
Abstract
To test the dynamic changes of the expression of genes and microRNA in the gastrocnemius muscle after LPS challenge, 36 piglets were assigned to a control group (slaughtered 0 h after saline injection) and LPS groups (slaughtered at 1 h, 2 h, 4 h, 8 h, and 12 h after LPS treatment, respectively). After LPS treatment, the mRNA expression of IL-1β, IL-6, and TNF-α reached maximal levels at 1 h, 2 h, and 1 h, respectively (P < 0.05), and mRNA expression of TLR4, NODs, muscle-specific ring finger 1, and muscle atrophy F-box peaked at 12 h (P < 0.05). Moreover, the expression of miR-122, miR-135a, and miR-370 reduced at 1 h, 1 h, and 2 h, respectively (P < 0.05), and miR-34a, miR-224, miR-132, and miR-145 reached maximum expression levels at 1 h, 1 h, 2 h, and 4 h, respectively (P < 0.05). These results suggested that mRNA expression of pro-inflammatory cytokines was elevated in the early stage, mRNA expression of genes related to TLR4 and NODs signaling pathways and protein degradation increased in the later phase, and the expression of microRNA related to muscle inflammation and protein degradation changed in the early stage after LPS injection.
Introduction
Skeletal muscle is not only the most widely distributed tissue in the body, it is an important site for regulating the metabolism of the whole body. However, inflammation can cause muscle wasting, which is characterized by accelerated protein degradation. 1 Many systemic diseases such as cancer are accompanied by muscle atrophy 2 , 3 and might result from the inflammation. 4
TLRs play an essential role in the innate immune system and TLR4 is the central component of the sole mammalian LPS sensor. 5 LPS can stimulate TLR4 to initiate MyD88, followed by activation of IL-1 receptor associated kinase (IRAK) 1, which finally leads to NF-κB activation. Similarly, NLR 1 and 2 play an important role in the development of the innate immune response. Receptor-interacting protein kinase (RIPK) 2 is an essential downstream adaptor protein in the NOD signaling pathway, which can result in an NF-κB-mediated pro-inflammatory response. 6 NF-κB can induce the transcription of cytokines, including IL-1β, IL-6, and TNF-α. These pro-inflammatory cytokines can directly induce muscle protein degradation, or lead to muscle atrophy by up-regulating the mRNA expression of forkhead box O1 (FOXO1) and two muscle-specific E3 ligases, muscle atrophy F-box (MAFbx) and muscle-specific ring finger (MuRF) 1, 7 , 8 which is the major pathway of protein degradation in skeletal muscle. 1
MicroRNAs (miRNAs) are a recently discovered class of small non-coding RNAs, which play a distinct role in inflammation. Many mammalian miRNAs can regulate immune and inflammatory responses. 9 , 10 In addition, miRNAs can be a new therapeutic frontier for muscle wasting in chronic kidney disease. 11
We have found that the levels of pro-inflammatory cytokines in plasma and mRNA expression of genes related to inflammation and protein degradation in muscles increased in weaned pigs 4 h after LPS challenge. 12 , 13 Previous studies have reported that innate immune cells could be in diverse states with different degrees of pro- and anti-inflammatory phenotypes, 14 , 15 accompanied by varying consequences for host defense and inflammation. 16 Although LPS has been used for decades for the induction of some pro-inflammatory cytokines, the response and kinetics of inflammation varied extensively depending on the LPS timing. Therefore, this study was conducted to test the expression of selected pro-inflammatory mediators and miRNAs in TLR4, NODs, and FOXO/MAFbx-MuRF1 signaling pathways in muscle at different time points after LPS challenge in weaning piglets. Our study might reveal a dynamic change mechanism for the inflammation and protein degradation in the muscle of weaning piglets challenged by LPS.
Materials and methods
Experimental design
The Animal Care and Use Committee of Wuhan Polytechnic University approved the animal use protocol for this research. A total of 36 crossbred castrated barrows (Duroc × Large White × Landrace, 7.02 ± 0.21 kg, 28 ± 3 d) were selected and assigned randomly to six groups (n = 6) including control (saline injection) and LPS (Escherichia coli serotype 055: B5; Sigma Chemical, St. Louis, MO) groups in which the piglets were slaughtered at five different times after LPS challenge. Each group had six replicated pens, and each pen had one pig. 13 , 17 Diets were formulated to meet or exceed National Research Council (NRC) requirements for all nutrients. 18 All piglets were kept under a 12 h light/dark cycle, allowed ad libitum access to water and feed, and placed individually in pens (1.80 × 1.10 m) during a 14-d adaption period. The ambient temperature was maintained at 22–25°C and the living environment was in accordance with animal welfare guidelines. Signs of diarrhea, sickness, and abnormal behavior were not observed throughout the adaption period.
Muscle sample collection
On d 15, the piglets were injected intraperitoneally with either E. coli LPS at 100 μg/kg body mass (BM) or the equivalent amount of sterile saline (0.9%) according to our previous study. 13 Then the piglets in the LPS groups were slaughtered under anesthesia with an intramuscular injection of sodium pentobarbital (80 mg/kg BM) at 1 h, 2 h, 4 h, 8 h, and 12 h after LPS injection. The piglets in the control group were slaughtered in the same way at 0 h after saline injection. Then the gastrocnemius muscle was removed under the semitendinosus and biceps flexor cruris (left hind leg), frozen in liquid nitrogen immediately, then stored at −80°C for further analysis. Previous studies found that piglets with muscle fiber atrophy had high levels of MAFbx in the gastrocnemius muscle, therefore this muscle was selected to study muscle atrophy. 19 , 20
Muscle mRNA expression analysis
The gene expression was conducted by real-time PCR according to the previous study. 17 Briefly, total RNA was isolated using Trizol reagent (#108, TaKaRa Biotechnology (Dalian) Co., Ltd., Dalian, China), and cDNA was synthesized by a PrimeScript® RT reagent kit (#RR047A, TaKaRa Biotechnology (Dalian) Co., Ltd., Dalian, China) according to the manufacturer’s instructions. Quantitative results of the mRNA expression for the target genes were carried out using a SYBR® Premix Ex TaqTM (Tli RNaseH Plus) quantitative PCR kit (#RR420A, TaKaRa Biotechnology (Dalian) Co., Ltd., Dalian, China). The PCR cycling conditions were 95°C × 30 s, followed by 40 cycles of 95°C × 5 s and 60°C × 34 s. The primer sequences are shown in Table 1, and the 2−△△CT method was used to calculate the mRNA expression of the target genes relative to GAPDH. 21
Primer sequences used for real-time PCR.

RIPK2, receptor-interacting serine/threonine-protein kinase 2; FOXO1: forkhead box O 1; FOXO4: forkhead box O 4; MAFbx: muscle atrophy F-box; MuRF1: muscle-specific ring finger 1.
Muscle miRNA expression analysis
Total RNA was isolated by the Trizol reagent (#9108, TaKaRa Biotechnology (Dalian) Co., Ltd., Dalian, China) according to the manufacturer’s protocol. Mature miRNAs were detected and quantified using primers obtained from the mirVana™ quantitative RT-PCR Primer Set (Gene Pharma, Shanghai, China) and SYBR Green PCR Mix (Takara, Japan). The miRNA expression was normalized to the level of U6 and analyzed by the 2−△△CT method. 21
Statistical analysis
The experimental data were analyzed by ANOVA with SPSS 17.0 software. Differences among treatments were determined by Duncan’s multiple range tests. All data were expressed as means ± SE. The statistical significance level for all analyses was set at P < 0.05. A one-way multivariate ANOVA (MANOVA) was used to determine the effect size (partial eta squared) and the statistical power of the model. As for the effect of LPS challenge on the TLR4 and NOD signaling pathways, a one-way MANOVA revealed a significant multivariate main effect for region, partial eta squared was 0.884, and power to detect the effect was 1.000. For the effect of LPS challenge on the FOXO/MAFbx-MuRF1 signaling pathway, one-way MANOVA revealed a significant multivariate main effect for region, partial eta squared was 0.628, and power to detect the effect was 1.000. For the effect of LPS challenge on the miRNA expression, a one-way MANOVA revealed a significant multivariate main effect for region, partial eta squared was 0.766, and power to detect the effect was 1.000.
Results
The mRNA expression of pro-inflammatory cytokines in muscle
As shown in Figure 1, IL-1β mRNA expression reached the maximal level at 1 h (P < 0.05) and declined to the basal level at 2 h after LPS injection. Maximum IL-6 mRNA expression was observed at 2 h (P < 0.05) and declined to the basal level at 4 h after LPS challenge. TNF-α mRNA expression increased and reached the maximal level 1 h after LPS injection (P < 0.05).
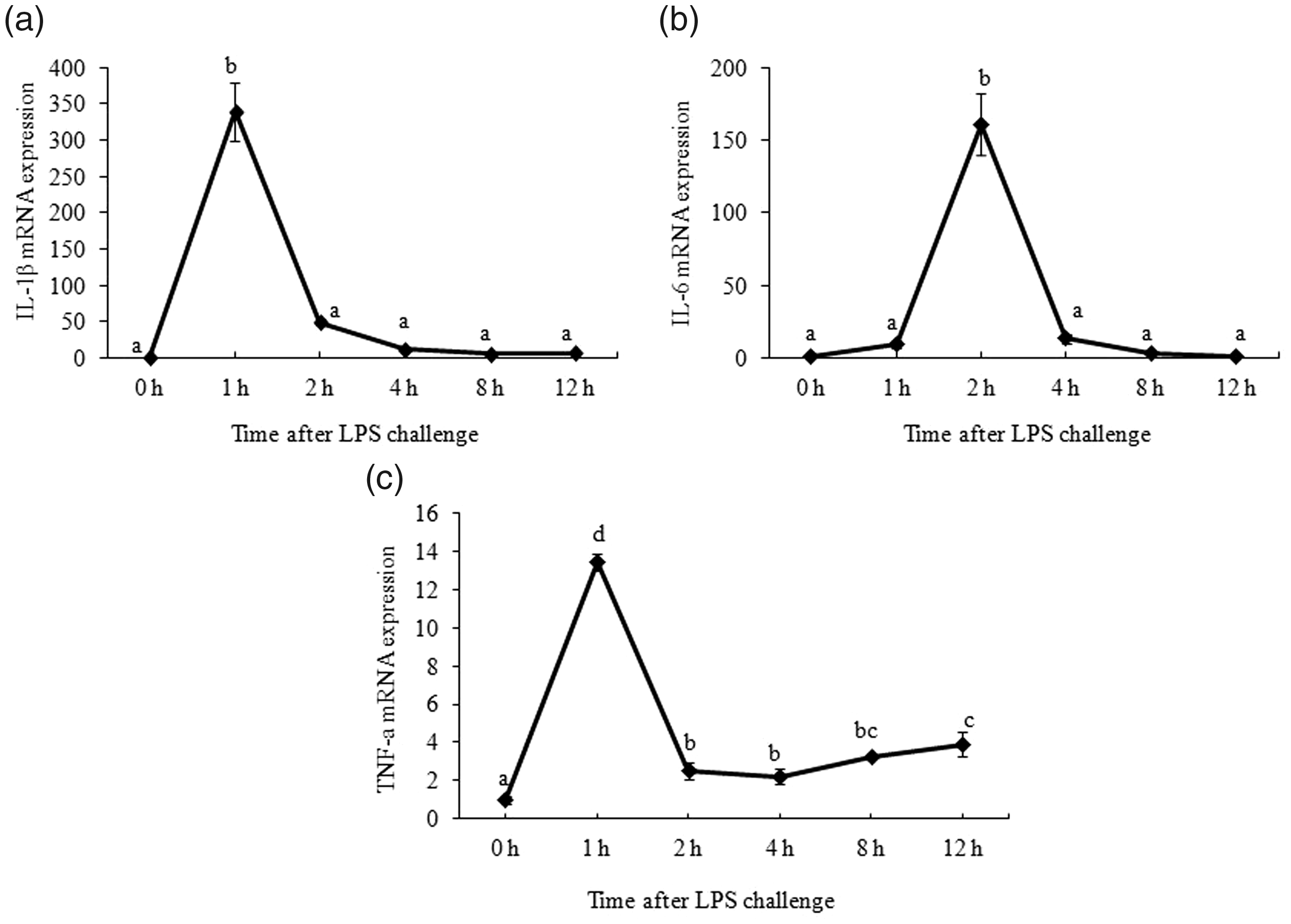
Effect of LPS on mRNA abundance of pro-inflammatory cytokines in muscle. Piglets were injected with 100 g/kg body mass (BM) or the equivalent amount of sterile saline. The piglets in the control group were slaughtered 0 h after saline injection. The piglets in LPS treatments were slaughtered at 1 h, 2 h, 4 h, 8 h, and 12 h after LPS injection. Values are single-point determinations and represent each of the time points listed above.
The mRNA expression of key genes in TLR4- and NOD-signaling pathways in muscle
As shown in Figure 2, LPS treatment elevated TLR4 mRNA expression at 4 h after LPS injection and peaked at 12 h after LPS treatment (P < 0.05). MyD88 mRNA expression increased at 2 h after LPS injection and reached the maximum level at 12 h after LPS treatment (P < 0.05). mRNA level of IRAK1 increased 1 h after LPS challenge and peaked at 8–12 h after LPS treatment (P < 0.05). In addition, NF-κB mRNA was elevated at 1 h (P < 0.05) and reached the maximum level at 8 h after LPS treatment (P < 0.05). Moreover, NOD1 mRNA expression started to increase at 8 h after LPS challenge (P < 0.05), and NOD2 mRNA expression started to increase at 1 h after LPS challenge (P < 0.05). The maximum mRNA expression levels of both NOD1 and NOD2 were detected at 12 h after LPS injection (P < 0.05). RIPK2 mRNA expression reached the maximum at 1 h (P < 0.05) and dropped to the basal level at 8 h after LPS challenge.
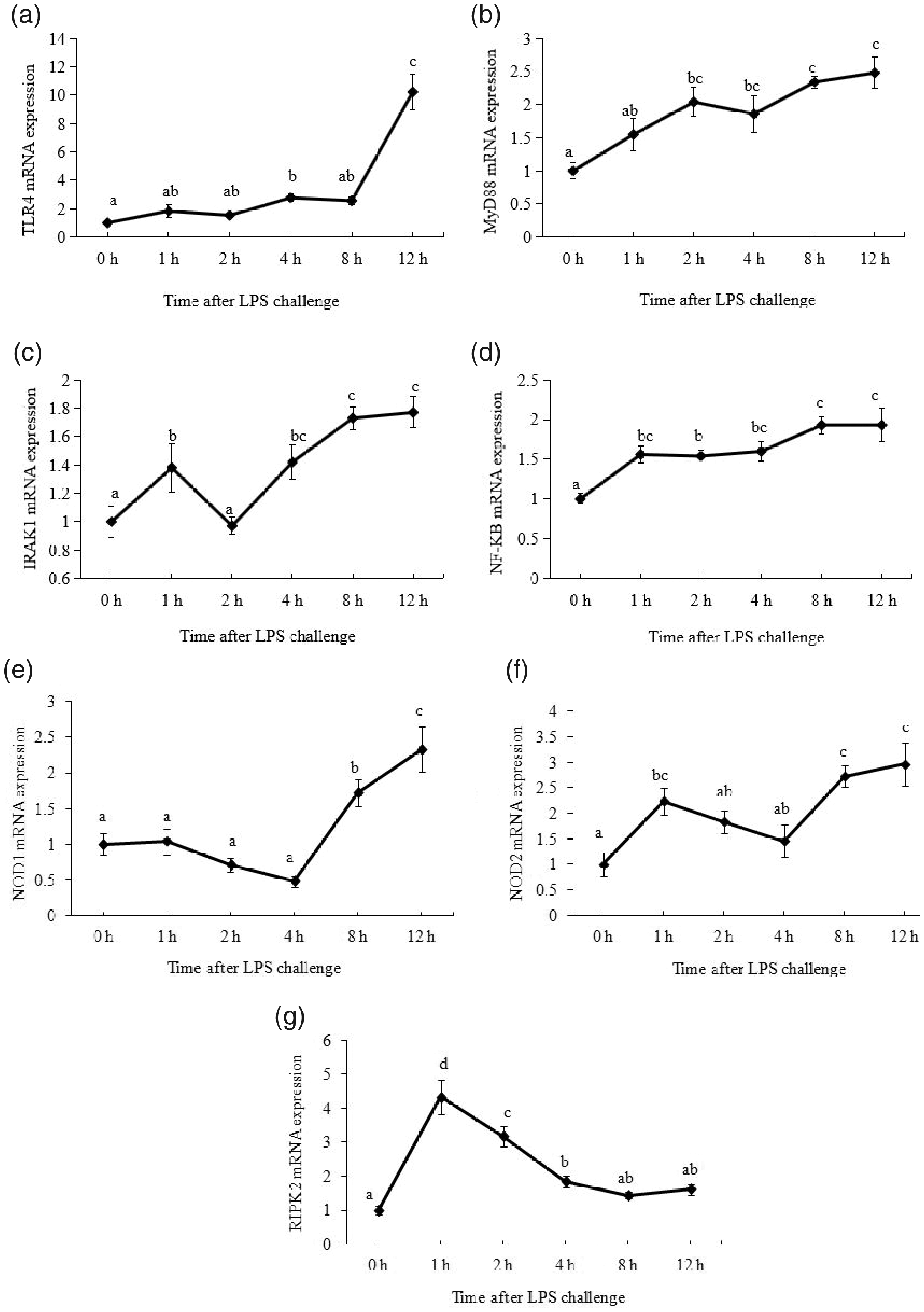
Effect of LPS on mRNA abundance of key genes in TLR4 and NOD signaling pathway in muscle. Piglets were injected with 100 g/kg body mass (BM) or the equivalent amount of sterile saline. The piglets in the control group were slaughtered 0 h after saline injection. The piglets in LPS treatments were slaughtered at 1 h, 2 h, 4 h, 8 h, and 12 h after LPS injection. Values are single-point determinations and represent each of the time points listed above.
The mRNA expression of key genes in FOXO/MAFbx-MuRF1 signaling pathway in muscle
As shown in Figure 3, maximum FOXO1 and FOXO4 mRNA expression was detected at 8 h after LPS challenge (P < 0.05). MuRF1 and MAFbx mRNA expression started to increase at 4 h, and reached the maximum levels at 12 h, respectively (P < 0.05).
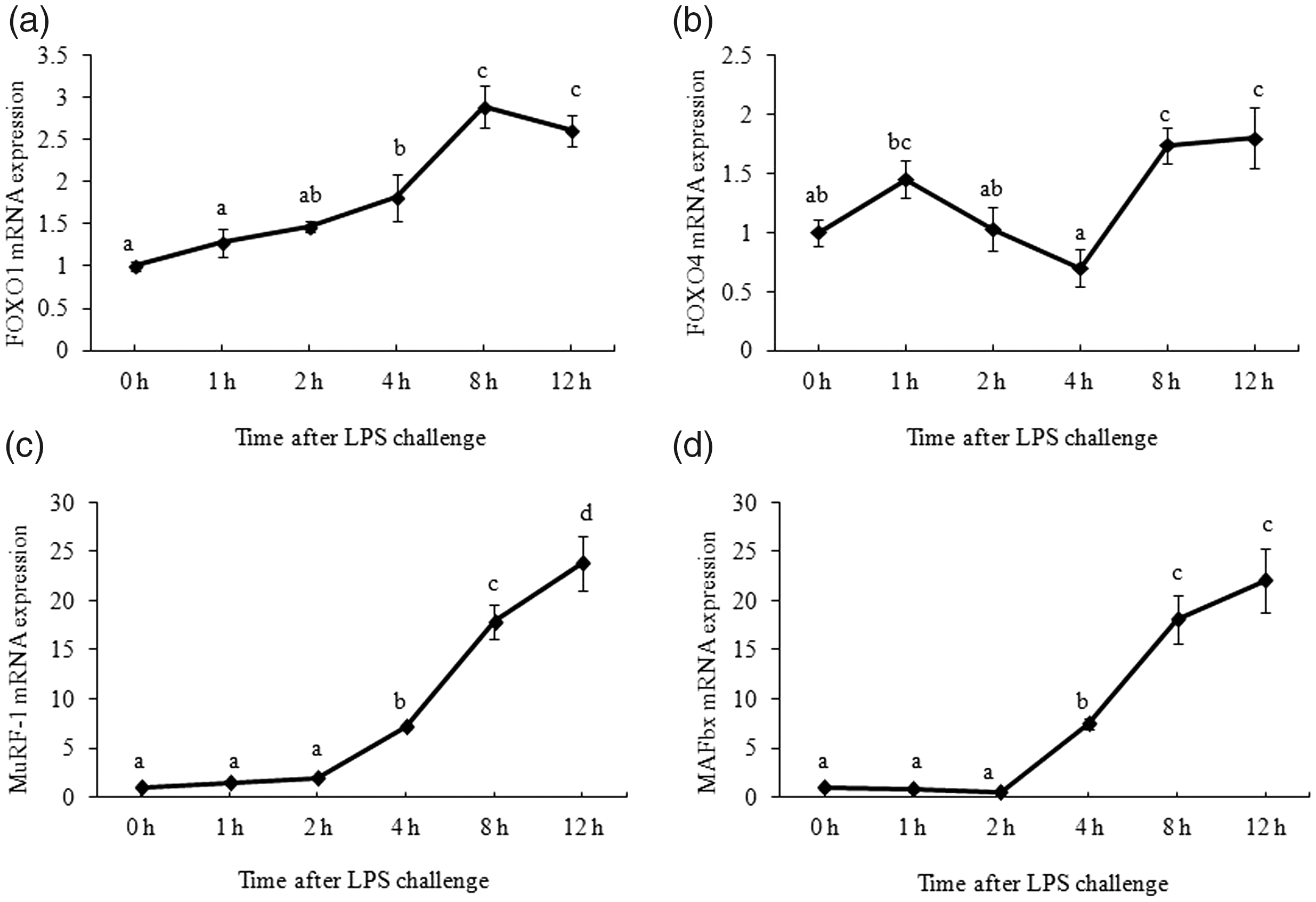
Effect of LPS on mRNA abundance of key genes in forkhead box O1 (FOXO)/ muscle atrophy F-box (MAFbx)-muscle-specific ring finger 1 (MuRF1) signaling pathway in muscle. Piglets were injected with 100 g/kg body mass (BM) or the equivalent amount of sterile saline. The piglets in the control group were slaughtered 0 h after saline injection. The piglets in LPS treatments were slaughtered at 1 h, 2 h, 4 h, 8 h, and 12 h after LPS injection. Values are single-point determinations and represent each of the time points listed above.
The expression of miRNAs in muscle
As shown in Table 2, compared with the control group, the expression of miR-135a and miR-370 reduced at 2 h (P < 0.05) and miR-122 expression decreased at 1 h (P < 0.05). However, after LPS challenge, the expression of miR-34a and miR-145 reached maximum expression levels at 1 h and 4 h, respectively (P < 0.05). In addition, miR-224 maximum expression level was observed at 1 h (P < 0.05) and reduced at 4 h after LPS challenge (P < 0.05). MiR-132 expression increased from 1 h (P < 0.05), reached the maximum level at 2 h (P < 0.05), and dropped to the basal level at 4 h after LPS challenge. Moreover, miR-130a expression decreased at 4 h after LPS challenge (P < 0.05), and miR-223 expression increased at 1 h (P < 0.05). Compared with the control, miR-146a had no significant change after LPS challenge (P > 0.05); however, its expression reduced significantly at 8 h compared with its expression at 2 h (P < 0.05). No significant difference was found in miR-127, miR-30b, miR-155, and miR-199a-3p expression after LPS treatment (P > 0.05).
Effect of LPS treatment on miRNAs abundance in muscle.e
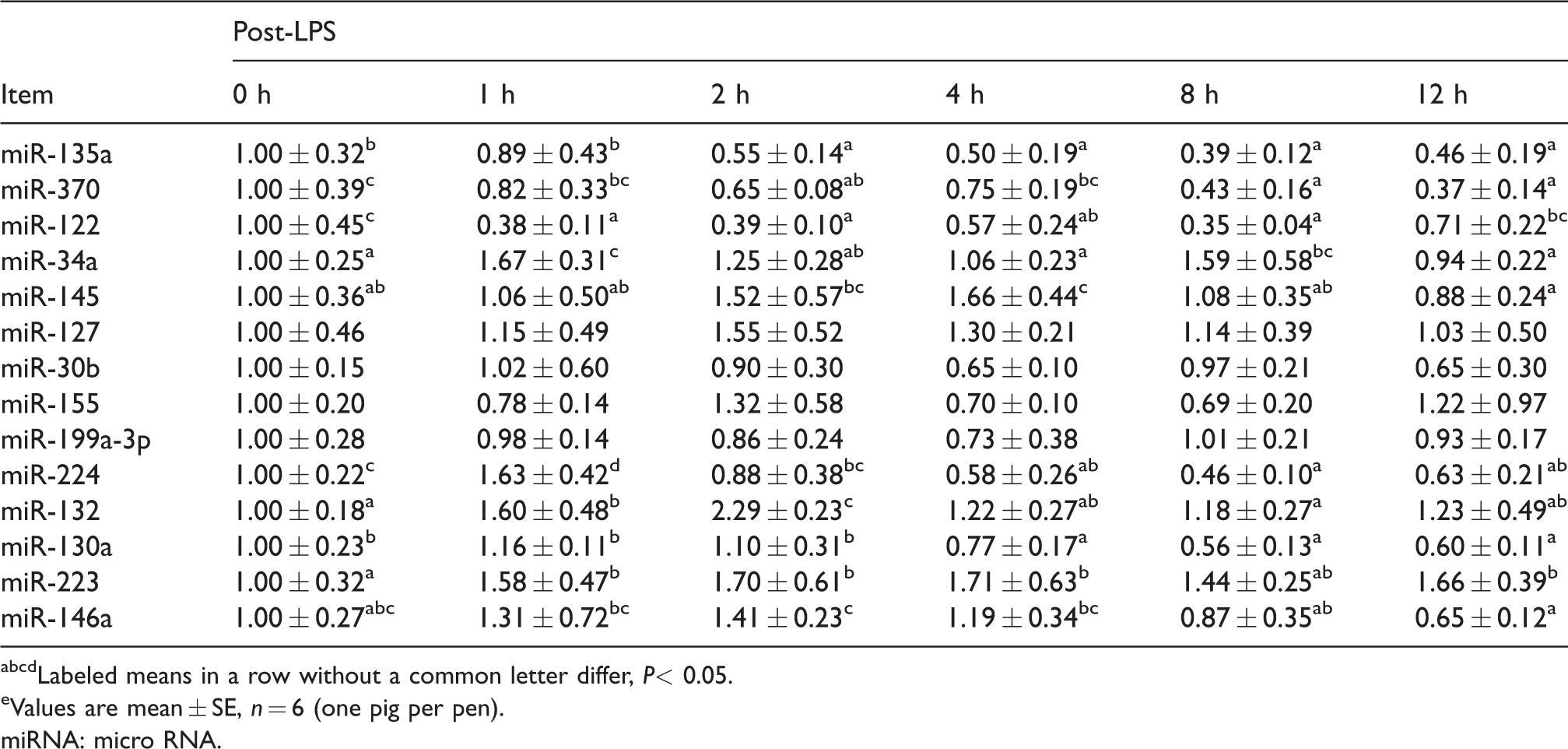
abcdLabeled means in a row without a common letter differ, P< 0.05.
eValues are mean ± SE, n = 6 (one pig per pen).
miRNA: micro RNA.
Discussion
The innate immune response is not only important for activating acquired immunity, but is the main contributor to acute inflammation induced by LPS. The inflammatory response mediated by pro-inflammatory cytokines can protect the body by removing detrimental stimuli and repairing damaged tissue. 22 However, over-production of pro-inflammatory cytokines may cause muscle atrophy evidenced by muscle protein degradation. 1 In addition, a lot of mammalian miRNA can regulate immune function 9 and inflammatory response. 10 Previous studies reported that innate immune cells could be in diverse states with different degrees of pro- and anti-inflammatory phenotypes accompanied with varying consequences for host defense and inflammation.14–16 Therefore, understanding the response and kinetics of changes in the expression of genes and miRNA related with inflammation and protein degradation following LPS challenge would be very important for further studies on alleviating inflammation and muscle protein loss.
As critical components of the innate immune response, TLR4 and NODs can initiate NF-κB activation through MyD88/IRAK1 signaling and RIPK2, respectively. Activation of NF-κB then triggers the expression of pro-inflammatory cytokines. In this study, we found that IL-1β, IL-6, and TNF-α mRNA expression reached maximum levels at an early stage after LPS injection; however, TLR4, MyD88, IRAK1, NOD1, and NOD2 mRNA expression reached peak levels at the later stage after LPS challenge. Basically, NF-κB mRNA expression increased at 1 h after LPS treatment, therefore these results indicated the elevated mRNA expression of pro-inflammatory cytokines could result from the increased NF-κB mRNA expression, and NF-κB could be activated through the MyD88-independent pathway at an early stage after LPS injection, which was in line with the report of Lu et al. 23 In addition, previous studies reported that TNF-α could directly activate NF-κB, 24 , 25 therefore increased NF-κB mRNA expression at an early stage after LPS treatment might partially result from elevated TNF-α mRNA expression, which was directly induced by LPS challenge.
LPS challenge can induce muscle atrophy, which is characterized by the elevated mRNA expression of FOXO1, MAFbx, and MuRF1. 7 , 8 Crossland et al. reported that pro-inflammatory cytokines could directly induce muscle protein degradation, or lead to muscle atrophy through activating these genes. 1 However, in our study, we found that FOXO1, FOXO4, MAFbx, and MuRF1 mRNA expression were elevated relatively late compared with IL-1β, IL-6, and TNF-α mRNA expression. The results suggested these increased pro-inflammatory cytokines at an early stage could result in the muscle wasting at a later stage, indicating the muscle-wasting response could happen later than the inflammatory response after LPS treatment.
Increasing evidence has indicated miRNA could stimulate or inhibit an inflammatory response. According to previous studies, a lack of miR-145 can induce pro-inflammatory signals in the innate immune system.26–28 Pekow et al. supposed that a loss of miR-145 pre-disposes patients to chronic inflammation in inflammatory bowel disease. 29 In line with these findings, we found that miR-145 expression reached the maximum level in the present study; however, the mRNA expression of IL-1β and TNF-α dropped to the basal level from the maximum 2 h after LPS challenge, indicating miR-145 had an anti-inflammatory response in a later stage of inflammation. In addition, miR-370 can reduce the inflammatory reaction by targeting TLR4. 30 In line with this study, in our experiment the minimum mRNA expression of miR-370 was observed at 12 h after LPS injection; on the contrary, the maximum mRNA expression of TLR4 and its downstream transcription factors MyD88 and IRAK1 was reached 12 h after LPS treatment.
MiR-127 can be significantly induced in an inflammation-related lung injury. 31 , 32 Increased expression of miR-127 in macrophages augments pro-inflammatory cytokine production. 33 Similarly, miR-155 is a major regulator of TLR signaling, and plays an important role in regulating immune cell development, function, and disease. 34 , 35 Tili et al. found a high expression of miR-155 could produce more TNF-α in mice challenged with LPS. 36 However, in this study, LPS injection had no effect on the expression of miR-155 and miR-127. The differences might result from the various LPS doses, tissues, animals, and sample collection times after LPS challenge. This could also explain our miR-30b expression results. In this study, we found LPS challenge had no effect on miR-30b expression over the experimental time points; however, Fordham et al. reported that enhanced miR-30b expression could inhibit the release of pro-inflammatory cytokines such as TNF-α and IL-6 induced by LPS. 37 miR-224 is a transcriptional target of NF-κB, and its up-regulation can activate the LPS- and NF-κB-dependent TNF-α inflammatory pathway. 38 In agreement with these results, in the present study we also found the maximum expression of miR-224 was reached at 1 h, as well as the mRNA expression of IL-1β and TNF-α after LPS injection.
In addition, we found the expression of miR-132, IL-1β, and NF-κB increased 1 h after LPS injection and the expression levels of miR-132 and IL-6 reached the maximum at 2 h. These results are inconsistent with previous studies, which reported that miR-132 played an anti-inflammatory role by suppressing IL-1β, IL-6, and NF-κB. 39 , 40 Li et al. thought there might be a highly negative correlation between miR-130a and TNF-α; 41 however, we did not find this relationship between them in our study. Moreover, Wang et al. reported that miR-223 inhibited inflammation by suppressing TLR4 signaling in macrophages. 42 However, in this study, we did not find this inhibitory response in muscle for miR-223 after LPS challenge.
Apart from stimulating or inhibiting inflammatory response, miRNA could be a new therapeutic frontier for muscle wasting in chronic kidney disease. 11 In this study, the mRNA expression of FOXO1, MuRF1, and MAFbx had a negative relationship with the expression of miR-34a, miR-224, miR-132, and miR-130a. These results indicated that these miRNAs might play an important role in inhibiting protein degradation by reducing the mRNA expression of FOXO1, MuRF1, and MAFbx in the late phase after LPS treatment. On the contrary, the expression of miR-145 was elevated 4 h after LPS injection, which was in line with the mRNA expression of FOXO1, MuRF1, and MAFbx, indicating that miR-145 could stimulate protein degradation after LPS injection.
In conclusion, the mRNA expression of pro-inflammatory cytokines such as IL-1β, IL-6, and TNF-α elevated in the early stages, mRNA expression of genes related to the TLR4 and NOD signaling pathway and protein degradation increased in the later phase after LPS injection, and miRNA related to muscle inflammation and protein degradation changed in the early stage after LPS treatment.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Project of Innovative Research Groups of the Natural Science Foundation of Hubei Province (2019CFA015), the National Natural Science Foundation of Hubei Province (2018CFB527), the Open Project of Hubei Key Laboratory of Animal Nutrition and Feed Science (Grant 201804), and the National Natural Science Foundation of China (31702203).