
Abstract
Asparagine (Asn), an activator of ornithine decarboxylase (ODC), stimulates cell proliferation in intestinal epithelial cells. We hypothesized that Asn can mitigate LPS-induced injury of intestinal structure and barrier function by regulating inflammatory signaling pathways. We executed the following experiment using weanling pigs for each of the groups: (1) non-challenged control; (2) LPS-challenged control; (3) LPS + 0.5% Asn; (4) LPS + 1.0% Asn. After 21-d feeding, pigs received an i.p. injection of either saline or LPS. Four h after injection, the mid-jejunum and mid-ileum samples were collected. We found that Asn restored ODC expression that was decreased by LPS treatment. Asn also restored intestinal morphology and barrier function that were impaired by LPS treatment. In addition, Asn down-regulated intestinal caspase-3 protein expression and TNF-α concentration, and decreased the mRNA expression of intestinal TLR4, TLR4 downstream signals (myeloid differentiation factor 88, IL-1 receptor-associated kinase 1 and TNF-α receptor-associated factor 6 and NOD1, NOD2 and their adaptor molecule (receptor-interacting serine/threonine-protein kinase 2). Moreover, Asn decreased p38 phosphorylation but increased ERK1/2 phosphorylation. Our results suggest that Asn improves intestinal integrity during an inflammatory insult, which appears to be related to the decrease of intestinal pro-inflammatory cytokine (via TLR4, NODs and p38) and of enterocyte apoptosis (via p38 and ERK1/2).
Introduction
The intestine is not only an important organ for nutrient digestion, absorption and metabolism, but is also the largest immune organ in the body, comprising more than 70% of the body’s immune cells. 1 However, multiple factors such as pathogenic and nonpathogenic challenges can damage intestinal mucosa structure and function.2–4
Activation of inflammatory signaling pathways plays a critical role in intestinal damage.5–7 TLRs and NODs are important protein families of inflammatory signaling pathways.8–10 TLRs and NODs are expressed in many tissues including the intestine,11–15 and play key roles in induction of innate antibacterial and inflammatory responses by recognition of PAMPs.8,9,16 Interactions of TLRs or NODs with their specific PAMPs trigger downstream signaling events that lead to activation of NF-κB and pathways involving MAPKs, which further provoke the expression of genes encoding pro-inflammatory cytokines, including IL-1β, IL-6 and TNF-α. 17 Consequently, pro-inflammatory cytokines mediate the host’s defense against invading pathogens. However, overproduction of these cytokines, especially TNF-α, can also elicit collateral injury in the intestine.
Many studies have shown that some specific amino acids, for example, glutamine (Gln), exert protective effects in the intestine under inflammatory conditions and can be used for treating diseases associated with intestinal injury. 18 However, Gln undergoes spontaneous degradation into ammonia and other metabolites at room temperature and is not a stable compound for animal feeds. 19 Asparagine (Asn), however, with a similar chemical structure to Gln, is more stable at room temperature and is a potential alternative to Gln. In fact, previous in vitro studies have shown that Asn protected against the induction of apoptosis in Gln-deprived rat intestinal epithelial cells.20,21 The protective effect of Asn was completely blocked by a Gln synthetase inhibitor, suggesting that the conversion of Asn to Gln is required for protection against apoptosis. 21 In addition, Asn increases ornithine decarboxylase (ODC) activity and cellular polyamine levels, and thus improves rat intestinal mucosal growth.20,22 However, to our best knowledge, in vivo studies that address whether Asn can modulate intestinal health under inflammatory stress are lacking. In this study, we used a piglet model in which the LPS of Escherichia coli was administrated to piglets to induce intestinal damage.23,24 We showed that Asn could attenuate intestinal injury induced by LPS, and regulate TLR4, NOD, p38 and ERK signaling.
Materials and methods
Animal care and diets
Ingredient composition of diets (as fed basis).

A rumen-stable fat powder, purchased from Berg + Schmidt (Hamburg, German).
In the 0.5% Asn diet, 1.35% alanine were replaced by 0.5% Asn, 0.68% alanine and 0.17% cornstarch. In the 1.0% Asn diet, 1.35% alanine was replaced by 1.0% Asn and 0.35% cornstarch. All diets were isonitrogenous.
A compound acidifier including lactic acid and phosphoric acid, provided by Wuhan Fanhua Biotechnology Co. (Wuhan, China).
The vitamin and mineral premix (defatted rice bran as carrier) provided the following amounts per kg of complete diet: retinol acetate, 2700 µg; cholecalciferol, 62.5 µg; DL-α-tocopheryl acetate, 20 mg; menadione, 3 mg; vitamin B12, 18 µg; riboflavin, 4 mg; niacin, 40 mg; pantothenic acid, 15 mg; choline chloride, 400 mg; folic acid, 700 µg; thiamin, 1.5 mg; pyridoxine, 3 mg; biotin, 100 µg; Zn, 80 mg (ZnSO4·7H2O); Mn, 20 mg (MnSO4·5H2O); Fe, 83 mg (FeSO4·H2O); Cu, 25 mg (CuSO4·5H2O); I, 0.48 mg (KI); Se, 0.36 mg (Na2SeO3·5H2O).
Based on diets containing cornstarch.
Calculated.
Analyzed.
Experimental design
The piglets (n = 6) were assigned randomly to one of four treatments: (1) non-challenged control (CONTR; pigs receiving a control diet and administered 0.9% NaCl solution); (2) LPS-challenged control [LPS; pigs receiving the same control diet and administered E. coli LPS (E. coli serotype 055:B5; Potency ≥ 500,000 EU/mg; Sigma Chemical, St. Louis, MO, USA)]; (3) LPS + 0.5% Asn treatment (pigs receiving a 0.5% Asn diet and administered LPS); and (4) LPS + 1.0% Asn treatment (pigs receiving a 1.0% Asn diet and administered LPS). The doses of Asn (purity > 99%; Amino Acid Bio-Chemical, Wuhan, China) were used in accordance with our previous study. 26 We supplemented 1.35%, 0.68% and 0% alanine (purity > 99 %; Amino Acid Bio-Chemical Co) to the control, 0.5% Asn and 1.0% Asn diets, respectively, to create isonitrogenous diets. BM and feed consumption were measured on d 1 and 21. On d 21, the challenged groups were administered i.p. with E. coli LPS at 100 µg/kg BM, and the control group was administered the equivalent amount of 0.9% NaCl solution. The dose of LPS was chosen in accordance with our previous studies.23,24
Intestinal sample collections
At 4 h after saline or LPS administration, all piglets were euthanized by i.m. injection of sodium pentobarbital (80 mg/kg BM). Immediately after slaughter, the small intestinal tract was dissected from the abdominal cavity. The intestinal samples were taken from two different sites along the proximo-distal axis: mid-jejunum at 50% between proximal jejunum and proximal ileum; mid-ileum at 50% between proximal ileum and ileo-caecal junction. The 3-cm and 10-cm segments were cut from the mid-jejunum and mid-ileum according to previous experiments.24,27 A variety of studies have shown that, within 3–6 h after injection, LPS results in morphologic impairment and barrier dysfunction in small intestine (such as jejunum and ileum), which is related with increased production of intestinal pro-inflammatory cytokines.23,28,29 Therefore, the time point of 4 h following saline or LPS administration was chosen for experimental measurements.
Intestinal segments (3 cm) were gently flushed and stored in 10% neutral-buffered formalin for histological analysis.23,24 Additional 10-cm intestinal samples were opened longitudinally and flushed gently to remove luminal chyme. Mucosa samples were then collected via scraping with sterile glass slides, rapidly frozen in liquid nitrogen and stored at –80℃. All intestinal samples were collected within 15 min of slaughter. The frozen intestinal mucosal samples were weighed, homogenized in ice-cold 0.9% NaCl solution with a 1:10 (w/v) ratio, and then centrifuged at 600 g for 10 min at 4℃ to collect the supernatants for analysis of diamine oxidase (DAO) activity and TNF-α concentration. The protein concentration of the supernatant was measured by the Bradford protein assay method. 30
Intestinal morphology analysis
After a 24-h fixation, the intestinal segments were dehydrated using a graded series of ethanol and cleared up with xylene, embedded in paraffin, cut into cross sections at a thickness of approximately 5 µm with a microtome, and then stained with hematoxylin and eosin. 31 The method for measurements of villus height and crypt depth was the same as described previously. 23
Intestinal mucosa DAO activity
Intestinal mucosa DAO activity was analyzed according to the method described by Liu et al. 32 The results were expressed as U/mg protein.
Intestinal mucosa TNF-α concentration
Intestinal mucosa TNF-α concentration was analyzed using a commercially available porcine ELISA kit (R&D Systems, Minneapolis, MN, USA) according to the manufacturer’s instructions. Briefly, the 100 µl of standard (23.4, 46.9, 93.8, 188, 375, 750, 1500 pg/ml TNF-α), control or diluted intestinal samples were added to a 96-well microtiter plate and incubated at room temperature (21–25℃) for 2 h. Following incubation, each well was aspirated and the plate was washed five times. After washing, a 100 -µl solution of conjugate was added to the wells. The plate was incubated at room temperature for 2 h and then washed five times. After that, 100 µl substrate solution was added. After incubation for 30 min at room temperature, 100 µl stop solution was added, and the plates were read at 450 nm using an ELISA plate reader (Model 550; Bio-Rad, Hercules, CA, USA). Results obtained using natural porcine TNF-α show dose–response curves that are parallel to the standard curves. The minimum detectable concentration was 3.7 pg/ml. The TNF-α concentration in intestine was expressed as pg/mg protein.
mRNA abundance analysis
Primer sequences used for real-time PCR.

Protein abundance analysis
The analysis of protein abundance in intestinal mucosa was carried out according to the method described previously. 24 Briefly, the intestinal mucosa samples (150–200 mg) were homogenized in 600 µl lysis buffer containing phenylmethanesulfonyl fluoride, protease and phosphatase inhibitors, and centrifuged at 12,000 g for 15 min at 4℃ to collect supernatants. Equal amounts of intestinal mucosa protein (65 µg) were transferred onto 10–15% polyacrylamide gel and separated via SDS-PAGE, and then transferred to polyvinylidene difluoride membranes for immunoblotting. Immunoblots were blocked with 5% nonfat milk in Tris-buffered saline/Tween-20 for 3 h at room temperature (21–25℃). The membranes were incubated overnight at 4℃ with primary Abs, and then with the secondary Ab for 2 h at room temperature. Specific primary Abs included rabbit anti-human ODC (polyclonal; 51 kDa; 1:500; #bs-1294R) from Bioss (Bioss Inc., Beijing, China), rabbit anti-human claudin-1 (polyclonal; 22 kDa; 1:1000; #51-9000) and rabbit anti-human occludin (polyclonal; 59 kDa, 1:1000; #ab31721) from Invitrogen (Invitrogen Technology Inc., Danvers, MA, USA), rabbit anti-human caspase-3 (polyclonal; 35 kDa; 1:1000; #9661), rabbit anti-human p38 (polyclonal; 43 kDa; 1:1000; #9212), rabbit anti-human phosphorylated p38 (polyclonal; 43 kDa; 1:1000; #9211), rabbit anti-rat extracellular signal-related kinase 1/2 (ERK1/2; polyclonal; 42/44 kDa; 1:1000; #9102) and rabbit anti-human phosphorylated ERK1/2 (polyclonal; 42/44 kDa; 1:1000; #9101) from Cell Signaling (Cell Signaling Technology Inc., Danvers, MA, USA), and mouse anti-β-actin (monoclonal; 42 kDa; 1:10000; #A2228) from Sigma Aldrich (Sigma Aldrich Inc., St. Louis, MO, USA). The secondary Abs included goat anti-rabbit IgG-HRP (polyclonal; 1:5000; #ANT020) and goat anti-mouse IgG-HRP (polyclonal; 1:5000; #ANT019) (Antgene Biotech, Wuhan, China). Blots were developed using an Enhanced Chemiluminescence Western blotting kit (Amersham Biosciences, Solna, Sweden), and visualized using a Gene Genome bioimaging system. Bands were analyzed by densitometry using Gene Tools software (Syngene, Frederick, MD, USA). β-Actin was used as a loading control in Western blotting analysis. The relative abundance of target proteins (ODC, claudin-1, occludin and caspase-3) were expressed as the target protein/β-actin protein ratio. Phosphorylated form of p38 and ERK1/2 were normalized with the total protein content of p38 and ERK1/2, respectively.
Eight samples from four treatments (n = 2) were run in one gel. Thus, 24 samples from four treatments (n = 6) were run in three gels at one time to minimize the gel-to-gel variations. The analyses were conducted in duplicate.
Statistical analysis
Data were analyzed by ANOVA appropriate for randomized complete-block designs using the GLM procedure of SAS (SAS Institute, Cary, NC, USA). Planned contrasts were used for comparison: CONTR vs. LPS, LPS vs. LPS + 0.5% Asn, LPS vs. LPS + 1.0% Asn. The LPS pigs (0% Asn) were compared with CONTR pigs to determine the effect of LPS challenge, and with LPS + 0.5% or 1.0% Asn pigs to determine the effect of Asn treatment within LPS-challenged pigs. Least significant differences were calculated with the LSMEANS statement. All data were expressed as means ± SE. P < 0.05 was considered as statistically significant.
Results
Growth performance
During the 21-d feeding trial (before LPS or saline injection), there were no differences in average daily feed intake (767 ± 68 g), average daily gain (488 ± 5 0 g) and feed/gain ratio (1.51 ± 0.14) among the four groups.
Intestinal morphology
Injection of LPS caused fever, vomiting, shivering, anorexia and inactivity within 1 h in all pigs (data not shown). The LPS pigs had higher crypt depth (P < 0.01; Figure 1) and lower villus height/crypt depth ratio (VCR) (P < 0.05) in ileum than CONTR pigs. Relative to LPS pigs, the LPS + 0.5% Asn pigs had higher villus height in ileum (P < 0.05). Relative to LPS pigs, the LPS + 1.0% Asn pigs displayed higher villus height (P < 0.01) and VCR (P < 0.05) in jejunum, and lower crypt depth in ileum (P < 0.01).
Effects of Asn supplementation on (a) villus height, (b) crypt depth and (c) villus height/crypt depth ratio of weaning piglets after 4 h of E. coli LPS challenge. Values are means with SE, n = 6 (1 pig/pen). CONTR vs. LPS: *P < 0.05, **P < 0.01; LPS vs. LPS + 0.5% Asn: +P < 0.05; LPS vs. LPS + 1.0% Asn: +P < 0.05, ++P < 0.01.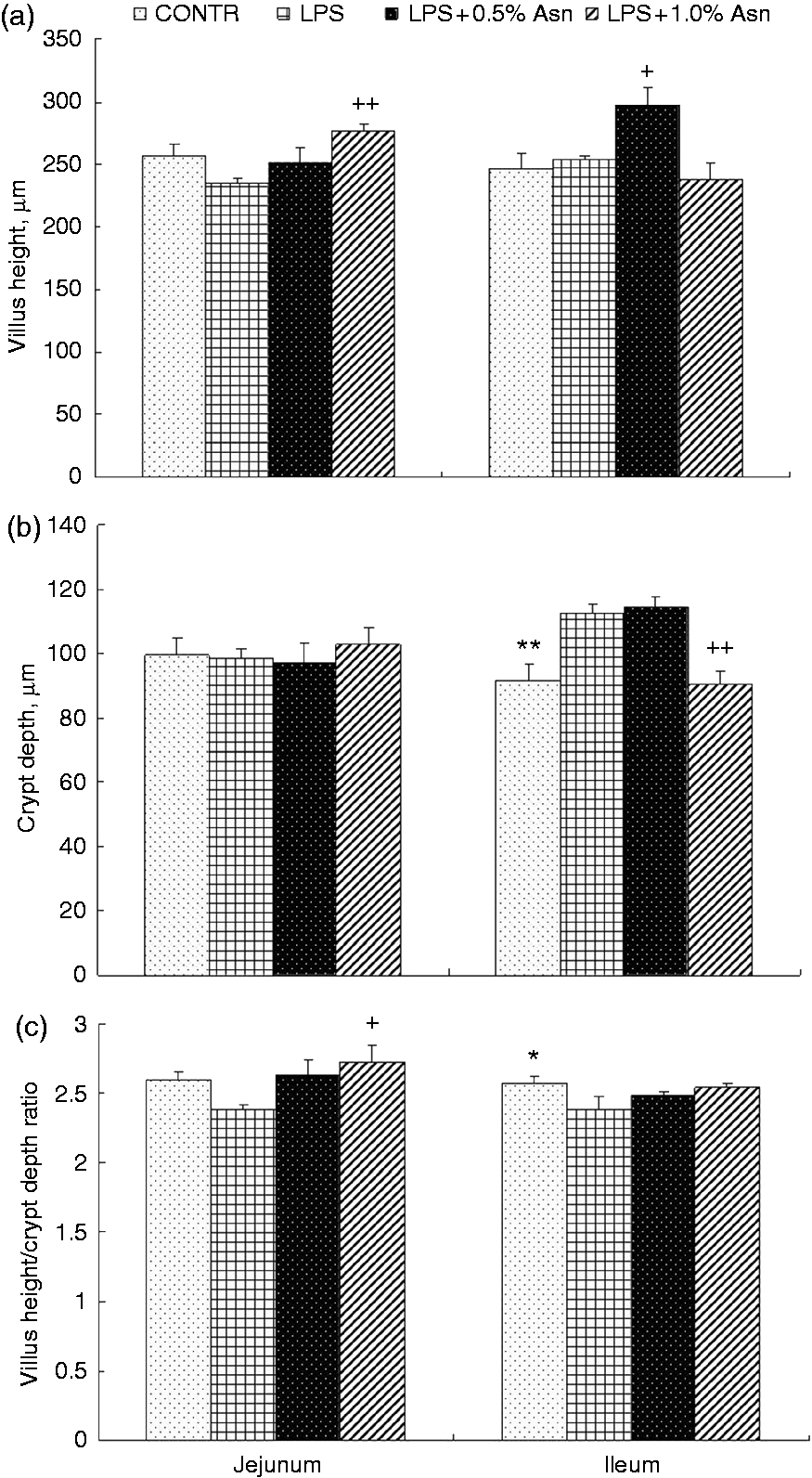
Intestinal DAO activity, and protein expression of ODC and tight junction proteins
Effects of Asn supplementation on intestinal DAO activity and protein expression of ODC and tight junction proteins after 4 h E. coli LPS challenge in weaned pigs.
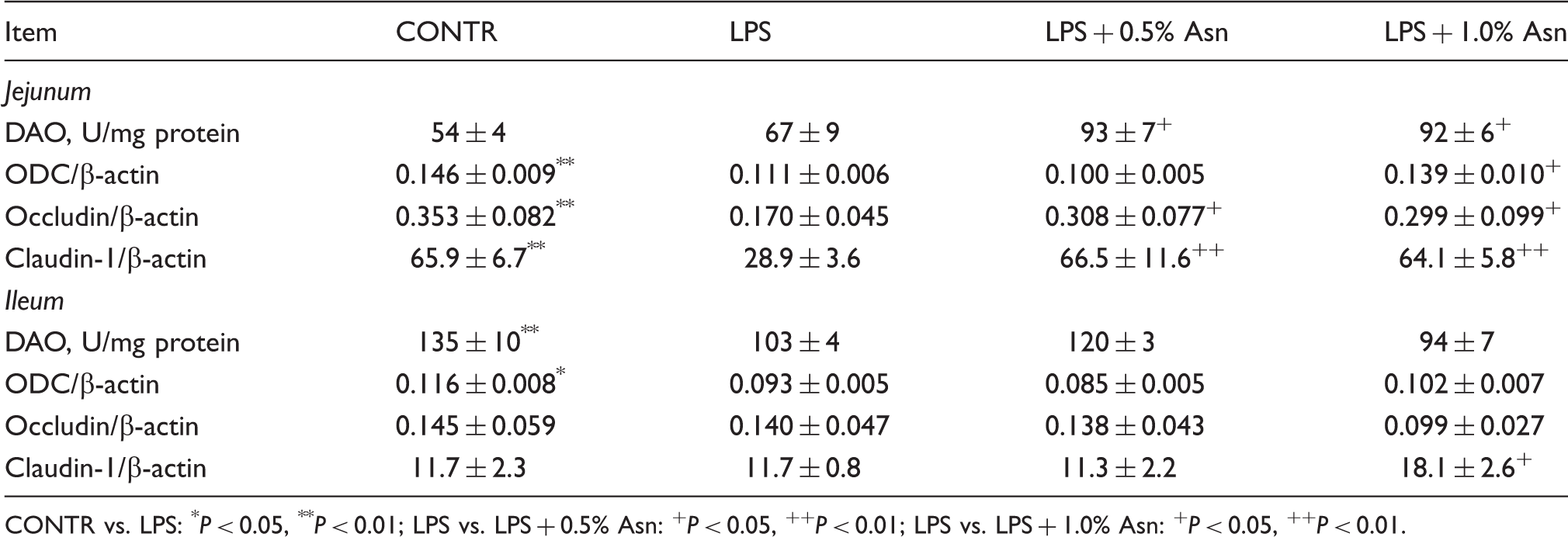
CONTR vs. LPS: *P < 0.05, **P < 0.01; LPS vs. LPS + 0.5% Asn: +P < 0.05, ++P < 0.01; LPS vs. LPS + 1.0% Asn: +P < 0.05, ++P < 0.01.

Effects of Asn supplementation on protein expression of ODC, occludin, claudin-1 and caspase-3 in (a) jejunum and (b) ileum after 4 h of E. coli LPS challenge in weaned piglets. The bands were the representative Western blot images of ODC, occludin, claudin-1, caspase-3 and β-actin.
Intestinal caspase-3 protein expression and TNF-α concentration
Effects of Asn supplementation on intestinal caspase-3 protein expression and TNF-α concentration after 4 h E. coli LPS challenge in weaned pigs.

CONTR vs. LPS: *P < 0.05; LPS vs. LPS + 0.5% Asn:+P < 0.05, ++P < 0.01, +++P < 0.001; LPS vs. LPS + 1.0% Asn: ++P < 0.01.
Intestinal mRNA expression of TLR4 and NODs and their downstream signals
Effects of Asn supplementation on intestinal mRNA abundance of TLR4 and NODs and their downstream signals after 4 h E. coli LPS challenge in weaned pigs.
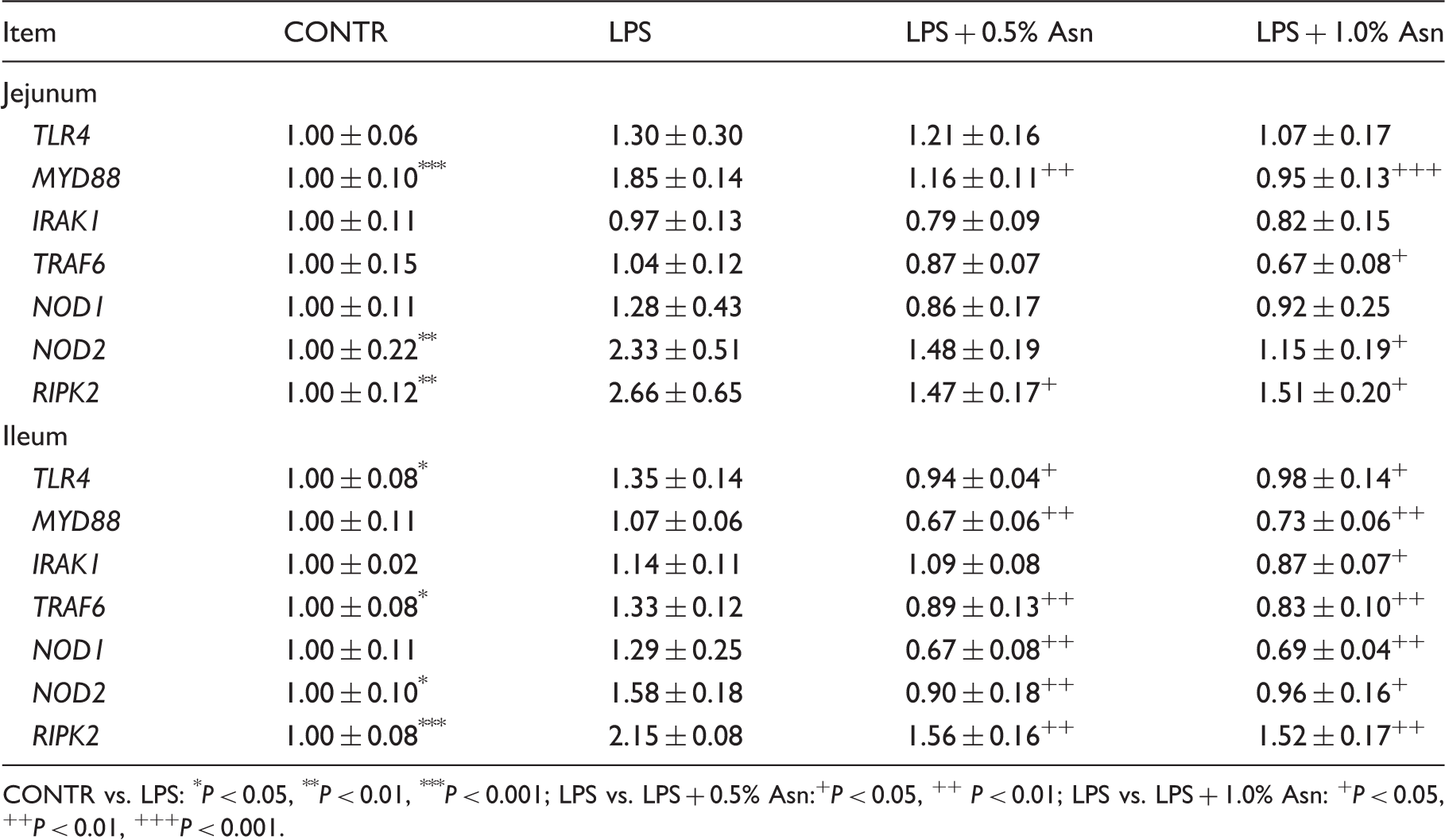
CONTR vs. LPS: *P < 0.05, **P < 0.01, ***P < 0.001; LPS vs. LPS + 0.5% Asn:+P < 0.05, ++ P < 0.01; LPS vs. LPS + 1.0% Asn: +P < 0.05, ++P < 0.01, +++P < 0.001.
Intestinal ERK and p38 protein expression
The LPS pigs had a higher p-p38/t-p38 ratio (P < 0.01; Figure 3) and a lower p-ERK/t-ERK ratio (P < 0.05) in jejunum. Relative to LPS pigs, both the LPS + 0.5% Asn pigs and the LPS + 1.0% Asn pigs had a lower ratio of p-p38/t-p38 in jejunum (P < 0.01), and higher p-ERK/t-ERK ratio in jejunum and ileum (P < 0.05).
Effects of Asn supplementation on the phosphorylated p38 (p-p38)/ total p38 (t-p38) ratio (a) and phosphorylated ERK (p-ERK)/ total ERK (t-ERK) ratio (b) of weaning piglets after 4 h of E. coli LPS challenge. The bands are the representative Western blot images of p-p38, t-p38, p-ERK and t-ERK. Values are means with SEs, n = 6 (1 pig/pen). CONTR vs. LPS: *P < 0.05, **P < 0.01; LPS vs. LPS + 0.5% Asn:++P < 0.01, +++P < 0.001; LPS vs. LPS + 1.0% Asn:+P < 0.05, ++P < 0.01.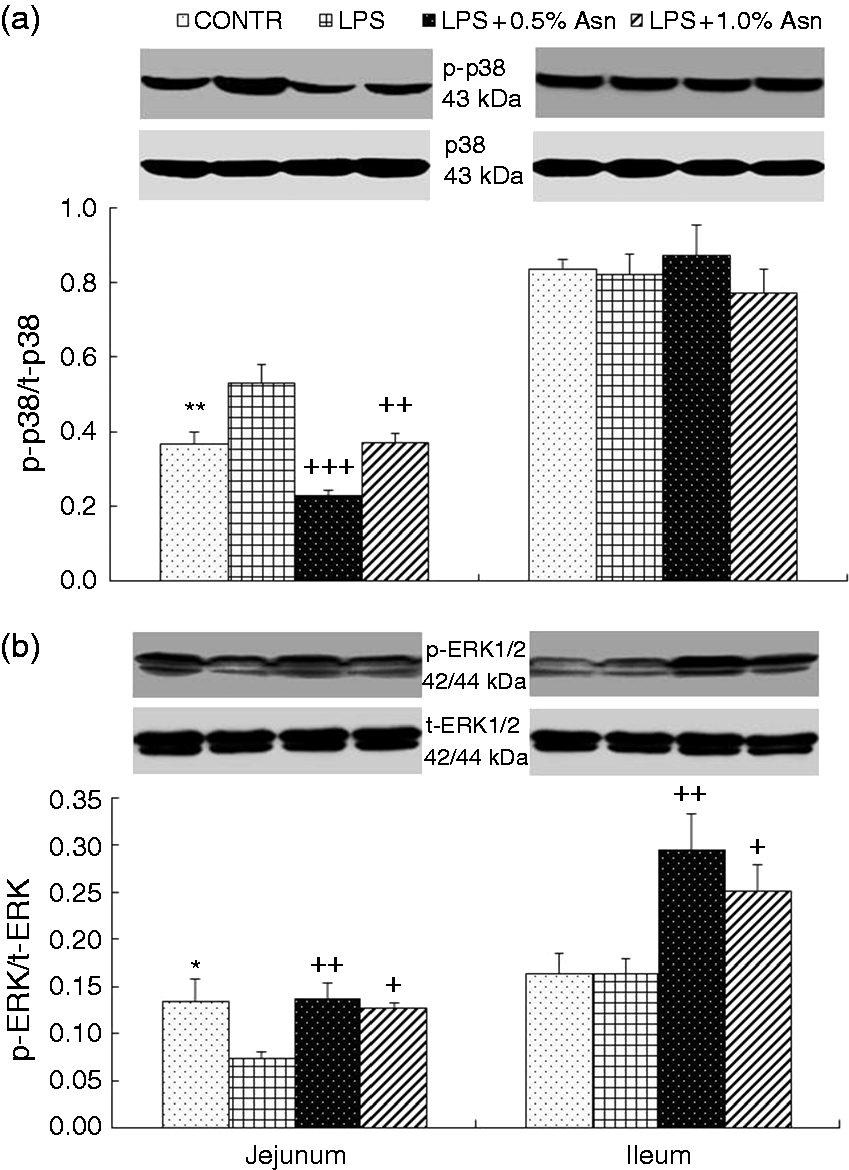
Discussion
Asn is an amino acid with similar chemical structure to Gln (with one less methylene group). It can be synthesized from aspartate and Gln. Thus, traditionally, it is considered as a nutritionally dispensable amino acid in mammals.35,36 However, some studies in vitro have shown that Asn, like Gln, is a powerful stimulator of ODC and stimulates cell proliferation in various cell types, including intestinal epithelial cells.20,22 To date, unlike Gln, the in vivo studies related to Asn modulation of intestinal health are limited. Thus, the aim of this experiment was to investigate whether Asn could mitigate intestinal damage induced by LPS challenge. Our previous studies (unpublished data) showed that, before LPS challenge, dietary addition of 0.5% and 1.0% Asn had no effect on growth performance, total and differential leukocyte counts and serum biochemical parameters of weanling pigs, suggesting that the Asn level in the basal diet was adequate to maintain growth and physiological function of weaned piglets in normal physiological condition. However, after LPS challenge, 0.5% Asn mitigated mass loss, and both 0.5% and 1.0% Asn mitigated the changes of total and differential leukocyte counts and serum biochemical parameters induced by LPS in weanling pigs, 26 indicating the importance of exogenous Asn supply under inflammatory situations. Therefore, in the current study, we focused on investigating the effect of dietary 0.5% and 1.0% Asn supplementation on intestinal variables in pigs subjected to LPS challenge, and did not investigate the effect of Asn in pigs without LPS challenge.
Indicators of intestinal morphology include villus height, crypt depth and VCR. Thus, an increase in villus height or VCR and a decrease in crypt depth indicate improvement in nutrient digestion and absorption.23,27 Many studies have shown that LPS challenge can result in a variety of morphologic alterations in the intestine such as submucosal edema, hemorrhage, mucosal necrosis and epithelial lifting, reduced villus height and increased crypt depth.23,28,29 As expected, our present results also showed that LPS challenge decreased ileal VCR and increased ileal crypt depth, indicating that LPS induced intestinal morphologic injury. Relative to LPS pigs, dietary addition of Asn increased villus height and VCR, and decreased crypt depth. These findings support the notion that Asn beneficially attenuates LPS-induced intestinal morphologic damage. Similarly, Papaconstantinou et al. 21 reported that addition of Asn resulted in increased cell number and viability in Gln-deprived rat intestinal epithelial cells. We speculate that the beneficial effect of Asn on intestinal morphology might be due to the following mechanisms. Polyamines are required for optimal cell growth and proliferation. 20 Intracellular polyamine levels are dependent primarily on the activity of ODC, which catalyzes the first rate-limiting step in polyamine biosynthesis, the decarboxylation of ornithine to form the diamine putrescine. 20 Many studies in vitro have demonstrated that Asn stimulates ODC activity and cellular polyamine levels, and thus improves intestinal mucosal growth and proliferation.20,22 In addition, our data in vivo also demonstrated that dietary Asn supplementation increased intestinal ODC protein expression in weanling piglets compared with LPS pigs, which further supports that Asn is a stimulator of ODC in intestine.
Intestinal epithelial barrier is important in the maintenance of gut homeostasis by preventing the penetration of luminal bacteria and dietary allergens into the mucosa. 1 Intestinal barrier function can be commonly evaluated by many indices including intestine mucosal permeability 37 and DAO activity. 38 Intestinal epithelial integrity is maintained by cohesive interactions between cells via forming tight junctions. 39 Members of the occludin and claudin family play a key role in tight junction formation, and determine permeability characteristics in many tissues, including the intestine. 39 Generally, decreased levels of tight junction proteins are associated with dysfunction of the epithelial barrier. 40 In addition to intestinal morphology, LPS challenge can have a detrimental effect on intestinal barrier function. 32 In accordance with this view, our present results also showed that LPS decreased ileal DAO activity, and jejunal claudin-1 and occludin protein expression. Relative to LPS pigs, dietary supplementation of Asn increased intestinal DAO activity, and claudin-1 and occludin protein expression, suggesting that Asn improves intestinal barrier function. Some research demonstrated that polyamines were necessary for the synthesis and stability of tight junction proteins and that polyamine depletion disrupts the epithelial barrier function, at least partially, by decreasing tight junction proteins.41,42 Our present data demonstrate that, as a stimulator of ODC, Asn might attenuate LPS-induced injury of intestinal barrier function partially by increasing the expression of intestinal tight junction proteins possibly via enhancing polyamine synthesis.
Intestinal epithelial cell turnover, including crypt cell proliferation, migration along the small intestine crypt–villus axis, differentiation and cell death via apoptosis, are dynamic processes. 23 To some degree, normal intestinal morphology is determined by the balance of epithelial cell turnover. 23 Increased enterocyte apoptosis may be one of key factors that contributes to intestinal morphological injury and barrier dysfunction. 23 Caspases are key mediators of programmed cell death (apoptosis). Among them, caspase-3 is a frequently activated death protease, catalyzing the specific cleavage of many key cellular proteins. 27 Caspase-3 has been shown to be an important marker of enterocyte apoptosis. 27 Consistent with improved intestinal structure and barrier function, dietary addition of Asn decreased ileal caspase-3 protein expression compared with LPS pigs. Similar to our data, Papaconstantinou et al. 21 reported that Asn protected against the induction of apoptosis in Gln-deprived rat intestinal epithelial cells. The protective effect of Asn was completely blocked by a Gln synthetase inhibitor, suggesting that the conversion of Asn to Gln was required for protection against apoptosis. 21 In our present study, it is possible that dietary Asn supplementation mitigated the LPS-induced intestinal structure and barrier damage partially via inhibiting enterocyte apoptosis.
We hypothesized that Asn exerted its beneficial effect on the intestine by reducing intestinal inflammatory response. A high level of intestinal pro-inflammatory cytokines, especially TNF-α, is an important marker of inflammation in intestine. 23 TNF-α is rapidly induced in the intestinal mucosa upon initial activation of immune cells, and is important for the further acceleration of the inflammatory response. 43 TNF-α has been identified to decrease the barrier function by disrupting tight junctions between the intestinal epithelial cells. 44 In the current study, consistent with improved intestinal structure and barrier function, Asn addition reduced intestinal TNF-α concentration compared with LPS pigs. These results support the notion that dietary supplementation with Asn is effective in reducing intestinal inflammatory response in weanling pigs.
To explore the molecular mechanism(s) by which Asn might alleviate the intestinal inflammatory response, we examined the role of inflammatory signaling pathways. TLRs and NODs are important protein families of inflammatory signaling pathways. They play a central role in detection of invading pathogens and induction of innate antibacterial and inflammatory responses by recognizing PAMPs.8–10 The intestine expresses multiple TLRs and NODs.12–15 Current research has demonstrated that activation of TLR and NOD signaling is associated with multi-layered inflammatory intestinal diseases.5,6 The binding of TLRs or NODs with their specific PAMPs triggers downstream signaling events that lead to activation of multiple intracellular signaling pathways, predominantly including NF-κB pathway, as well as MAPKs pathways, which include the Jun N-terminal kinase, ERK1/2 and p38.8,9 Activation of these intracellular signaling pathways further leads to expression and release of pro-inflammatory cytokines such as IL-1β, IL-6 and TNF-α. 17 In the current experiments, consistent with decreased intestinal TNF-α concentration, addition of Asn down-regulated intestinal mRNA expressions of TLR4 and its downstream signals (MyD88, IRAK1 and TRAF6), and NOD1, NOD2 and their adaptor molecule (RIPK2) compare with LPS pigs. In addition, Asn reduced intestinal p38 phosphorylation. The inhibition of p38, which can effectively suppress the expression of inflammatory mediators, significantly improves post-ischemic intestinal epithelial barrier functional recovery.45,46 Feng and Li 45 reported that human intestinal microvascular endothelial cells, intestinal epithelium, fibroblasts and myofibroblasts, which participate in inflammatory bowel disease, are affected by p38 directly or indirectly. Therefore, the beneficial effects of Asn addition on intestinal structure and barrier function might be related to reducing TNF-α via inhibition of TLR4 and NODs and p38 signaling.
Interestingly, unlike p38, our present study showed that LPS decreased intestinal ERK1/2 phosphorylation, and Asn attenuated LPS-induced decrease of ERK1/2 phosphorylation, which is inconsistent with decreased intestinal TNF-α concentration. In addition to regulation of inflammation, MAPKs also play a critical role in regulation of cell apoptosis. Some studies have shown that p38 and ERK1/2 may play opposing roles in cell apoptosis. The p38 pathway has a pro-apoptotic effect, and but ERK pathway has a protective effect on cell apoptosis.47–49 In some models of cell apoptosis, activation of p38 and concurrent inhibition of ERK are critical for induction of cell apoptosis.47–49 Thus, it is possible that Asn supplementation attenuated LPS-induced enterocyte apoptosis via inhibition of p38 and activation of ERK1/2.
In our current study, the effects of LPS challenge or Asn supplementation on some of intestinal measurements differ between two intestine segments. This may be associated with the difference in the anatomy and physiology among the various segments of the intestine. 50 Currently, it is not clear whether Asn affected intestinal integrity and inflammatory signaling pathways directly or indirectly by influencing ODC, and awaits further investigations.
In summary, as a stimulator of intestinal ODC, dietary Asn supplementation attenuates LPS-induced damage of intestinal morphology and barrier function. The beneficial effects of Asn on the intestine may be closely related to: (1) decreasing expression of intestinal pro-inflammatory cytokine via inhibiting TLR4 and NODs and p38 signaling pathways; (2) decreasing enterocyte apoptosis via inhibition of p38 and activation of ERK1/2.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The authors express their gratitude to the National Natural Science Foundation of China (31422053 and 31372318) and the Hubei Provincial Department of Education (T201508) for the financial support.