a million hospitalizations, resulting in an estimated cost of $3.5 billion annually in the US alone. 2 Uropathogenic Escherichia coli (UPEC) is the most frequent pathogen of asymptomatic bacteriuria and symptomatic UTIs. 3
Recent studies highlight the importance of innate immunity in the development of UTI.4–6 When E. coli and other pathogens overcome various physical barriers by adhering to the epithelium, a robust innate immune response in the epithelial cells is generated.2,7,8 The effectors of this response include host defense proteins, antimicrobial peptides, cytokines and chemokines that attract phagocytes to the threatened site and enhance their microbicidal capacity and phagocytosis. 9
Surfactant proteins A and D (SP-A and SP-D) are members of the C-type lectin family that share a collagen-like region and a calcium-dependent globular carbohydrate-recognition domain (CRD). 10 SP-A and SP-D play an important role in the pulmonary innate immune system and protect the lung against various pathogens.11,12 They interact directly with a variety of pathogens, inhibit their growth and enhance clearance by phagocytic cells, 13 including E. coli K-12, 14 Mycoplasma pneumonia, Klebsiella pneumoniae and Histoplasma capsulatum.15,16
SP-A and SP-D function as PRRs by binding to receptors on the surface of epithelial and phagocytic cells, 17 thereby modulating inflammatory processes via inflammatory signaling pathways like NF-κB pathway. 18 Under normal environmental conditions in the lung, SP-A and SP-D bind to signal regulatory protein α by their globular CRD heads, which inhibits p38 MAPK activation and attenuates the production of various inflammatory mediators. 18 In the presence of microbes, the globular CRD domains of SP-A and SP-D interact with molecules on the surface of the microbes, 19 and the collagenous tails of SP-A and SP-D bind to calreticulin/CD91 of cells, thereby stimulating p38 MAPK phosphorylation and NF-κB activation, and resulting in increased cytokine and chemokine production (e.g. TNF-α, IL-8 and MCP-1). 18 SP-A and SP-D also interact with TLRs (TLR2 and TLR4) and TLR4 adaptor MD-2 via their CRD domains, 15 thus interfering with the interaction of pathogens and TLR receptors. 20
SP-A and SP-D knockout (SP-A and SP-D KO, respectively) mice have shown increased susceptibility to lung infection by various pathogens, including Pseudomonas aeruginosa, respiratory syncytical virus, M. pneumonia, K. pneumoniae and Pneumocystis carinii.17,21 As there is functional similarity and overlap between SPA and SP-D in host defense, SP-A and SP-D double KO (SP-A/D KO) mice were generated. 22 SP-A/D KO mice are more susceptible to P. aeruginosa lung infection compared with wild type (WT), single gene SP-A KO and SP-D KO mice. 23
The expression of SP-A and SP-D has been observed in the mucosal surface of the lung and several extra-pulmonary organs, including kidney.24–27 Mucosal epithelial cells and surfactant defense proteins form a physical barrier in the lung and urinary tract to prevent pathogens from entering the body. Decreased levels of urinary SP-A and SP-D were recently associated with recurrent UTIs in females. 28 We previously showed that SP-D functions as an innate immune factor and modulates inflammation in renal tubular epithelial cells in vitro. 26 Recently, it has been shown that SP-D protein could inhibit adherence of UPEC to bladder epithelial cells and the bacterium-induced cytotoxicity. 29 The current study examines the role of SP-A and SP-D in murine UTI and provides evidence that SP-A and SP-D function as mediators of innate immunity by inhibiting bacterial growth and modulating renal inflammation in part by regulating p38 MAPK-related pathways in murine UTI.
Materials and methods
Mice
Female mice (8–10 wk old) were used in this study. Original SP-A/D KO mice with C57BL/6 background were generated by Hawgood et al., 22 and SP-A/D KO mice were bred in the animal core facility at SUNY Upstate Medical University and maintained under pathogen-free conditions. Age-matched WT C57BL/6 mice were purchased from Jackson Laboratories (Bar Harbor, ME, USA). All animal experiments were conducted with an approved protocol (IACUC #253) in accordance with IACUC guidelines of SUNY Upstate Medical University and the National Institutes of Health guidelines on the use of laboratory animals.
Bacterial strain and mouse UTI model
UPEC (strain CFT073) was purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA). Serial passages of static growth of E. coli (CFT073) were made in lysogeny broth (LB) at 37℃, by which the expression of type 1 fimbriae was increased. Bacteria were harvested by centrifugation at 2000 g for 10 min at 4℃ and re-suspended in PBS buffer. The bacterial solution was adjusted to OD600 = 0.5 with PBS buffer. UTI was induced as previously described, 30 with some modifications. In brief, mice were anesthetized by i.p. injection with ketamine/xylazine (90 mg/kg ketamine and 10 mg/kg xylazine), and were gently massaged and pushed down on the bladder to expel urine. Then, bacterial solution (OD600 = 0.5, 50 μl/mouse) was delivered transurethrally using a sterile 0.28-mm inner diameter polyethylene catheter. Control mice underwent a sham procedure with administration of 50 μl of sterile PBS instead of bacterial suspension. In a pilot study the peak of bacterial load in the kidneys was found to be around 24 h after infection. Therefore, mice were sacrificed two time points—24 h or 48 h post-infection under anesthesia condition with intraperitoneal ketamine/xylazine. Tissue samples (kidneys) were excised and either immediately frozen in liquid nitrogen or placed in 10% neutral formalin for subsequent histological analysis. Sections were stained with hematoxylin and eosin in a standard fashion and assessed quantitatively the inflammatory score by two experienced investigators. Neutrophils in urine were quantified with countess automatic cell counter (Life Technologies, Grand Island, NY, USA) and were further confirmed using side-by-side comparisons with trypan blue straining in a hemocytometer, and with strained cytospin slides examined by light microscopy. Previous studies have shown that 99% of the infiltrated inflammatory cells were neutrophils. 31
RT-PCR
Total RNA was isolated from the kidney and lung of mouse using the RNA-Bee reagent (Tel-test, Friendswood, TX, USA) according to the manufacturer's instructions. cDNA was synthesized from 1 μg total RNA with oligo-dT primer using the superscript III First-strand synthesis system (Invitrogen, Carlsbad, CA, USA). PCR was performed with primers for SP-A (sense primer: GTGTGCGGGGATCTGAAGTTG; antisense primer: CCGGCTCTGGTACACATCTC), SP-D (sense primer: GCTGGGCCCAAAGGAGAAGTAGGT; antisense primer: TAACAAGGCGCTGCTCTCCACAAG), β-actin (sense primer: GGGAATGGGTCAGAAGGACT; antisense primer TTTGATGTCACGCACGATTT), respectively. To avoid any genomic DNA contamination in the PCR amplification the primers of each pair are located in the different exons of SP-A, or SP-D, or β-actin gene. The PCR products were examined by 1.2% agarose gel electrophoresis.
Histology and immunohistochemistry
The kidney was fixed in 10% neutral formalin and embedded in paraffin. Sections were cut and stained with hematoxylin and eosin. The inflammatory scores in the bladders and kidneys were determined in a blinded fashion using the histological grading criteria as previously described. 32 In brief, grade 0 = normal; 1 = focal inflammation; 2 = focal inflammation (more severe) from pelvis to medulla, with and without moderate edema; 3 = multifocal inflammation and cells from pelvis to medulla; 4 = extensive segmental inflammation and necrosis evident from pelvis to cortex; 5 = diffuse tissue necrosis and inflammatory cell infiltration extending from pelvis to cortex. Immunohistochemistry (IHC) was performed as per our previously protocol. 33 Sections were incubated with Abs against mouse SP-A (gift from Dr. McCormack, University of Cincinnati, OH, USA), mouse SP-D (gift from Dr. Wright, Duke University Medical Center, Durham, NC, USA) or neutrophil marker NIMP-R14 (1:200; Hycult Biotech, Uden, the Netherlands), overnight (14–16 h) at 4℃, followed by biotinylated secondary Ab and avidin–biotin peroxidase complex (Vector Laboratories, Burlingame, CA, USA). Peroxidase activity was visualized by DAB Kit (Vector Laboratories) and the sections were then counter-stained with hematoxylin. Negative controls were performed by nonimmune serum instead of first Ab. To quantitate filtrating neutrophils in the slides of each sample, neutrophils were counted in a total of 20 fields per sample (400×).
Determination of bacterial loads
Kidney tissue was homogenized with an Ultra Turrax T8 homogenizer (IKA Labortechnik, Staufen, Germany) in 0.5 ml sterile PBS. Homogenates were serially diluted in PBS, 100 μl aliquots of kidney homogenate or urine were plated onto LB agar plates, and the number of colonies in kidney or urine was determined after overnight culture at 37℃. CFU were enumerated using the Quantity One colony-counting software (Bio-Rad, Hercules, CA, USA). Quantitative cultures were expressed as CFU/ mg of kidney tissue or CFU/ml of urine.
ELISA
Samples of mouse kidneys were homogenized in RIPA buffer. The levels of keratinocyte-derived chemokine (KC; the functional homolog of human IL-8) and IL-17 in kidney homogenates were measured by KC Assay Kit (SABiosciences, Frederick, MD, USA) according to the manufacturer's instructions. Total protein content of the samples was determined by using BCA protein assay kit (Thermo Scientific, Rockford, IL, USA).
Western blotting analysis
Samples of mouse kidney and lung were homogenized in RIPA buffer containing a cocktail of protein inhibitors and phosphatase inhibitors (Roche Molecular Biochemicals, Indianapolis, IN, USA). Protein samples were subjected to electrophoresis with Tris-glycine gel and transferred to PVDF membranes. Membranes were probed with Abs against mouse SP-A, mouse SP-D, p38 MAPK (Santa Cruz Biotechnology, Santa Cruz, CA, USA), phosphorylated p38 MAPK (Santa Cruz Biotechnology) or β-actin (Santa Cruz Biotechnology). Bands were detected using ECL method (Thermo Scientific, Rockford, IL, USA).
Preparation of SP-A and SP-D proteins
Native human SP-A was purified from bronchoalveolar lavage fluid (BALF) as described previously. 34 Human SP-D was purified from stably transfected Chinese hamster ovary cell lines with the pEE14–hSP-D construct as described. 26 SP-D was purified from the conditioned culture medium using maltose-affinity chromatography according to the previous method. 35 The purity of the SP-A and SP-D preparation were verified by SDS-PAGE followed by silver staining. All proteins were filtered through a 0.2-micron filter to remove any potential contamination.
In vitro study of bacterial growth
For in vitro growth inhibition with SP-A and SP-D, UPEC CFT 073 was grown in 5 ml LB medium at 37℃ overnight. Bacterial cells were harvested by centrifugation at 2000 g for 10 min. Bacteria were re-suspended and diluted to a final OD at 600 nm of approximately 0.2 in sterile TBS (20 mM Tris/140 mM NaCl, 2.5 mM Ca++, pH7.4). 36 In dose-dependent experiments, 80 µl diluted CFT 073 suspensions was mixed with SP-A at concentrations ranging from 0 to 40 µg/ml of SP-A or SP-D at concentrations ranging from 0 to 20 µg/ml of SP-D, and then cultured for 5 h. In time-course experiments, diluted CFT073 solution was mixed with SP-A (final concentration at 40 µg/ml of SP-A) or SP-D (final concentration at 20 µg/ml of SP-D) and then cultured for analysis. Bacterial density was monitored by measuring at OD600 at 1-h intervals. BSA protein was used as negative control. 14 After 5 h of incubation, samples were diluted to 106 times with TBS and TBS plus 2 mM EDTA. Of the diluted suspensions, 100 μl was cultured on LB agar plate. Plates were incubated overnight at 37℃ and then colonies were counted.
Statistical analysis
Data were expressed as means ± SEM, and statistical analyses were performed using SigmaStat 3.5 (Jandel Scientific, San Rafael, CA, USA). Student's t-test or Mann–Whitney U-test was performed to assess the statistical significance of differences. A P-value < 0.05 was considered statistically significant.
Results
SP-A and SP-D expression in the kidney of WT mice
To examine the expression and distribution of SP-A and SP-D in mouse kidney, RT-PCR, IHC and Western blot analysis were performed. SP-A and SP-D mRNA and protein expression was detected in the kidney of WT mice but not SP-A/D KO mice (Figure 1). IHC showed that immunoreactivity for SP-A and SP-D is present in the proximal tubules and medullary collecting tubules of WT mice but not in the SP-A/D KO mice (Figure 1b). Furthermore, no significant difference of SP-A and SP-D expression before and after infection was observed in WT mice (Figure 1c).
SP-A and SP-D expression in the kidney of WT mice. Expression of SP-A and SP-D were analyzed in the kidney tissues of WT and SP-A/D KO mice by (a) RT-PCR, (b) IHC and (c) Western blot analysis. (a) SP-A mRNA (540 bp RT-PCR product, pointed by arrow) and SP-D mRNA (694 bp RT-PCR product, pointed by arrow) expression was detected in the kidney and lung (positive control) of WT mice but not in SP-A/D KO mice. (b) Positive staining of SP-A and SP-D expression was detected in the tubular epithelial cells (arrows) of WT kidney but not in SP-A/D KO mice. (c) SP-A and SP-D proteins were detected by Western blot analysis but no difference between infected and uninfected WT mice was found (n = 4–6 mice/group).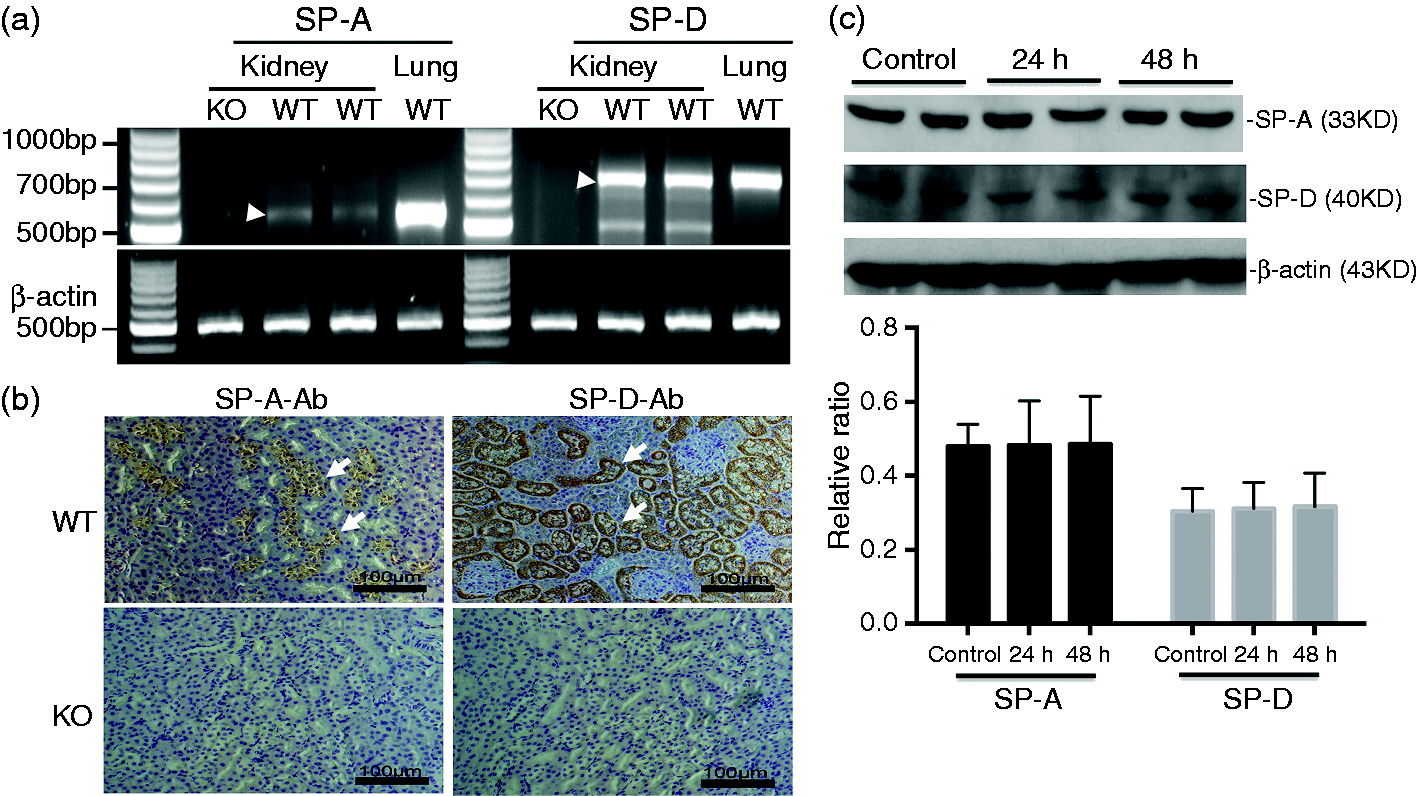
Increased susceptibility to UTI in SP-A/D KO mice
In order to study the effects of SP-A and SP-D in UTI, SP-A/D KO and WT female mice were infected with UPEC CFT073 by intravesical inoculation as previously described.
30
Renal and urine bacterial loads in the infected mice were determined 24 h and 48 h after infection. At both time points, the SP-A/D KO mice demonstrated increased CFU of E. coli in both the kidney and the urine compared with WT mice (Figure 2), suggesting that the ability of SP-A/D KO mice to remove and/or kill urinary E. coli is impaired relative to WT mice.
In vivo bacterial loads in WT and SP-A/D KO mice. CFUs in kidney or urine from infected SP-A/D KO and WT mice were determined by culturing overnight at 37℃. At 24 h and 48 h after inoculation, SP-A/D KO mice had significantly increased numbers of CFU in both (a) kidney and (b) urine when compared with WT mice (n = 8–12 mice/group).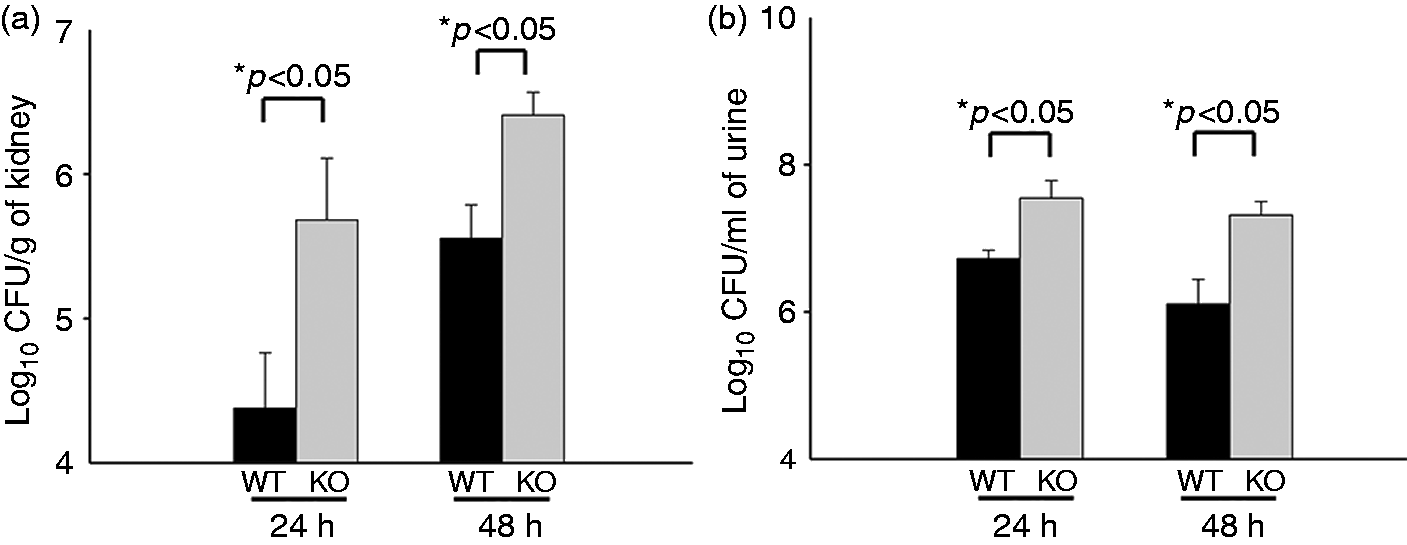
An essential aspect of the host response to invading bacteria is the recruitment of neutrophils to the site of infection. Therefore, renal tissues of the experimental mice were analyzed histologically for the presence of neutrophils. The kidneys of control SP-A/D KO and WT mice were histologically normal, showing no inflammation or other alterations. In contrast, infected WT and SP-A/D KO mice demonstrated inflammatory infiltrates in the medullary region in the kidney, consisting of almost exclusively neutrophils and monocytes (Figure 3). Kidneys of infected SP-A/D KO mice showed more neutrophil/moncyte infiltration and a few scattered microabscesses when compared with infected WT mice (Figure 3b). The inflammatory score was higher in the kidney of SP-A/D KO mice compared with WT mice 24 and 48 h after infection (Figure 3c).
Histological analysis of the kidney of infected WT and SP-A/D KO mice. Histological assessment of the kidney pyelum in (a) infected WT mouse and (b) infected SP-A/D KO mouse at 24 h after infection. Infected KO mice exhibited prominent inflammatory cell infiltration (arrows). (c) Kidney inflammatory score of SP-A/D KO mice was higher than that of WT mice 24 and 48 h after infection. Hematoxylin and eosin, bar = 150 µm (n = 4–6 mice/group).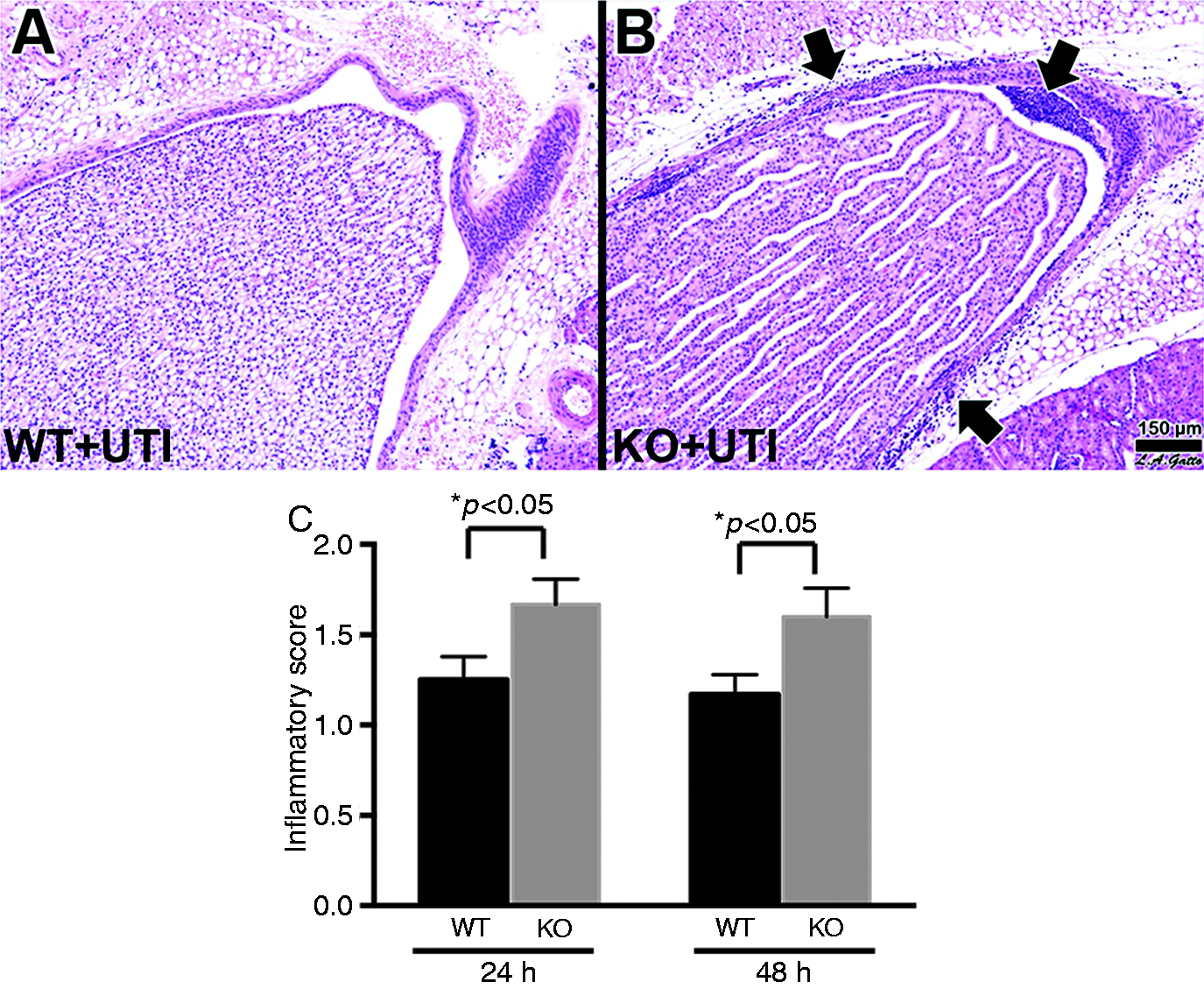
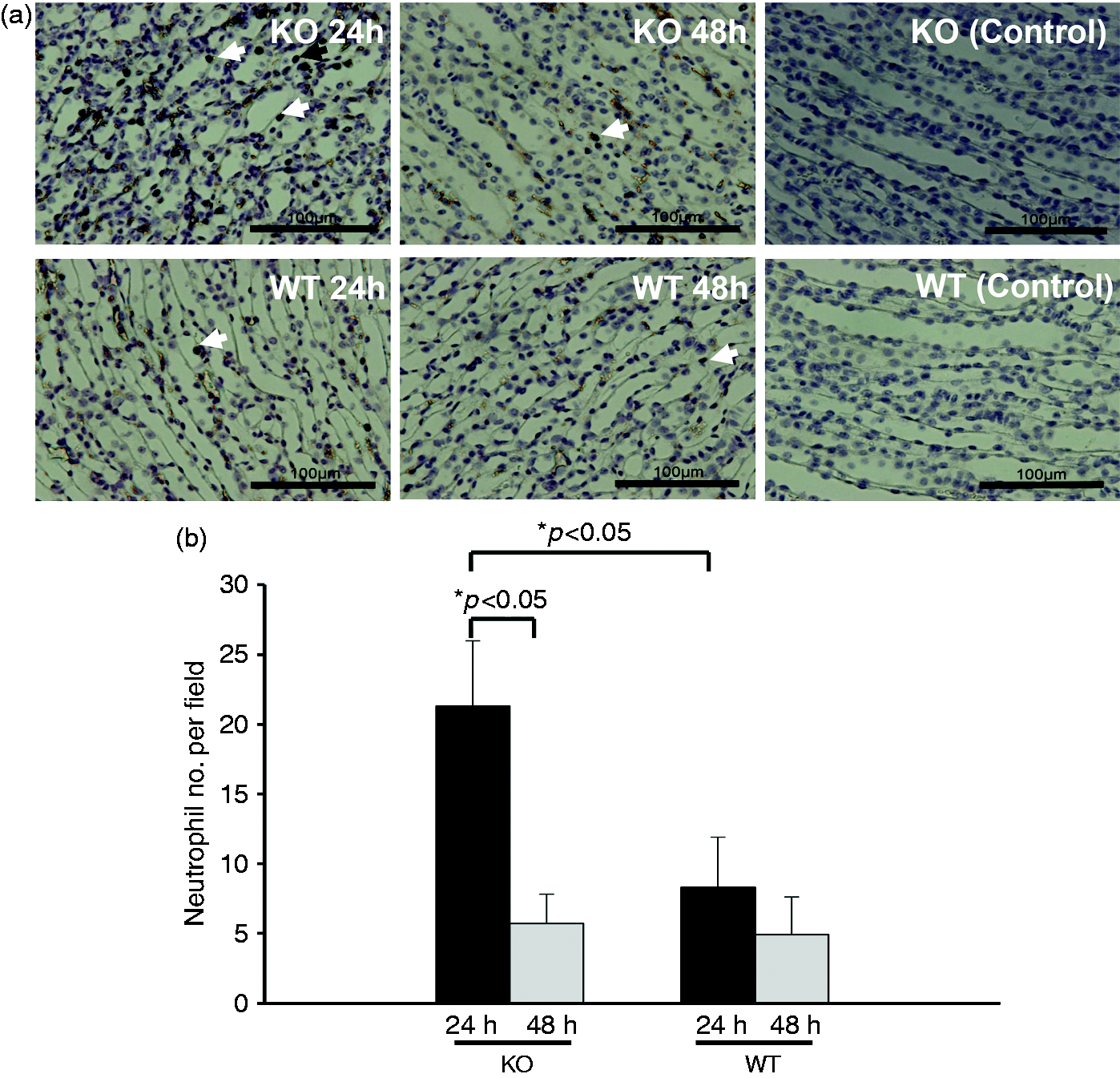
We further quantified the number of neutrophils in the urine collected from infected SP-A/D KO and WT mice. The results indicate that infected SP-A/D KO mice had more neutrophils in the urine 24 h and 48 h post infection than infected WT mice (Figure 4). We also examined the number of monocytes/macrophages in the urine. The results indicate that the number of monocytes/macrophages was < 1% neutrophils and there was no significant difference between infected SP-A/D KO and WT mice. Furthermore, neutrophil/monocyte infiltration in the kidneys of infected SP-A/D KO and WT mice was examined by IHC using anti-neutrophil-specific Ab NIMP-R14 (Hycult Biotech) (Figure 5). The results shown in Figure 5a indicate that infiltrating neutrophils and monocytes were scattered throughout the collecting tubules in the kidney from both infected SP-A/D KO and WT mice. However, the number of infiltrating neutrophils and monocytes in kidney tissues from infected SP-A/D KO mice is greater than observed in infected WT mice 24 h post infection (P < 0.05) (Figure 5b). Infiltrating neutrophils and monocytes decreased from 24 h to 48 h post-infection in SP-A/D KO mice (P < 0.05). On the basis of these observations, we conclude that SP-A/D KO mice are more susceptible to UTI than WT mice, suggesting that SP-A and SP-D are critical effectors of innate immunity in UTI.
Neutrophils in the urine of infected WT and SP-A/D KO mice. (a) Numbers of neutrophils in the urine were significantly increased in infected SP-A/D KO mice compared with WT mice at both 24 h and 48 h after infection (n = 5–8 mice/group). (b) A representative image of inflammatory cells (neutrophils appointed by arrows) from infected mouse urine. Neutrophils in the renal tissues of infected SP-A/D KO and WT mice. (a) Neutrophils were detected in kidney tissues by IHC using neutrophil-specific RIMP-R14 Ab. At 24 h after infection, increased neutrophils were observed in the infected SP-A/D KO mice compared with the infected WT mice. However, at 48 h after infection, a few neutrophils were detected in both the infected SP-A/D KO and infected WT mice. There was no positive staining in the negative control (nonimmune serum instead of first Ab). Bar = 100 µm. (b) The quantitative analysis of neutrophils indicate that numbers of neutrophils in the kidney were significantly increased in infected SP-A/D KO mice compared with infected WT mice at 24 h after infection; and numbers of neutrophils in the kidney of infected SP-A/D KO mice decreased from 24 h to 48 h post infection (n = 5–8 mice/group)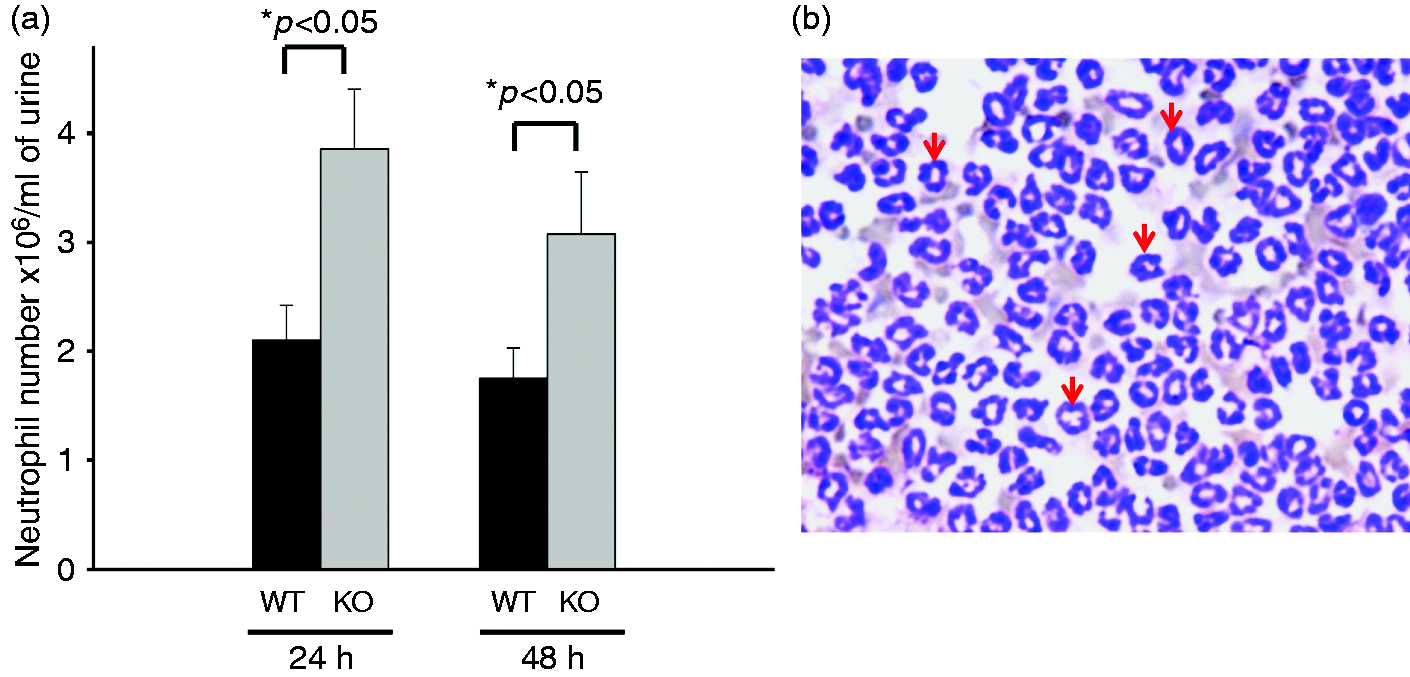
Differential KC and IL-17 expressions in infected SP-A/D KO and WT mice
KC (functional homolog of human cytokine IL-8) is an important inflammatory mediator in the pathogenesis of kidney infection. Both SP-A and SP-D have been shown to regulate inflammatory mediator expression in the infected tissues. We therefore examined KC expression in the kidney of infected SP-A/D KO and WT mice. Basal levels of KC in both WT and SP-A/D KO mice were comparable. KC levels in WT mice increased significantly 24 h after infection and decreased to near baseline level at 48 h (Figure 6a). However, in the SP-A/D KO mice KC level failed to rise after infection (Figure 6a). These results suggest that mice lacking SP-A and SP-D do not efficiently up-regulate the kidney KC expression in the murine UTI model. Furthermore, we examined IL-17 expression in this study. Compared with control, the level of IL-17 was elevated in the kidney of infected WT and SP-A/D KO mice (Figure 6b; P < 0.01); the infected SP-A/D KO mice had higher IL-17 expression than the infected WT mice 24 and 48 h post-infection (Figure 6b; P < 0.05), suggesting the immunoregulatory effects of SP-A and SP-D in the UTI.
Renal KC and IL-17 levels in the infected WT and SP-A/D KO mice. KC and IL-17 levels in the kidney tissues of WT and SP-A/D KO mice were measured by ELISA. The data of KC and IL-17 levels were expressed as pg/mg of total protein. Basal levels of KC and IL-17 were determined with uninfected mice. Panel A shows that the level of renal KC in SP-A/D KO mice showed no significant changes after infection, but the KC level of infected WT mice increased significantly at 24 h when compared with infected SP-A/D KO mice, or the basal level of WT mice. The KC level of infected WT mice decreased to near baseline at 48 h after infection. Panel B depicts IL-17 expression in the kidney of WT and SP-A/D KO mice. Compared with control, the level of IL-17 was elevated in the kidney of infected WT and SP-A/D KO mice (P < 0.01); and the infected SP-A/D KO mice had higher IL-17 expression than the infected WT mice 24 and 48 h post infection (P < 0.05) (n = 4–6 mice/group).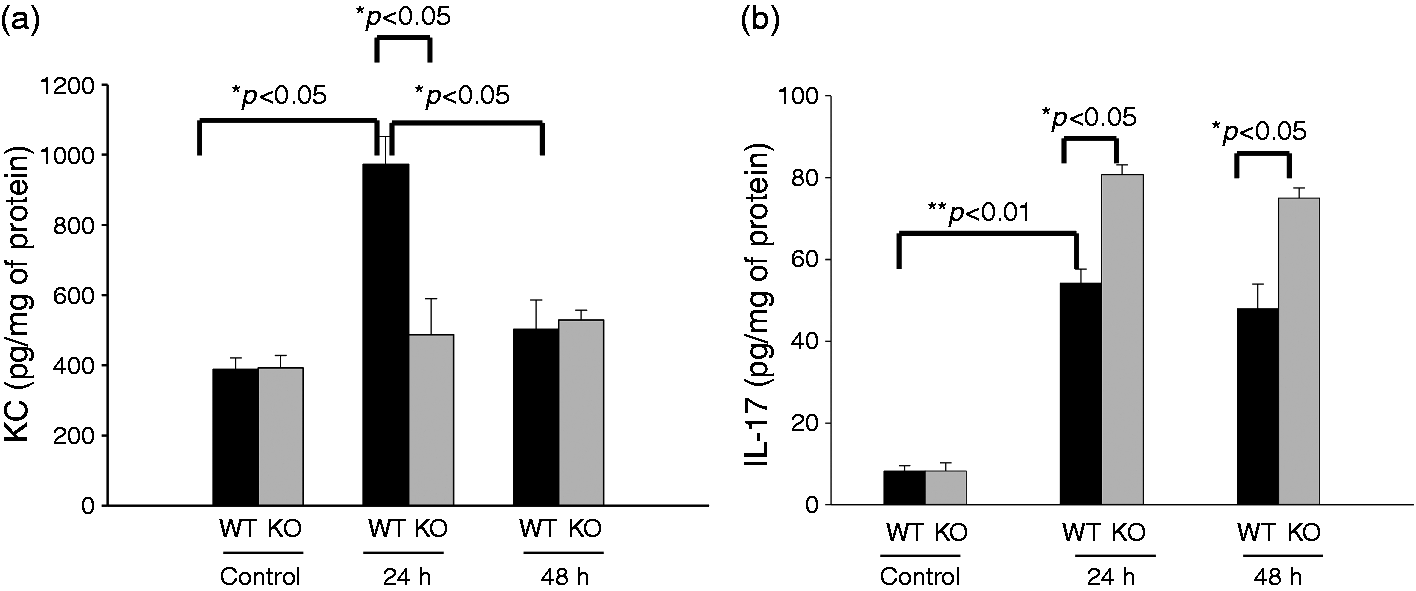
p38 MAPK phosphorylation in kidney tissue
Previous in vitro study has shown that SP-A and SP-D could regulate p38 MAPK phosphorylation and its downstream activation of the signaling pathway.
18
Therefore, we examined levels of p38 MAPK phosphorylation in the kidneys of SP-A/D KO mice and WT mice by Western blot with phosphorylated p38 MAPK-specific Ab. Interestingly, the basal level of p38 MAPK phosphorylation was significantly increased in kidney from healthy SP-A/D KO mice relative to WT control mice (Figure 7a). The increased level of phosphorylated p38 MAPK was observed in the kidney of infected WT mice 24 and 48 h after infection (Figure 7b); however, the level of phosphorylated p38 MAPK decreased in the infected SP-A/D KO mice 24 h after infection compared with uninfected mice (control), and then increased to a similar level as the control 48 h after infection (Figure 7c).
Level of p38 MAPK phosphorylation in the kidney. Total protein (20 μg) of kidney tissue was subjected to electrophoresis and then transferred onto a PVDF membrane. Phosphorylated p38 MAPK (p-p38 MAPK) and p38 MAPK (un-phosphoryated) were detected with specific p-p38 MAPK Ab and un-phosphorylated p38 MAPK Ab, respectively. (a) Relative level of p-p38 MAPK was normalized to un-phosphorylated p38 MAPK. Higher p38 MAPK phosphorylation level in the kidney of SP-A/D KO mice was observed compared with WT mice. (b) The level of p-p38 MAPK increased 24 and 48 h post infection in WT mice; however, (C) the level of p-p38 MAPK decreased in infected SP-A/D KO mice 24 h post infection and then increased to similar level as control 48 h post infection. (n = 4–6 mice/group)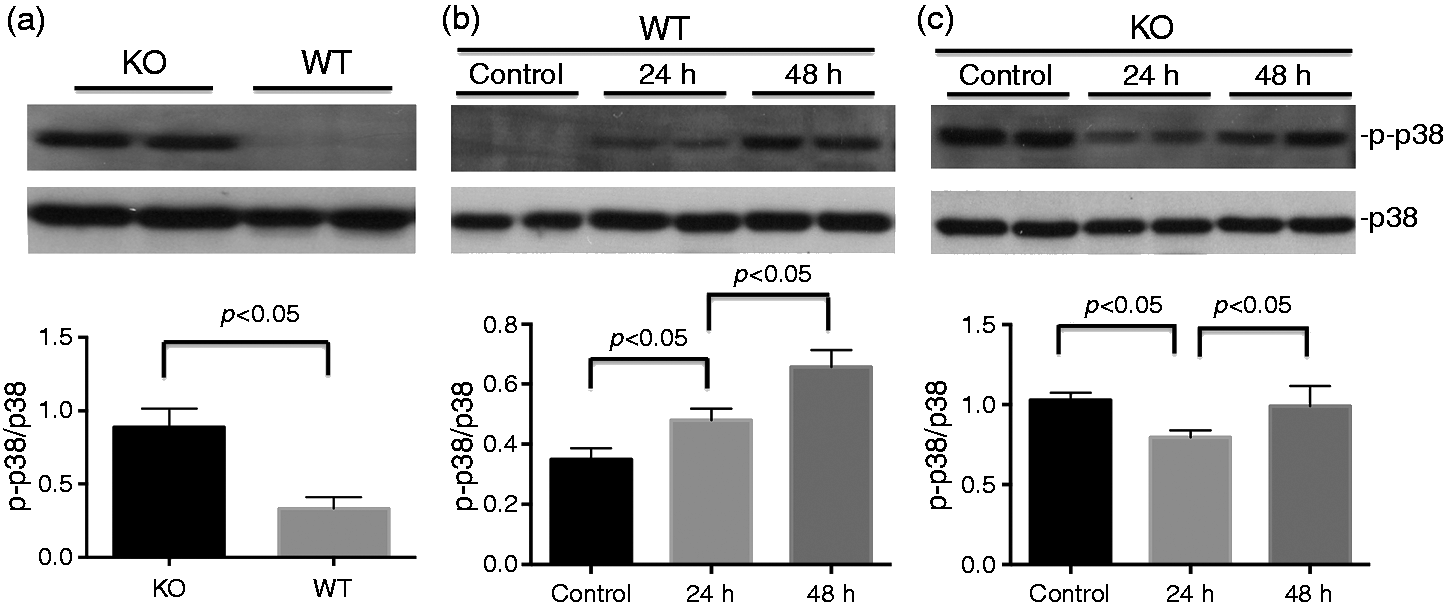
Effects of SP-A and SP-D on the inhibition of in vitro growth of UPEC
The effects of SP-A and SP-D on in vitro growth of E. coli were measured by monitoring the OD (OD600) of cultured bacteria in TBS buffer containing 2.5 mM Ca++, with or without SP-A or SP-D, and final CFU of the culture were quantified. In the absence of SP-A or SP-D, E. coli growth and the OD600 increased steadily from 1 to 5 h cultivation. The results from dose–course experiments indicated that 40 µg/ml SP-A or 20 µg/ml SP-D could significantly inhibit the growth of UPEC (Figure 8a, b). Therefore, these concentrations of SP-A and SP-D were used in the following experiments. The results indicated that, in the presence of SP-A (40 µg/ml) or SP-D (20 µg/ml), bacterial growth was markedly inhibited from 2 to 5 h for SP-A (P < 0.05) (Figure 8c) and from 3 to 5 h for SP-D (P < 0.05) compared with only BSA (Figure 8d). To further examine the effects of SP-A and SP-D on bacterial viability, the final CFU of the culture were determined by colony-count assays. The results showed that E. coli treated with SP-A (40 µg/ml) (Figure 8e) or SP-D (20 µg/ml) (Figure 8f) resulted in significantly lower CFUs (P < 0.05) compared with the BSA controls. Additionally, to eliminate the effect of BSA on the growth of the bacteria, we also cultured bacteria in TBS buffer without BSA as control; the results of the bacterial growth were similar to those with BSA. These results suggest that both SP-A and SP-D may inhibit the growth of UPEC, or are bactericidal under these experimental conditions.
Effects of SP-A and SP-D on the UPEC growth inhibition in vitro. For dose-dependent experiments, UPEC (strain CFT 073) bacterial solutions in TBS were added with (a) SP-A at concentrations ranging from 0 to 40 µg/ml or of (b) SP-D at concentrations ranging from 0 to 20 µg/ml, and then were incubated at 37℃ for 5 h. For time-course experiments, bacterial solutions in TBS were added with (c) 40 µg/ml of SP-A or (d) 20 µg/ml of SP-D, or BSA (control) and then incubated for 5 h. Bacterial growth was monitored by measuring OD at 600 nm. Relative OD600 value was used to represent bacterial growth. CFU of the bacterial culture were determined after mixing SP-A or SP-D for 5 h. The results showed bacterial growth was inhibited threefold by (E) 40 µg/ml of SP-A and twofold by (F) 20 µg/ml of SP-D (n = three independent experiments). *P < 0.05.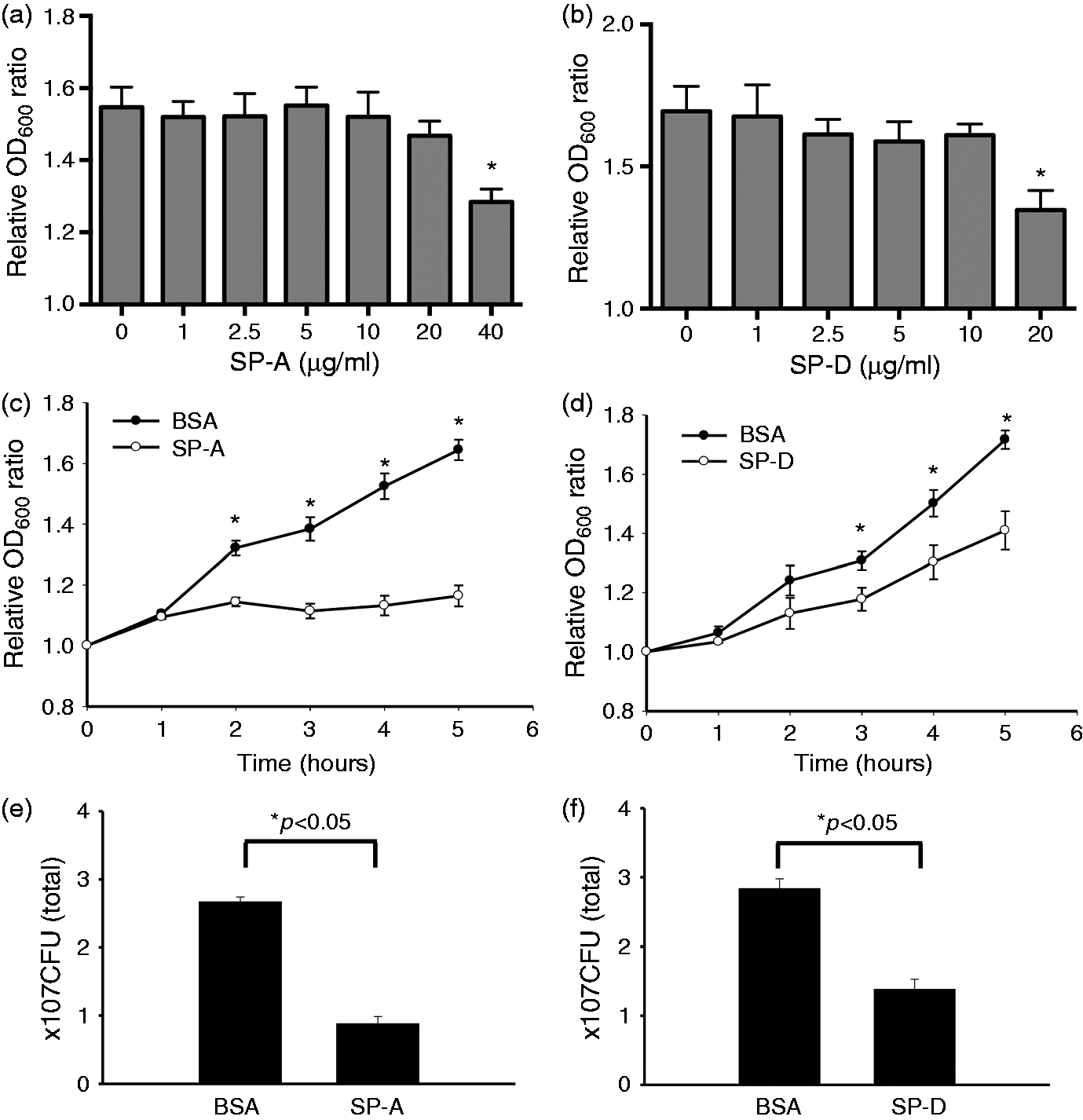
Discussion
UTI is one of the most common infections in women. Numerous host factors have been implicated in the defense mechanisms against UTI induced by UPEC.6,37 In a previous study, we found decreased levels of SP-A and SP-D in urine from female patients with recurrent UTI, 28 suggesting that SP-A and SP-D, two important innate immune proteins, might be involved in the host defense against UTI. To better understand the mechanisms underlying the role of SP-A and SP-D in UTI, SP-A/D KO and age-matched female WT mice were studied in a murine model of UPEC-induced UTI. This is the first study to investigate the roles of SP-A and SP-D in an experimental model of UTI. Our results provide evidence that SP-A/D KO mice are more susceptible to UPEC infection, showing increased bacterial loads and more infiltrating neutrophils in the kidney, the bladder and the urine, as well as severe histopathological changes with higher inflammatory score in the kidney, when compared with WT mice. Of interest, we found increased basal levels of p38 MAPK phosphorylation in SP-A/D KO mice but lack response to infection in KC expression in the SP-A/D KO mice. Furthermore, UPEC growth was inhibited by SP-A and SP-D in vitro. These observations suggest that SP-A and SP-D exerted a direct antibacterial effect and modulated inflammatory response to UTI.
SP-A and SP-D were originally identified as surfactant-associated proteins, dominantly expressed in lung epithelial cells. 11 SP-A and SP-D expression was subsequently observed in several extra-pulmonary organs, including kidney, salivary gland, prostate gland, pancreas, intestine, stomach and female reproductive tract.24,25,27,38,39 Several studies demonstrated that both SP-A and SP-D proteins play roles not only in the respiratory system, 13 but also in the gastrointestinal tract and the reproductive system.40,41 The urinary tract system is exposed to various environmental factors including pathogens and requires multiple host defense mechanisms against environmental insults, including agents of the innate immunity such as SP-A and SP-D.
The immunoregulatory roles of SP-A and SP-D have recently received more attention.13,20 SP-A/D KO mice were shown to be more susceptible to lung infection with P. aeruginosa, when compared with single gene KO, e.g. SP-A KO, or SP-D KO, and WT mice. 23 Although SP-A and SP-D have shown the ability to inhibit the growth of nonpathogenic E. coli K-12, 14 it was unclear whether they would have the same effects on the growth of pathogenic E. coli as pathogenic strains usually have different composition/structure of endotoxin (LPS) from nonpathogenic strains. SP-A and SP-D can bind directly to rough LPS but not to smooth LPS from E. coli and other Gram-negative bacteria.14,42,43 Recent report indicates that SP-D has the effects on the inhibition of UPEC adherence to bladder epithelial cells and the bacterium-induced cytotoxicity. 29 This study demonstrated that both SP-A and SP-D could inhibit the growth of UPEC in vitro, but at relative higher concentrations than the nonpathogenic E. coli strain, 14 which may reflect difference between pathogenic and nonpathogenic strains. More effective agglutination of nonpathogenic E. coli K-12 mediated by SP-A and SP-D was observed compared with pathogenic E. coli CFT073 in this study (data not shown). Purified SP-A and SP-D proteins from the BALF of patients with alveolar proteinosis or recombinant mammalian cell expression systems have been widely used for functional studies in vitro and in vivo.14,42,44 The biological roles of SP-A and SP-D on the inhibition of bacterial growth and adherence serve as an effective innate immune mechanism against UTI. The notion is further confirmed by the results from the in vivo UTI, where SP-A/D KO mice, compared with WT mice, had higher bacterial loads in the infected kidney and the urine, indicating their decreased ability to remove E. coli in this UTI model.
The p38 MAPK signaling pathway plays a central role in the regulation of pro-inflammatory cytokine and chemokine production, because p38 MAPK activation can regulate several downstream pathways involved in pro-inflammatory cytokine expression. 45 Our previous study also found SP-A/D KO mice had higher basal level of autophagy in liver and a more pronounced inflammatory response to sepsis than WT mice.46,47 Of interest, in the present study, SP-A/D KO mice exhibited significantly higher basal levels of p38 MAPK phosphorylation compared with uninfected WT mice. Previous studies have shown that p38 MAPK is phosphorylated in response to bacterial infection and regulates the activation of nuclear transcription factors, including NF-κB, in UTI.48,49 Based on the observations from this study and other published information,18,44,49 SP-A and SP-D are two important mediators of innate immunity in the pathogenesis of UTI. SP-A and SP-D proteins can bind to E. coli and epithelial cell receptors, 50 like TLR2 and TLR4, through their CRD domain. When the urinary tract is infected, the CRD domain of SP-A and SP-D first binds to bacteria (UPEC), and thus the collagen-like region of SP-A and SP-D interacts with CD91/calreticulin receptor in the epithelial cells of the kidney, 51 by which the p38 MAPK signaling pathway could be activated and the expression of inflammatory mediators could be upregulated. In contrast, the level of p38 MAPK phosphorylation decreased 24 h post infection and the level of KC expression showed no response to UTI owing to the lack of SP-A and SP-D expression in the SP-A/D KO mice. This model also explains, at least in part, the increased NF-κB activity observed in lung tissue from uninfected SP-D KO mice. 52
Neutrophils are the most abundant and rapid cellular responders to the infected urinary tract. 53 Neutrophils also represent a major cause of renal scarring, and are implicated in the antibacterial defense of the urinary tract system. 53 In the present UTI model, more bacteria and neutrophil infiltration were observed in kidney and urine of the infected SP-A/D KO mice than those in the infected WT mice. One possible mechanism is the lack of the effects of SP-A and SP-D host defense. As an unexpected finding, no cytokine KC response but higher IL-17 expression to bacterial infection at 24 and 48 h in the infected SP-A/D KO mice were observed in this study, it is possible that IL-17 and other cytokines or chemokines, like MIP-2, MCP-1, GM-CSF and CCL5, may replace the effects of KC in the recruitment of neutrophils in the UTI model. For example, increased IL-17 and GM-CSF levels were found in uninfected SP-A/D KO mice and in UPEC-induced UTI animals,54,55 which may have an impact on the recruitment of neutrophils to the infectious site. Future studies are needed to explore the mechanisms of p38 MAPK activation response to UTI and to evaluate comprehensively cytokine and chemokine expression in SP-A/D KO mice. Epithelial cells of the upper urinary tract not only form a barrier to prevent pathogens, but also produce a variety of cytokines and chemokines in response to bacterial stimulation. 56 Murine KC is a functional homolog of human cytokine IL-8. IL-8 expression has been observed to be significantly increased in the bladder and kidney of human patients with UTI. 57 Human IL-8 production in the urinary tract epithelial cells is dependent on the p38 MAPK-mediated NF-κB activation. 58 The lack of KC response to E. coli infection in SP-A/D KO mice may be caused by the absence of interactions between SP-A, SP-D and cell receptors, as well as the activation of the p38 MAPK signaling pathway. Previous studies found SP-A and SP-D can interact with the various receptors, resulting in increased phosphorylation of p38 MAPK and up-regulation of the NF-κB pathway with enhanced expression of inflammatory factors in the infected lung.18,44,59 In the present study, the phosphorylation of p38 MAPK in response to bacterial infection was observed in WT mice but not SP-A/D KO mice. The process of p38 MAPK phosphorylation is a rapid process in the cells after stimulation/infection.49,58 Changes in abundance or activity of p38 activation or deactivation by phosphatases (phosphorylation/dephosphorylation) represent potential mechanisms. In addition, monocytes are known to arrive at the sites of infection in UTI model. 60 Monocytes can further differentiate into macrophages and clean up dying neutrophils. Both SP-A and SP-D can interact with receptors on the surface of macrophages and promote the clearance of dying cells. 13 SP-A and SP-D also play roles in the regulation of inflammatory cells and removal of dying cells in the UTI. However, compared with neutrophils, only a few monocytes and macrophages were determined in the urea if infected mice in the present study.
In summary, our results suggest that the protective effects of SP-A and SP-D in UTI are potentially mediated by two different mechanisms: (1) direct inhibition of UPEC growth and (2) SP-A and SP-D modulate inflammatory processes through regulation of p38 MAPK phosphorylation and its downstream signaling pathways (e.g. NF-κB activation). Collectively, our results provide evidence SP-A and SP-D function as critically important innate immune molecules in the urinary tract. Additional studies are needed to further elucidate the mechanisms by which SP-A and SP-D affect bacterial growth and aggregation of UPEC and tissue inflammatory responses in UTI, as well as the correlation between human SP-A and SP-D genetic variants and individual susceptibility to UTI using SP-A or SP-D single KO, and humanized transgenic mice.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by NIH grant HL096007 and grants from the National Science Foundation of China (30670985, 81070556, 81300617).
Acknowledgments
We would like to thank Dr. Francis X. McCormack of the University of Cincinnati for kindly providing human BAL fluid from AP patients and mouse SP-A Ab, Dr. Erika Crouch of the School of Medicine of Washington University in St. Louis for detailed protocol of SP-D purification, and Dr. Jo Rae Wright of the Duke University Medical Center for mouse SP-D Ab.