Melatonin is a well-documented hormone that plays central roles in the regulation of sleep–wake cycles. There is cumulative evidence to suggest that melatonin is also a pleiotropic regulator of inflammation, and luzindole has been widely used as a melatonin receptor antagonist. This study investigated the potential effects of luzindole on LPS/d-galactosamine (d-GalN)-induced acute hepatitis. The results indicated that treatment with luzindole alleviated histological damage in the liver, reduced the level of transaminases in plasma and improved the survival of LPS/d-GalN-exposed mice. Treatment with luzindole also suppressed the production of the pro-inflammatory cytokines TNF-α and IL-6 in LPS/d-GalN-exposed mice. In addition, treatment with luzindole inhibited the activation of caspase-3, -8 and -9, and suppressed the cleavage of caspase-3 and poly(ADP-ribose) polymerase. Therefore, treatment with luzindole attenuates LPS/d-GalN-induced acute liver injury, suggesting that luzindole might have potential value for the intervention of inflammation-based hepatic disorders.
The uncontrolled inflammatory response is one of the primary mechanisms underlying the development of acute hepatitis induced by infections, drugs, toxins and so on.1 LPS, the major virulence factor from Gram-negative bacteria, is a representative stimulator of inflammation, which is extensively involved in various inflammatory disorders, including acute hepatitis.2 Exposure to LPS in d-galactosamine (d-GalN)-sensitised mice is a widely used approach to induce acute hepatitis in experimental studies.3,4
The inflammatory response is tightly regulated by various endogenous factors, such as hormones, neurotransmitters and metabolites.5 Melatonin is a well-documented hormone that is mainly released from the pineal gland and plays central roles in the regulation of sleep–wake cycles.6 In addition, cumulative evidence suggests that melatonin also functions as a pleiotropic regulator of inflammation in peripheral tissues, and experimental studies have found that melatonin might act as both an activator and an inhibitor in inflammatory response.7
Luzindole is an antagonist of melatonin receptor, and its competitive binding to the melatonin receptor has been observed by competition experiments with 2-[125I]iodomelatonin.8,9 Luzindole has been widely used to block the activities of endogenous or exogenous melatonin in experimental studies.10,11 Although some studies have found that luzindole abolished the anti-inflammatory benefits of melatonin,12,13 treatment with luzindole also resulted in alleviated inflammatory injury under certain circumstances.14,15 In this study, the melatonin receptor antagonist luzindole was administered into mice with LPS/d-GalN-induced acute hepatitis, and its potential effects on inflammatory response, hepatocyte apoptosis, histological abnormalities and animal survival were determined.
Materials and methods
Animal and experimental materials
Male BALB/c mice (6–8 wk old, 18-20 g) were provided by the Experimental Animal Center of Chongqing Medical University. LPS (Escherichia coli, 055:B5), d-GalN and luzindole were purchased from Sigma–Aldrich (St Louis, MO). The mouse TNF-α and IL-6 ELISA kits were purchased from NeoBioscience (Shenzhen, PR China). The alanine aminotransferase (ALT) and aspartate aminotransferase (AST) assay kits were products from the Nanjing Jiancheng Bioengineering Institute (Nanjing, PR China). The Total Protein Extract Kit and the Caspase-3, -8, -9 Colorimetric Assay Kit were products from Beyotime Biotechnology Institute (Jiangsu, PR China). The in situ cell death assay kit was from Roche (Indianapolis, IN). The Abs against cleaved caspase-3, poly(ADP-ribose) polymerase (PARP) and β-actin were obtained from Cell Signaling Technology (Danvers, MA). The HRP goat anti-rabbit Ab, the BCA Protein Assay Kit and the enhanced chemiluminescence (ECL) reagent were from Pierce Biotechnology (Rockford, IL).
Experimental protocol
BalB/c male mice (n = 144) were divided into three sets. Set 1 comprised 32 mice randomised into four groups (n = 8): (a) the control group that received an i.p. injection of solvent; (b) the luzindole group that received an i.p. injection of the melatonin receptor antagonist luzindole at a dose of 40 mg/kg dissolved in 10% DMSO (diluted with edible oil; the dose of luzindole was chosen based on the previous literature and our preliminary experiment16); (c) the LPS/D-GalN group that received an i.p. injection of a mixture of LPS (10 μg/kg) and d-GalN (700 mg/kg); and (d) the luzindole intervention group where LPS/d-GalN was injected 30 min after the luzindole injection. Mice were sacrificed by cervical dislocation 90 min after LPS/d-GalN treatment, and the plasma samples were collected for detection of TNF-α and IL-6. Set 2 also comprised 32 mice divided into four groups (n = 8). Mice were sacrificed 6 h after LPS/d-GalN injection, and plasma samples and liver were collected. Set 3 comprised 80 mice divided into four groups (n = 20) and were used for survival observation. The mice were monitored for survival every 6 h for at least 7 d, and the survival rate of the mice was analysed by Kaplan–Meier curve.
Histopathological examination
The liver samples were fixed in 4% formaldehyde solution, follow-up conventional paraffin embedding, sectioning, hematoxylin and eosin staining and observation of photographs under an optical microscope (Olympus, Tokyo, Japan).
Analysis of AST and ALT levels
The levels of ALT and AST were determined for evaluating the degree of liver injury. The activity of liver enzymes was assessed by using the detection kits following the manufacturer’s instructions (Nanjing Jiancheng Bioengineering Institute).
ELISA assay
The plasma samples were collected 1.5 h after LPS/d-GalN exposure. Levels of TNF-α and IL-6 were measured by the ELISA kits following the manufacturer's instructions (NeoBioscience).
Determination of caspase activity
The liver samples were lysed according to the manufacturer’s instructions, and the protein content was determined by the Bradford method. A 96-well microtiter plate was taken, and each sample was set up with a measuring well and a blank well. The experimental procedure was carried out according to the instructions, and finally the OD value was measured at a wavelength of 405 nm. The relative activity of caspase was calculated and normalised by the protein content of each sample.
TUNEL assay
The sections were dewaxed by xylene for 1 h and hydrated in 100%, 90%, 80% and 70% gradient alcohol for 0.5 h at room temperature (25°C), and the slice was washed with PBS (three times for 5 min). The experiment was then performed according to the following steps: add proteinase K at 37°C for 0.5 h, wash the slice with PBS, incubate the slice with 0.1% Triton X-100 solution containing 0.1 g sodium citrate for 4 min at 4°C, wash the slice with PBS, incubate the slice with freshly prepared 3% H2O2 methanol solution for 20 min at room temperature, wash with PBS, add 20 µl TUNEL reaction solution at 37°C for 1 h, wash the slice with PBS, add 20 µl POD for 30 min at 37°C, and wash the slice with PBS. Finally, add the DAB solution to develop colour.
Western blot
The liver tissue was homogenised and centrifuged, and the supernatant was collected to extract total tissue protein. The protein concentration of each sample was determined by bicinchoninic acid assay (BCA), and the total protein in each sample was boiled and denatured. The protein samples were separated by a 12% SDS-PAGE and then transferred to a nitrocellulose membrane. After blocking with 5% defatted high protein milk, the membrane was incubated with the primary Ab overnight at 4°C. Then, the membrane was washed with TBST three times, and the secondary Ab was added and incubated at room temperature for 2 h. After washing, the blot was developed with the ECL reagent. As an internal parameter, β-actin was semi-quantitatively analysed for the grey value of each band.
Statistical analysis
The data are expressed as the mean ± SD. The differences in the measurement data between groups were compared by one-way ANOVA, followed by Turkey’s test. In addition, the survival data were expressed with the Kaplan–Meier curve and analysed by the log-rank test. All the statistical analyses was performed using SPSS software (v23, IBM Corp., Armonk, NY). A P value of < 0.05 was considered statistically significant.
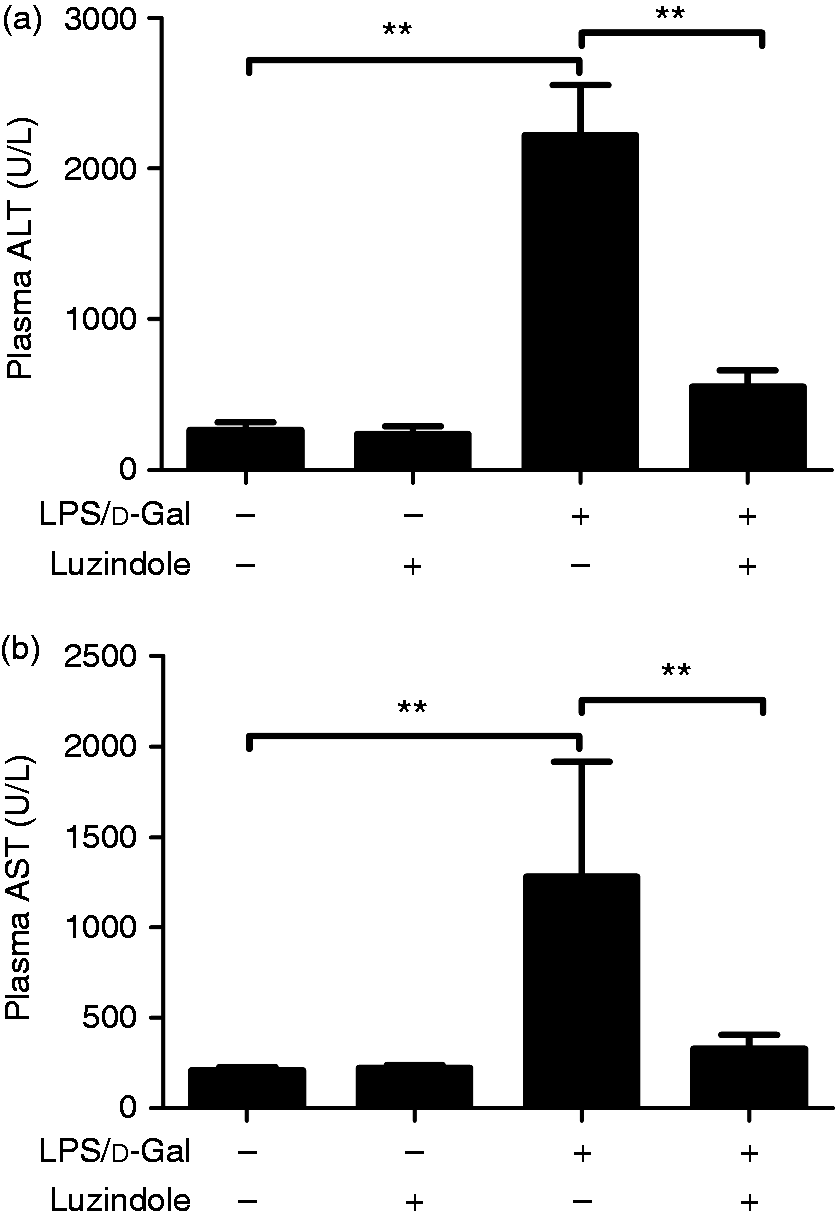
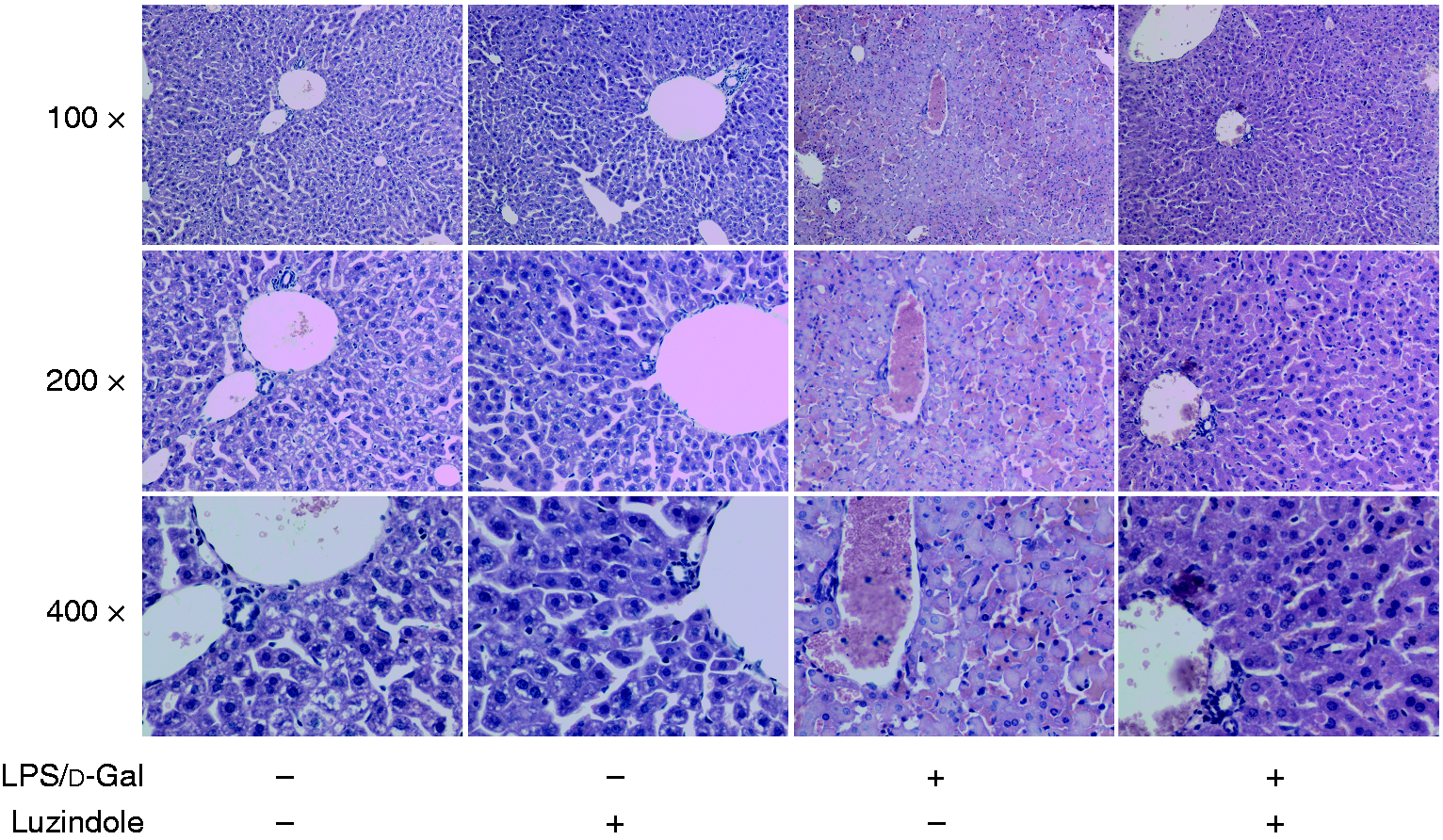
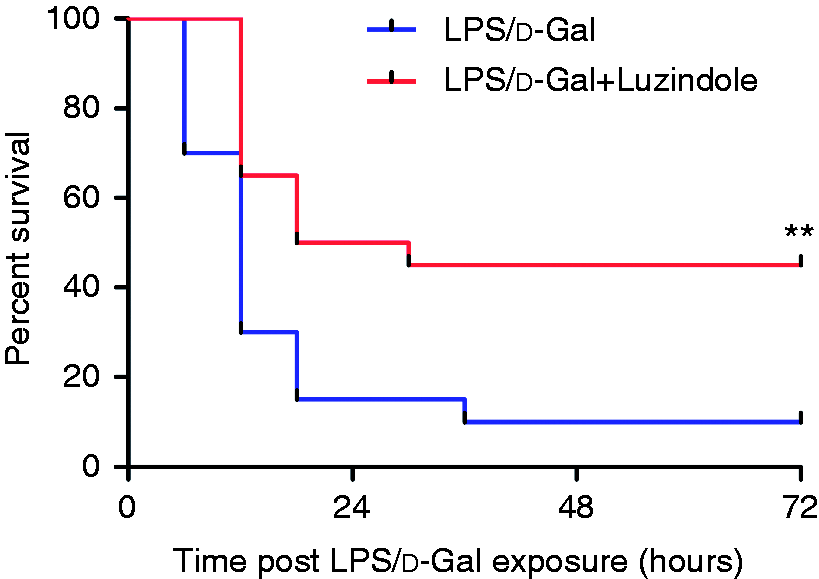
As shown in Figure 1, the level of ALT and AST in plasma significantly increased in LPS/d-GalN-exposed mice. The LPS/d-GalN-induced elevation of ALT and AST was suppressed in mice receiving the luzindole intervention. In the histological examinations, the liver lobule structure in mice exposed to LPS/d-GalN was blurred, and the hepatocyte cell line was disordered. These pathological changes were significantly alleviated in mice receiving the luzindole intervention (Figure 2). The survival analysis showed that treatment with luzindole significantly improved survival of LPS/d-GalN-exposed mice (Figure 3). These data suggested that LPS/d-GalN-induced liver injury was alleviated by luzindole.
Luzindole reduced LPS/d-GalN-induced elevation of aminotransferase (ALT) and aspartate aminotransferase. Acute hepatitis was induced in mice by i.p. injection of LPS/d-GalN, and luzindole was injected 30 min before LPS/d-GalN exposure. The plasma samples were collected 6 h after LPS/d-GalN exposure, and the levels of ALT (a) and AST (b) were determined. Data are expressed as the mean ± SD. Compared to the LPS/d-GalN group, n = 8, **P < 0.01.
Luzindole attenuated LPS/d-GalN-induced liver tissue damage. Acute hepatitis was induced in mice by i.p. injection of LPS/d-GalN, and luzindole was injected 30 min before LPS/d-GalN exposure. The liver tissue was collected 6 h after LPS/d-GalN exposure, and the liver sections were stained with hematoxylin and eosin for histological examination. The representative liver sections are shown (original magnification 100×, 200× and 400×).
Luzindole improved the survival of LPS/d-GalN-exposed mice. Acute hepatitis was induced in mice by i.p. injection of LPS/d-GalN, and luzindole was injected 30 min before LPS/d-GalN exposure. The mortality of the experimental animals was monitored every 6 h, and the cumulative survival curve was recorded by a Kaplan–Meier survival curve. Compared to the LPS/d-GalN group, n = 20, **P < 0.01.
Luzindole suppressed the induction of TNF-α and IL-6
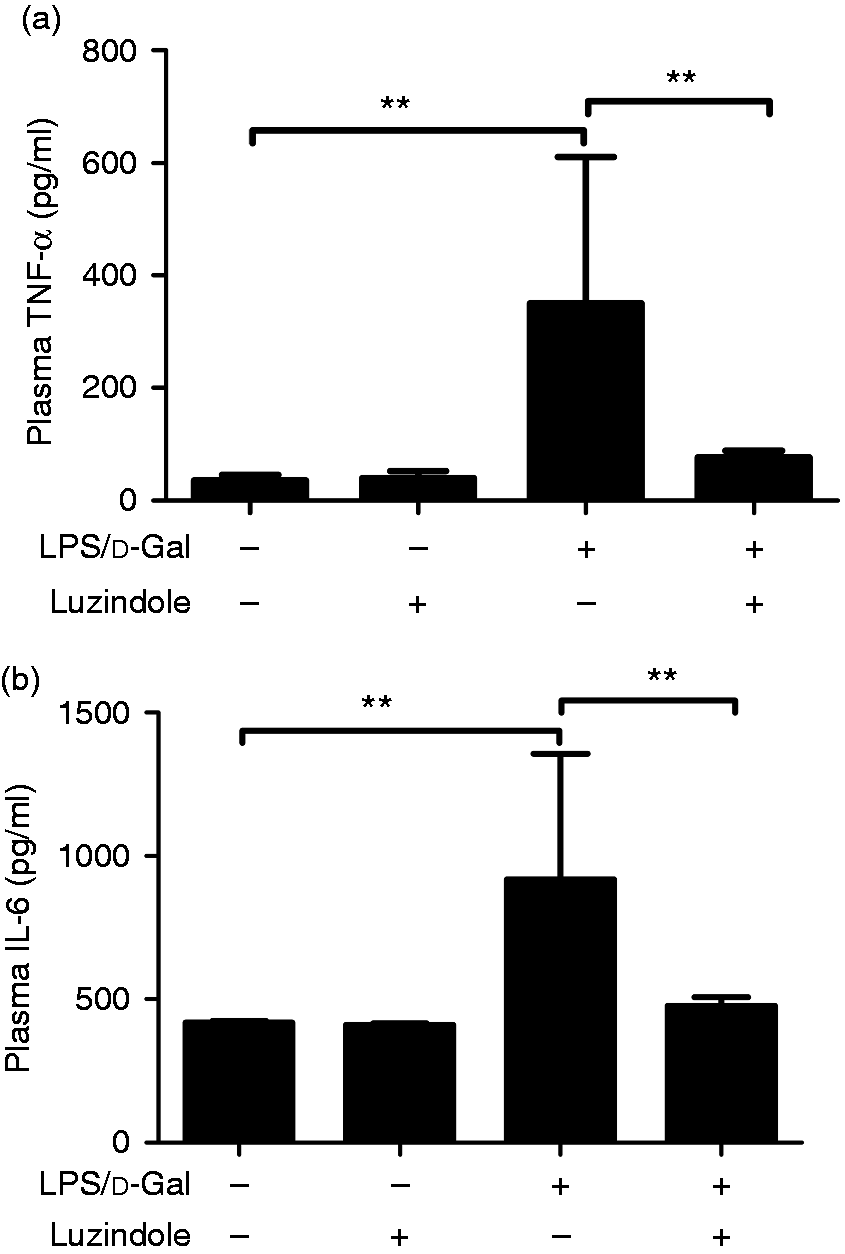
In mice exposed to LPS/d-GalN, the levels of TNF-α and IL-6, two representative pro-inflammatory cytokines,17 in the model group increased significantly. After the luzindole intervention, the induction of TNF-α and IL-6 was significantly suppressed (Figure 4), suggesting that LPS/d-GalN-induced inflammation was suppressed by luzindole.
Luzindole suppressed LPS/d-GalN-induced production of TNF-α and IL-6. Acute hepatitis was induced in mice by i.p. injection of LPS/d-GalN, and luzindole was injected 30 min before LPS/d-GalN exposure. The plasma samples were collected 1.5 h after LPS/d-GalN exposure, and the levels of TNF-α (a) and IL-6 (b) were determined. Data are expressed as the mean ± SD. Compared to the LPS/d-GalN group, n = 8, **P < 0.01.
Luzindole inhibited LPS/D-GalN-induced apoptosis
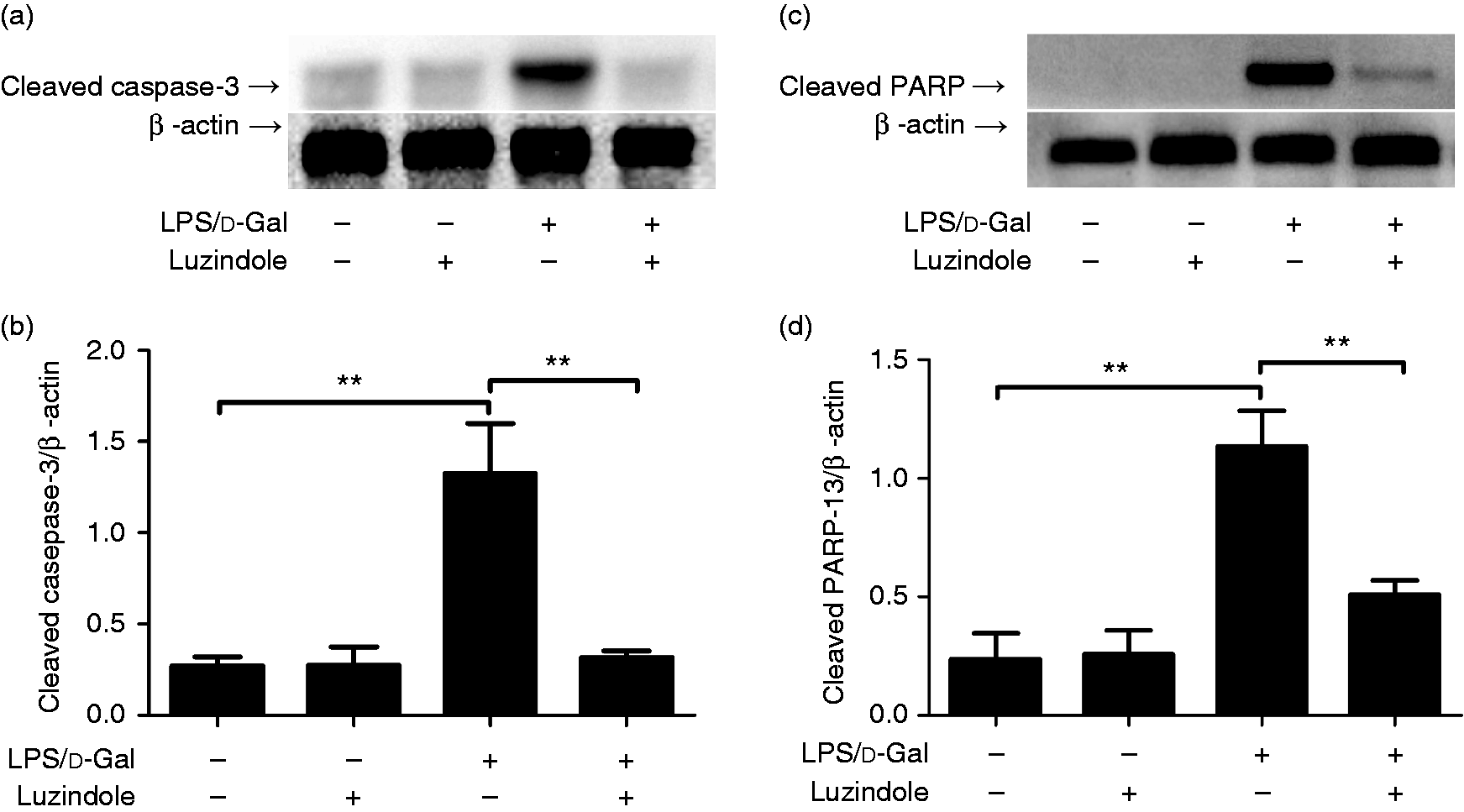
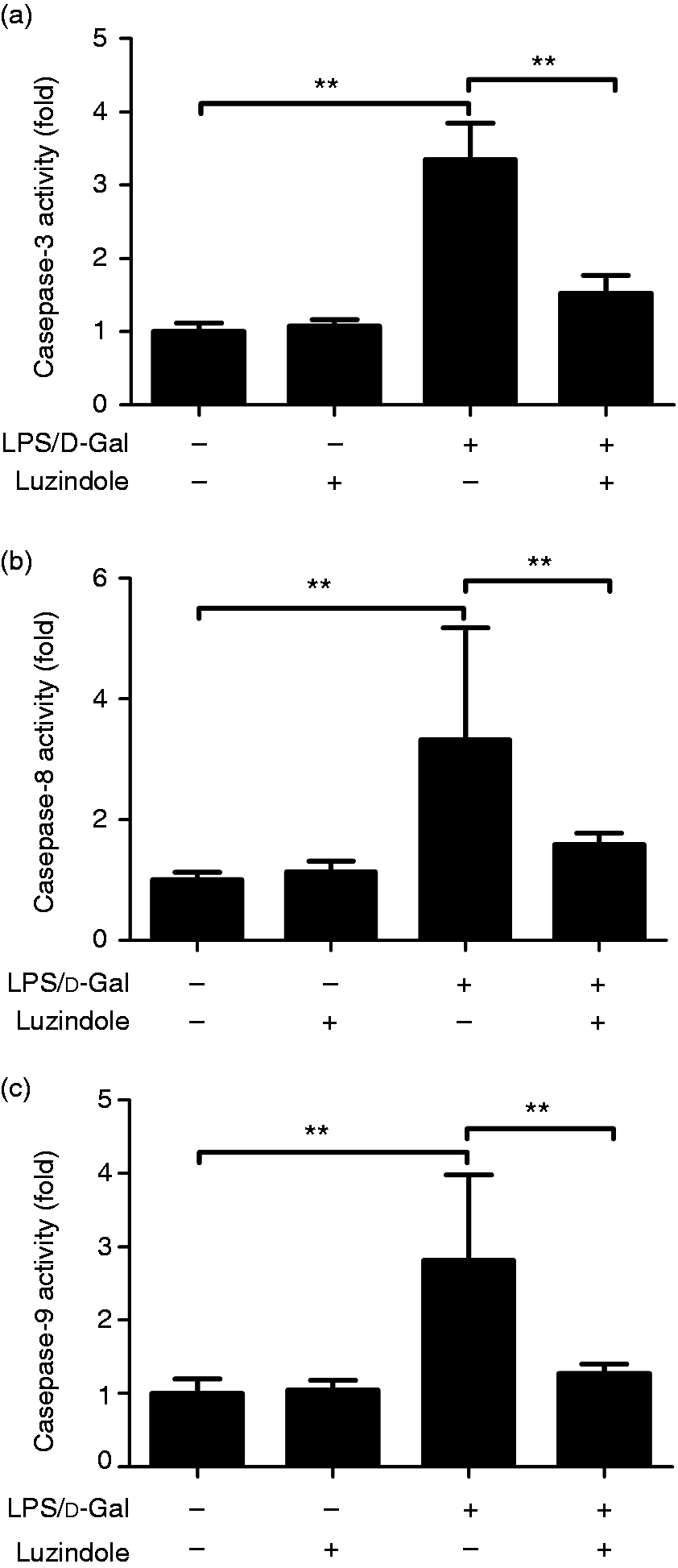
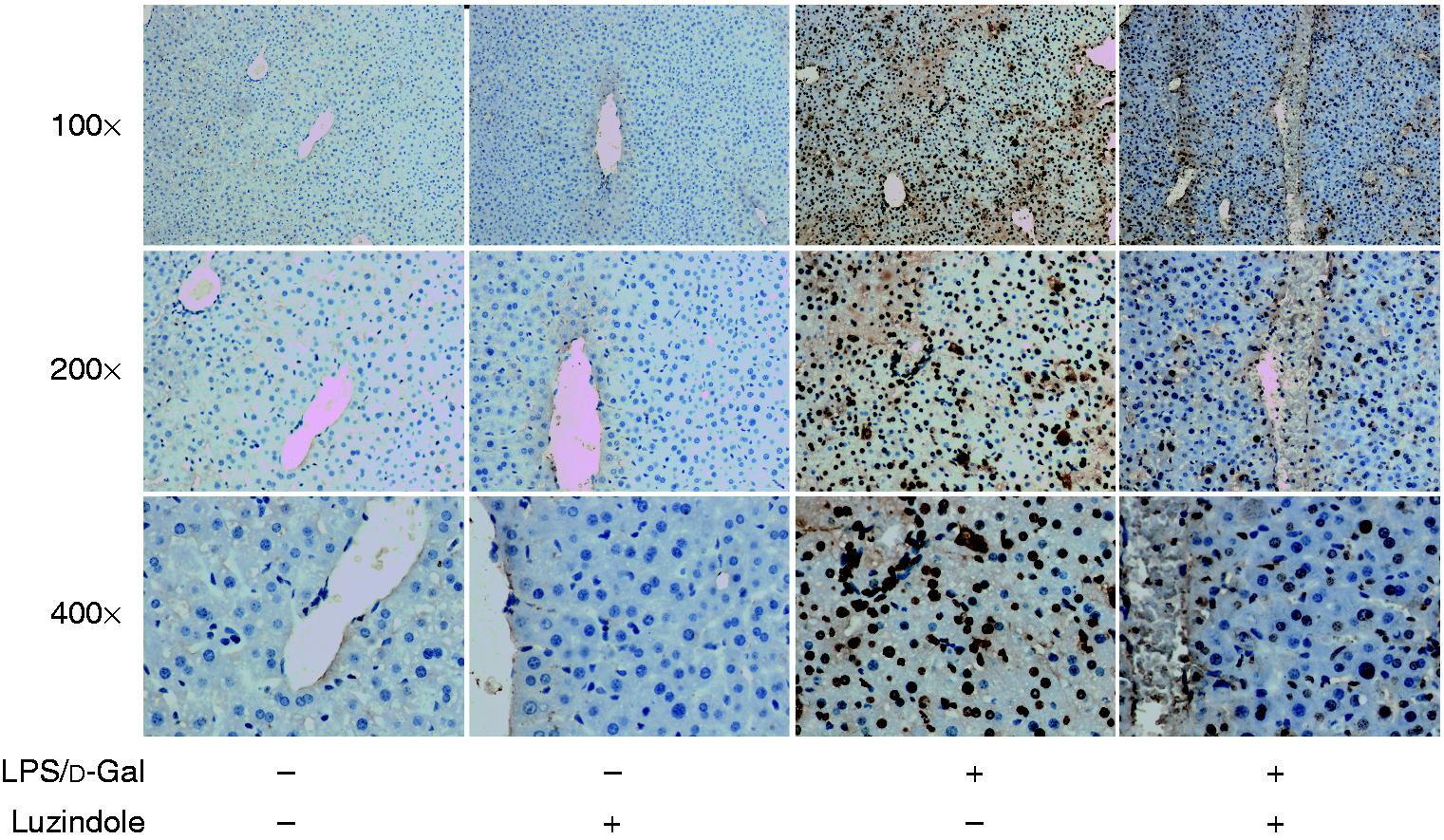
The data from Western blot analysis indicated that treatment with luzindole significantly inhibited LPS/d-GalN-induced cleavage of caspase-3 and PARP (Figure 5). In agreement with these findings, treatment with luzindole inhibited the activities of caspase-3, -8 and -9 in LPS/d-GalN-exposed mice (Figure 6). In addition, the TUNEL assay indicated that LPS/d-GalN-induced up-regulation of TUNEL-positive cells was inhibited by luzindole (Figure 7). These data suggested that LPS/d-GalN-induced apoptosis was suppressed by luzindole.
Luzindole suppressed LPS/d-GalN-induced cleavage of caspase-3 and poly(ADP-ribose) polymerase (PARP). Acute hepatitis was induced in mice by i.p. injection of LPS/d-GalN, and luzindole was injected 30 min before LPS/d-GalN exposure. The liver samples were collected 6 h after LPS/d-GalN exposure, and the levels of cleaved caspase-3 (a) and cleaved PARP (c) were determined. The blot was scanned, and the data are expressed as relative intensity units (b and d). Data are expressed as the mean ± SD. Compared to the LPS/d-GalN group, n = 4, **P < 0.01.
Luzindole inhibits LPS/d-GalN-induced activation of caspase cascade. Acute hepatitis was induced in mice by i.p. injection of LPS/d-GalN, and luzindole was injected 30 min before LPS/d-GalN exposure. The liver sample were collected 6 h after LPS/d-GalN exposure, and the activity of caspase-3 (a), caspase-8 (b) and caspase-9 (c) activities was measured. Data are expressed as the mean ± SD. Compared to the LPS/d-GalN group, n = 8, **P < 0.01.
Luzindole inhibits LPS/d-GalN-induced apoptosis. Acute hepatitis was induced in mice by i.p. injection of LPS/d-GalN, and luzindole was injected 30 min before LPS/d-GalN exposure. The liver samples were collected 6 h after LPS/d-GalN exposure, and the apoptotic cells were detected by TUNEL assay. The representative liver sections were shown (original magnification 100×, 200× and 400×).
Discussion
In addition to its critical regulatory roles in sleep–wake cycles, increasing evidence indicates that melatonin also plays important roles in the modulation of inflammatory responses.7 It has been reported that treatment with the melatonin receptor antagonist luzindole suppressed experimental autoimmune encephalomyelitis in mice.14 In agreement with these findings, the present study found that treatment with luzindole significantly alleviated LPS/d-GalN-induced acute hepatitis, suggesting that luzindole might have potential value in the intervention of inflammation-based liver injury.
In the LPS/d-GalN model, the activation of inflammatory cells and the induction of pro-inflammatory mediators are the primary mechanisms underlying the development of acute liver injury.18,19 Melatonin has been reported to function as a dual regulator in inflammation.7 Some studies have found that treatment with melatonin resulted in beneficial outcomes in experimental animals with colitis and those with pancreatitis ischaemia/reperfusion injury.20–22 On the contrary, treatment with melatonin also promoted an inflammatory response via activating 5-lipoxygenase and extending the lifespan of recruited leucocytes under certain circumstances.23 In the present study, the alleviated liver injury in luzindole-treated mice was associated with suppressed production of pro-inflammatory cytokines, including TNF-α and IL-6. These data suggest that the anti-inflammatory potential of luzindole might be responsible for the beneficial outcomes in LPS/d-GalN-exposed mice.
Hepatocyte apoptosis is an important pathological manifestation of acute hepatitis induced by LPS/d-GalN as well as other harmful factors.24,25 In the principle of LPS/d-GalN-induced acute hepatitis, the production of TNF-α plays central roles in the induction of hepatocyte apoptosis.26 TNF-α initiates the death receptor-dependent apoptotic pathway via its receptor, which leads to the activation of caspase cascade.27 Finally, the activated executive caspase-3 cleaves functional proteins such as PARP, which is a crucial molecular event for the induction of apoptosis.28 In the present study, the activity of caspase-8, -9 and -3, the level of cleaved caspase-3 and cleaved PARP, and the count of TUNEL-positive cells were markedly reduced in mice that had received luzindole treatment, suggesting that LPS/d-GalN-induced hepatocyte apoptosis was suppressed by luzindole.
It has been reported that mice lacking the genes encoding TNF-α or its receptor were resistant to LPS/d-GalN-induced liver injury.24 Therefore, suppression of TNF-α production might be an important reason for the suppressed apoptosis in luzindole-treated mice. In addition to the regulatory roles in the production of pro-inflammatory cytokines, several studies have found that melatonin is also directly involved in regulating apoptosis. For example, treatment with melatonin induced apoptosis in gastric cancer cells, breast cancer cells, cervical cancer cells and osteoblastic cells.29–32 Treatment with melatonin also enhanced apoptosis induced by other stimulators such as docetaxel and cisplatin.31,33 Interestingly, it has been reported that treatment with melatonin induced apoptosis in hepatoma cells.34,35 Therefore, it is possible that treatment with luzindole might block the pro-apoptotic property of melatonin and provides protective benefits in LPS/d-GalN-exposed mice.
Although luzindole has been widely used as a melatonin receptor antagonist, experimental studies have also found that some pharmacological activities of melatonin could not be blocked by luzindole. In rats with experimental reflux oesophagitis, luzindole failed to antagonise the protective effects of melatonin.36 In addition, luzindole also failed to reverse the effects of melatonin on the proliferation of granulosa cells under thermal stress.37 Therefore, it could not be excluded that the protective benefits of luzindole in the present study might be independent of melatonin.
Interestingly, a study found that luzindole, but not other melatonin receptor antagonists, suppressed LPS-induced generation of malondialdehyde, a representative marker for oxidative stress.38 In addition, an in vitro study investigated the radical scavenging activity of luzindole by using a spectrophotometrical scavenger competition assay. The study found that luzindole reduced the level of radical more intensely than did ascorbic acid.39 This evidence suggests that luzindole might have anti-oxidative activity, which could be responsible for some of the melatonin receptor–independent pharmacological effects of luzindole.
In fact, reactive oxygen species play crucial roles in the LPS/d-GalN model. It has been reported that genetic deficiency or pharmacological inhibition of antioxidant enzyme resulted in exacerbated liver injury in mice exposed to LPS/d-GalN.40,41 On the contrary, induction of heme oxygenase-1 or overexpression of thioredoxin attenuated LPS/d-GalN-induced liver injury.42,43 In addition, supplementing with antioxidants, such as edaravone, N-acetylcysteine and α-lipoic acid, markedly suppressed LPS/d-GalN-induced elevation of ALT, inhibited the up-regulation of pro-inflammatory cytokines and alleviated histological abnormalities.44–46 Thus, the anti-oxidative effects of luzindole might also contribute to the beneficial outcomes in the present study.
Taken together, the present study found that treatment with the widely used melatonin receptor antagonist luzindole suppressed LPS/d-GalN-induced inflammatory response, attenuated hepatocyte apoptosis and improved the survival of the experimental animals. These beneficial outcomes might result from the prevention of the pro-inflammatory and pro-apoptotic effects of melatonin by luzindole or could be attributed to the anti-oxidative effect of luzindole, but the detailed underlying mechanisms remain to be further investigated. This study suggests that luzindole might have potential value for the pharmacological intervention of inflammation-based hepatic disorders.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship and/or publication of this article: this work was supported by the grants from the National Natural Science Foundation of China (No. 81671953).
ORCID iDs
Yisheng Luo
Yongqiang Yang
References
1.
MontriefTKoyfmanALongB.Acute liver failure: a review for emergency physicians.Am J Emerg Med2019;
37:329–337.
2.
VergadiEVaporidiKTsatsanisC.Regulation of endotoxin tolerance and compensatory anti-inflammatory response syndrome by non-coding RNAs. Front Immunol2018;
9:2705.
3.
KimHJJoeYRahSY, et al.
Carbon monoxide-induced TFEB nuclear translocation enhances mitophagy/mitochondrial biogenesis in hepatocytes and ameliorates inflammatory liver injury. Cell Death Dis2018;
9:1060.
4.
WuJZhaoYParkYK, et al. Loss of PDK4 switches the hepatic NF-κB/TNF pathway from pro-survival to pro-apoptosis. Hepatology. Epub ahead of print 30 March 2018; 68: 1111–1124.
5.
FullertonJNGilroyDW.Resolution of inflammation: a new therapeutic frontier.Nat Rev Drug Discov2016;
15:551–567.
6.
ClaustratBLestonJ.Melatonin: physiological effects in humans.Neurochirurgie2015;
61:77–84.
7.
RadognaFDiederichMGhibelliL.Melatonin: a pleiotropic molecule regulating inflammation.Biochem Pharmacol2010;
80:1844–1852.
8.
DubocovichMLShankarGMickelM.2-[125I]iodomelatonin labels sites with identical pharmacological characteristics in chicken brain and chicken retina.Eur J Pharmacol1989;
162:289–299.
9.
BlazynskiCDubocovichML.Localization of 2-[125I]iodomelatonin binding sites in mammalian retina.J Neurochem1991;
56:1873–1880.
10.
DengSLSunTCYuK, et al.
Melatonin reduces oxidative damage and upregulates heat shock protein 90 expression in cryopreserved human semen.Free Radic Biol Med2017;
113:347–354.
11.
ShuTFanLWuT, et al.
Melatonin promotes neuroprotection of induced pluripotent stem cells-derived neural stem cells subjected to H2O2-induced injury in vitro.Eur J Pharmacol. 2018;
825:143–150.
12.
FinkTGlasMWolfA, et al.
Melatonin receptors mediate improvements of survival in a model of polymicrobial sepsis.Crit Care Med2014;
42:e22–31.
13.
MathesAMKubulusDPradaruttiS, et al.
Melatonin pretreatment improves liver function and hepatic perfusion after hemorrhagic shock.Shock2008;
29:112–118.
14.
ConstantinescuCSHilliardBVenturaE, et al.
Luzindole, a melatonin receptor antagonist, suppresses experimental autoimmune encephalomyelitis.Pathobiology1997;
65:190–194.
15.
SugawaraTSievingPAIuvonePM, et al.
The melatonin antagonist luzindole protects retinal photoreceptors from light damage in the rat. Invest Ophthalmol Vis Sci1998;
39:2458–2465.
16.
ChernCMLiaoJFWangYH, et al.
Melatonin ameliorates neural function by promoting endogenous neurogenesis through the MT2 melatonin receptor in ischemic-stroke mice.Free Radic Biol Med2012;
52:1634–1647.
17.
ZhouZXuMJGaoB.Hepatocytes: a key cell type for innate immunity.Cell Mol Immunol2016;
13:301–315.
18.
HeYXiaZYuD, et al.
Hepatoprotective effects and structure-activity relationship of five flavonoids against lipopolysaccharide/d-galactosamine induced acute liver failure in mice.Int Immunopharmacol2019;
68:171–178.
19.
TangFFanKWangK, et al.
Amygdalin attenuates acute liver injury induced by d-galactosamine and lipopolysaccharide by regulating the NLRP3, NF-κB and Nrf2/NQO1 signalling pathways.Biomed Pharmacother2019;
111:527–536.
20.
ChamanaraMRashidianAMehrSE, et al.
Melatonin ameliorates TNBS-induced colitis in rats through the melatonin receptors: involvement of TLR4/MyD88/NF-κB signalling pathway.Inflammopharmacology2019;
27:361–371.
21.
LePHKuoCJChengHT, et al.
Melatonin attenuates acute pancreatitis-induced liver damage through Akt-dependent PPAR-γ pathway.J Surg Res2019;
236:311–318.
22.
ShiSLeiSTangC, et al.
Melatonin attenuates acute kidney ischemia/reperfusion injury in diabetic rats by activation of the SIRT1/Nrf2/HO-1 signaling pathway.Biosci Rep2019;
39.
23.
KepkaMSzwejserEPijanowskiL, et al.
A role for melatonin in maintaining the pro- and anti-inflammatory balance by influencing leukocyte migration and apoptosis in carp.Dev Comp Immunol2015;
53:179–190.
24.
BrennerCGalluzziLKeppO, et al.
Decoding cell death signals in liver inflammation.J Hepatol2013;
59:583–594.
25.
HuKGongXAiQ, et al.
Endogenous AMPK acts as a detrimental factor in fulminant hepatitis via potentiating JNK-dependent hepatocyte apoptosis.Cell Death Dis2017;
8:e2637.
26.
WenJLinHZhaoM, et al.
Piceatannol attenuates d-GalN/LPS-induced hepatoxicity in mice: involvement of ER stress, inflammation and oxidative stress.Int Immunopharmacol2018;
64:131–139.
27.
KokBLesterELWLeeWM, et al.
Acute liver failure from tumor necrosis factor-α antagonists: report of four cases and literature review.Dig Dis Sci2018;
63:1654–1666.
28.
JulienOWellsJA.Caspases and their substrates.Cell Death Differ2017;
24:1380–1389.
29.
SongJMaSJLuoJH, et al.
Melatonin induces the apoptosis and inhibits the proliferation of human gastric cancer cells via blockade of the AKT/MDM2 pathway.Oncol Rep2018;
39:1975–1983.
30.
TanMWuALiaoN, et al.
Inhibiting ROS-TFE3-dependent autophagy enhances the therapeutic response to metformin in breast cancer.Free Radic Res2018;
52:872–886.
31.
ChenLLiuLLiY, et al.
Melatonin increases human cervical cancer HeLa cells apoptosis induced by cisplatin via inhibition of JNK/Parkin/mitophagy axis.In Vitro Cell Dev Biol Anim2018;
54:1–10.
32.
MengXZhuYTaoL, et al.
Overexpression of septin-7 inhibits melatonin-induced cell apoptosis in human fetal osteoblastic cells via suppression of endoplasmic reticulum stress. Mol Med Rep2018;
17:4817–4822.
33.
Alonso-GonzalezCMenendez-MenendezJGonzalez-GonzalezA, et al.
Melatonin enhances the apoptotic effects and modulates the changes in gene expression induced by docetaxel in MCF7 human breast cancer cells.Int J Oncol2018;
52:560–570.
34.
Carbajo-PescadorSSteinmetzCKashyapA, et al.
Melatonin induces transcriptional regulation of Bim by FoxO3a in HepG2 cells.Br J Cancer2013;
108:442–449.
35.
FanLSunGMaT, et al.
Melatonin overcomes apoptosis resistance in human hepatocellular carcinoma by targeting survivin and XIAP.J Pineal Res2013;
55:174–183.
36.
LahiriSSinghPSinghS, et al.
Melatonin protects against experimental reflux esophagitis.J Pineal Res2009;
46:207–213.
37.
FuYHeCJJiPY, et al.
Effects of melatonin on the proliferation and apoptosis of sheep granulosa cells under thermal stress.Int J Mol Sci2014;
15:21090–21104.
38.
RequintinaPJOxenkrugGF.Effect of luzindole and other melatonin receptor antagonists on iron- and lipopolysaccharide-induced lipid peroxidation in vitro.Ann N Y Acad Sci2007;
1122:289–294.
39.
MathesAMWolfBRensingH.Melatonin receptor antagonist luzindole is a powerful radical scavenger in vitro.J Pineal Res2008;
45:337–338.
40.
JiaMJingYAiQ, et al.
Potential role of catalase in mice with lipopolysaccharide/d-galactosamine-induced fulminant liver injury.Hepatol Res2014;
44:1151–1158.
41.
MatsuoYIrieKKiyonariH, et al.
The protective role of the transmembrane thioredoxin-related protein TMX in inflammatory liver injury.Antioxid Redox Signal2013;
18:1263–1272.
42.
WenTWuZMLiuY, et al.
Upregulation of heme oxygenase-1 with hemin prevents d-galactosamine and lipopolysaccharide-induced acute hepatic injury in rats.Toxicology2007;
237:184–193.
43.
OkuyamaHNakamuraHShimaharaY, et al.
Overexpression of thioredoxin prevents acute hepatitis caused by thioacetamide or lipopolysaccharide in mice.Hepatology2003;
37:1015–1025.
44.
ItoKOzasaHNodaY, et al.
Effects of free radical scavenger on acute liver injury induced by d-galactosamine and lipopolysaccharide in rats.Hepatol Res2008;
38:194–201.
45.
WeiLRenFZhangX, et al.
Oxidative stress promotes d-GalN/LPS-induced acute hepatotoxicity by increasing glycogen synthase kinase 3β activity.Inflamm Res2014;
63:485–494.
46.
XiaXSuCFuJ, et al.
Role of α-lipoic acid in LPS/d-GalN induced fulminant hepatic failure in mice: studies on oxidative stress, inflammation and apoptosis.Int Immunopharmacol2014;
22:293–302.