
Abstract
Murine NK cell Ly49 receptors, functionally analogous to KIRs in humans recognize MHC class I molecules and play a key role in controlling NK cell function. We have previously shown that the paired activating Ly49s4 and inhibitory Ly49i4 receptors recognize undefined non-classical MHC-Ib ligands from the RT1-CE region in rats. Here, the RT1-CE16 gene of the RT1d haplotype was stably transfected into the mouse RAW macrophage cell line, termed RAW-CE16d cells. Combining RAW-CE16d cells with Ly49 expressing reporter cells demonstrated Ly49i4 and Ly49s4 specificity for CE16d. The Ly49s4/i4:CE16d interaction was confirmed by specific MHC-I blocking monoclonal Abs. Further, we used our in vitro model to study the effect of Listeria monocytogenes (LM) on CE16d after infection. LM infection and IFN-γ stimulation both led to enhanced CE16d expression on the surface of transfected RAW-CE16d cells. Interestingly, the reporter cells displayed increased response to LM-infected RAW-CE16d cells compared with IFN-γ-treated RAW-CE16d cells, suggesting a fundamental difference between these stimuli in supporting enhanced Ly49 recognition of CE16d. Collectively, our data show that Ly49s4 and Ly49i4 recognize the non-classical RT1-CE16d molecule, which in turn is up-regulated during LM infection and thereby may contribute to NK-mediated responses against infected cells.
Keywords
Introduction
Listeria monocytogenes (LM) is a Gram-positive facultative intracellular bacterium enable to cross the intestinal barrier, disseminate via the bloodstream and invade the liver, spleen, central nervous system and the fetus. 1 LM may cause sepsis and meningitis in immune-compromised individuals and severe fetal infections in pregnant women. 2 The bacterium primarily infects epithelial cells and macrophages, and has a unique intracellular life cycle that allows spreading from cell to cell without exposure to the extra-cellular environment. 2 Therefore, any immune response aiming to combat the disease requires destruction of infected cells. Since LM-infected macrophages display altered expression of MHC molecules and also present LM peptides on their MHC molecules, they can both serve as Ag-presenting cells and targets for cytotoxic lymphocytes with receptors monitoring altered MHC-I expression. 3 During primary infection, innate immune cells including neutrophils, NK cells and activated macrophages contribute to the early control of LM infection. Cytotoxic CD8+ T cells are required for clearance and ultimately elimination of the pathogen. 4
NK cells represent an arm of the innate immune system that is able to respond immediately by distinguishing between normal and abnormal cells. 5 They use a sophisticated repertoire of inhibitory and activating receptors that control their activation, proliferation and effector functions. Activation of NK cells occurs when there is a net excess of stimulatory over inhibitory signals. 6 The killer cell Ig-like receptors (KIRs) in humans and the Ly49 receptors in rodents are structurally distinct but functionally similar in that they both recognize classical and non-classical MHC-I molecules.7,8 In the rat, the activating Ly49 receptors recognize ligands within the non-classical MHC-I (termed RT1 in rats). The physiological functions of the non-classical class Ib molecules remain mostly elusive. 9 They typically have a limited tissue distribution and low cell surface expression. Non-classical class I molecules in the rat are encoded in the RT1-CE/N/M region located telomeric of the class II/III regions on chromosome 20, with mostly undiscovered functions. 10 This region is orthologous to the one encoding H2-D, L, Q/T/M molecules in the mouse and similar to HLA-E in humans. The most centromeric class Ib cluster, termed RT1-CE, harbors 13 functional genes in the Brown Norway (BN) rat. 11 The contents of the class I RT1-CE cluster vary considerably between different rat haplotypes.9,12
NK cells play an important role against viral infection. 6 Viruses have evolved multiple mechanisms to evade NK cell receptors by expression of homologue ligand proteins or modulating the NK cell ligands including MHC-Ia and -Ib molecules.13,14 For instance, HIV counteracts stimulation of NK cells by up-regulating surface expression of the non-classical HLA-E molecule while down-regulating classical MHC-I molecules. 15 There is a limited understanding of NK cell responses against bacterial infection including LM.16,17 To investigate the contribution of NK cells in the defense against the LM bacterium, we previously established an in vitro infection model to study the interaction between Ly49 receptors and RT1 molecules. We demonstrated that the RT1-CE molecules were up-regulated in response to LM infection.18,19 Further, the up-regulation of non-classical RT1-CE encoded ligand(s) of haplotype d (RT1-CEd) in LM-infected cells caused stimulation of the paired Ly49s4 and Ly49i4 receptors. 18 Hence, we postulated that induction of stronger recognition by the activating Ly49 receptors was simply due the up-regulation of RT1-CE ligands.18,19 The CE region of haplotype d probably contains 16 functional encoding genes and it is not known whether all these ligands can be recognized by Ly49i4 and Ly49s4 receptors.
In the current investigation, we explored our in vitro pathogen infection model to study Ly49 receptor engagement with a single non-classical Ib molecule influenced by the LM bacterium. By expressing a non-classical class I gene from the CE region of RT1d, termed CE16d, in the heterologous mouse RAW macrophage cell line, our experiments demonstrate that both LM infection and IFN-γ stimulation induced an enhanced CE16d expression on RAW-CE16d cells. However, the interactions of Ly49s4 and Ly49i4 with the CE16d molecule were significantly enhanced against LM-infected cells compared with IFN-γ stimulated cells. These findings suggest a possible post-translational difference in CE16d under these two circumstances, which may consequently influence the interaction of Ly49 receptors with non-classical MHC class I molecules.
Materials and methods
Bacteria and infection
Freeze-dried LM (strain L 242/73 type 4 b, a kind gift from Arja De Klerk, National Institute of Public Health and the Environment, Bilthoven, The Netherlands) was used for in vitro infection. Briefly, cells were infected at a MOI of 1:5 and incubated at 37°C for 1 h, followed by washing with RPMI and re-suspension in 10 µg/ml gentamicin-containing complete RPMI (cRPMI; RPMI 1640 supplemented with 10% FCS, 5 × 10–5 M 2-ME,
Generation of FLAG-tagged CE16d (RAW-CE16d) with RAW264.7 cells
An expression construct consisting of the full-length rat CE16 open reading frame followed by a C-terminal FLAG-tag was generated in pEMCV-SR. RAW264.7 cells (ATCC) were transfected by using FuGENE HD transfection reagent (Roche), followed by sub-cloning and selection in 400 μl cRPMI supplemented with 1 mg/ml G418 sulfate (PAA Laboratories). Cells were kept at 37°C and 5% CO2 and medium replaced on d 2 with fresh medium. Cells were expanded and surface expression of the CE16 molecule analyzed by flow cytometry. RAW-CE16 cells were cultured in Puromycin (InvivoGen) selection medium (0.5 mg/ml).
mAbs and flow cytometry
Panel of mAbs used in blocking experiments or flow cytometry. The specificity and subclasses of the Abs are shown.

BWZ reporter assay
The generation of the Ly49s4 reporter cell has been previously described.
18
In short, the BWZ.36 cell line, kindly provided by N. Shastri (University of California, Berkeley, CA) contains a LacZ reporter construct under transcription control by a 3 × NFAT1 promoter.
12
The cells were stably transfected with the activating Ly49s4 receptor and FLAG-tagged human DAP12. The efficiency of transfection and permanent expression was routinely monitored by flow cytometric analysis. Cells were maintained in cRPMI supplemented with 1 µg/ml Puromycin and 0.5 mg/ml Hygromycin B (all from Invitrogen). Triplicates of target cells were plated at 5.0 × 104 cells/well. BWZ reporter cells were then added at 5.0 × 104 cells/well in cRPMI supplemented with 10 ng/ml PMA (phorbol 12-myristate 13-acetate that activates protein kinase C) and incubated for 18 h. In blocking experiments, mAbs against MHC-I molecules were added. Plates were incubated overnight at 37°C and β-galactosidase activity was assessed by incubating of cells with 150 µM chlorophenol red-β-
BWN reporter assay
The generation of the Ly49i4 reporter cell has been previously described. 18 In short, the BWN reporter cell line is derived from BW5147 T cells stably transfected with EGFP under the control of NFAT1. 13 The BWN cells were stably transfected with the Ly49i4-CD3ζ chimeric receptor, created by fusing the inverted intracellular domain of the CD3ζ-chain with the Ly49 transmembrane and extra-cellular ligand-binding domain of the Ly49i4 receptor. Cells were maintained in cRPMI supplemented with 1 mg/ml G418 sulfate and 0.5 mg/ml Hygromycin B and expression routinely monitored. Similar to with the BWZ reporter, 10 ng/ml PMA was added to BWN-Ly49i4 reporter cells before mixing with target cells. After 18 h of incubation, cells were analyzed for EGFP expression by flow cytometry. Dead cells were excluded by PI labeling.
Statistical analysis
All data are presented as mean ± SEM and were analyzed by paired or unpaired Student’s t-test or one-way ANOVA with Bonferroni correction or by Students t-test, using Prism software (GraphPad). The significance level was set at P < 0.05.
Results
Generation of RAW-CE16d cells
The complete genomic sequence of the BN rat strain MHC (n haplotype) is available.
11
The first cluster of class I genes of the non-classical RT1-CE/N/M region, the CE region, contains up to 16 genes, depending on haplotype. We used the BN genomic sequence to design primers for cloning of RT1-CE16d from the d haplotype,
18
which in turn was cloned into FLAG-tagged expression vectors and transfected into the RAW mouse cell line. The success of stable transfection and expression of CE16d in RAW cells was determined by FACS analysis (Figure 1). The recombinant CE16d molecule was low to moderately expressed on RAW-CE16d cells as judged by specific mAb staining with anti-rat classical and non-classical MHC-Ia,b (OX18), anti-non-classical MHC-Ib (AAS5) and anti-FLAG (Figure 1). The weak expression of transfected CE16d on RAW cells was in line with the typical low expression level of non-classical molecules.18,19
Expression of rat CE16d on CE16d-transfected mouse RAW cells. Mouse RAW cells (upper panel), or RAW cells transfected to express rat CE16d (RAW-CE16d, lower panel) were surface stained with the following mAbs specific for rat (solid lines); anti-FLAG (M2, left panel), anti-MHC-I (classical MHC-Ia and non-classical MHC-Ib, OX18, middle panel), or anti-MHC-Ib (AAS5, right panel) for flow cytometric analysis. Negative secondary Ab control is depicted in stippled line. Shown are representative plots from four independent experiments.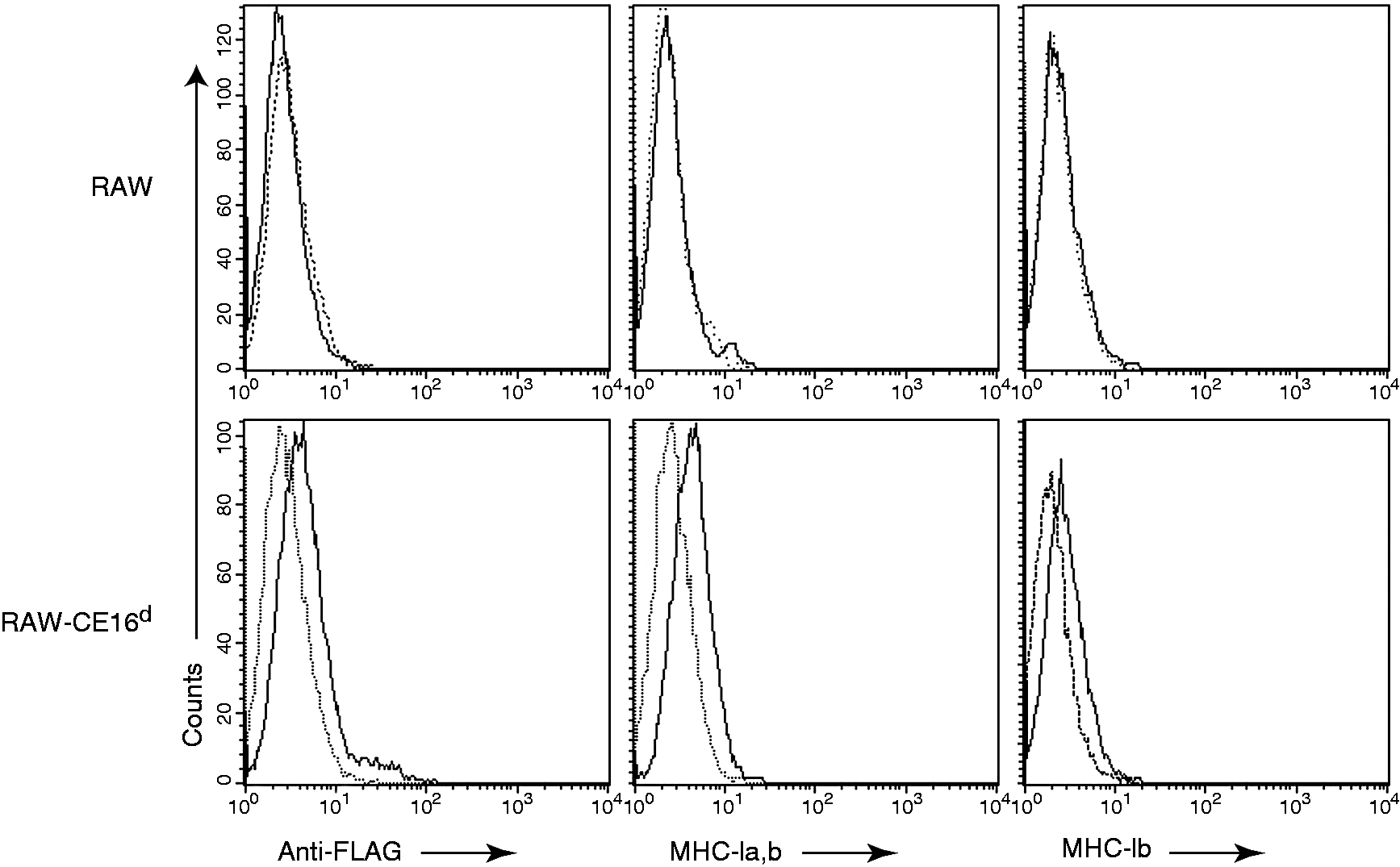
LM infection and IFN-γ stimulation enhances expression of CE16d on RAW-CE16d cells
We have previously established optimal conditions for infection of various cell types with LM.18,19 RAW cells were heavily LM infected but still viable after 48 h post infection and employed for CE16d transfection. We examined the influence of LM infection and IFN-γ on CE16d expression by the transfected CE16d cells. RAW-CE16d cells were infected with LM. In parallel, we stimulated the RAW-CE16d cells with 100 U/ml IFN-γ for CE16d expression. We observed that both LM-infected and IFN-γ-stimulated RAW-CE16d cells displayed increased expression of CE16d molecules, as revealed by FACS staining with anti-FLAG (M2, left plot) and anti-MHC-Ib (AAS1, middle plot and AAS5, right plot) mAbs (Figure 2a and b). These experiments demonstrate that the CE16d molecule is IFN-γ-inducible and the expression is altered in response to LM infection.
LM infection and IFN-γ stimulation enhance expression of CE16d on RAW-CE16d cells. (a) Representative histogram overlays of LM-infected (gray shaded), IFN-γ-stimulated (solid thick line) and untreated (solid thin line) RAW-CE16d cells. Negative secondary Ab control is denoted (stippled line). Staining with anti-FLAG (left panel) and the anti-MHC-Ib mAbs AAS1 (middle panel) or AAS5 (right panel) are presented. (b) Summary data show geometric mean (gMFI) ± SEM of RAW-CE16d cells treated as described above versus untreated controls, stained with AAS1 (left panel) and AAS5 (right panel). The plots and values shown are representative of four independent experiments. P < 0.05 with Student’s t-test.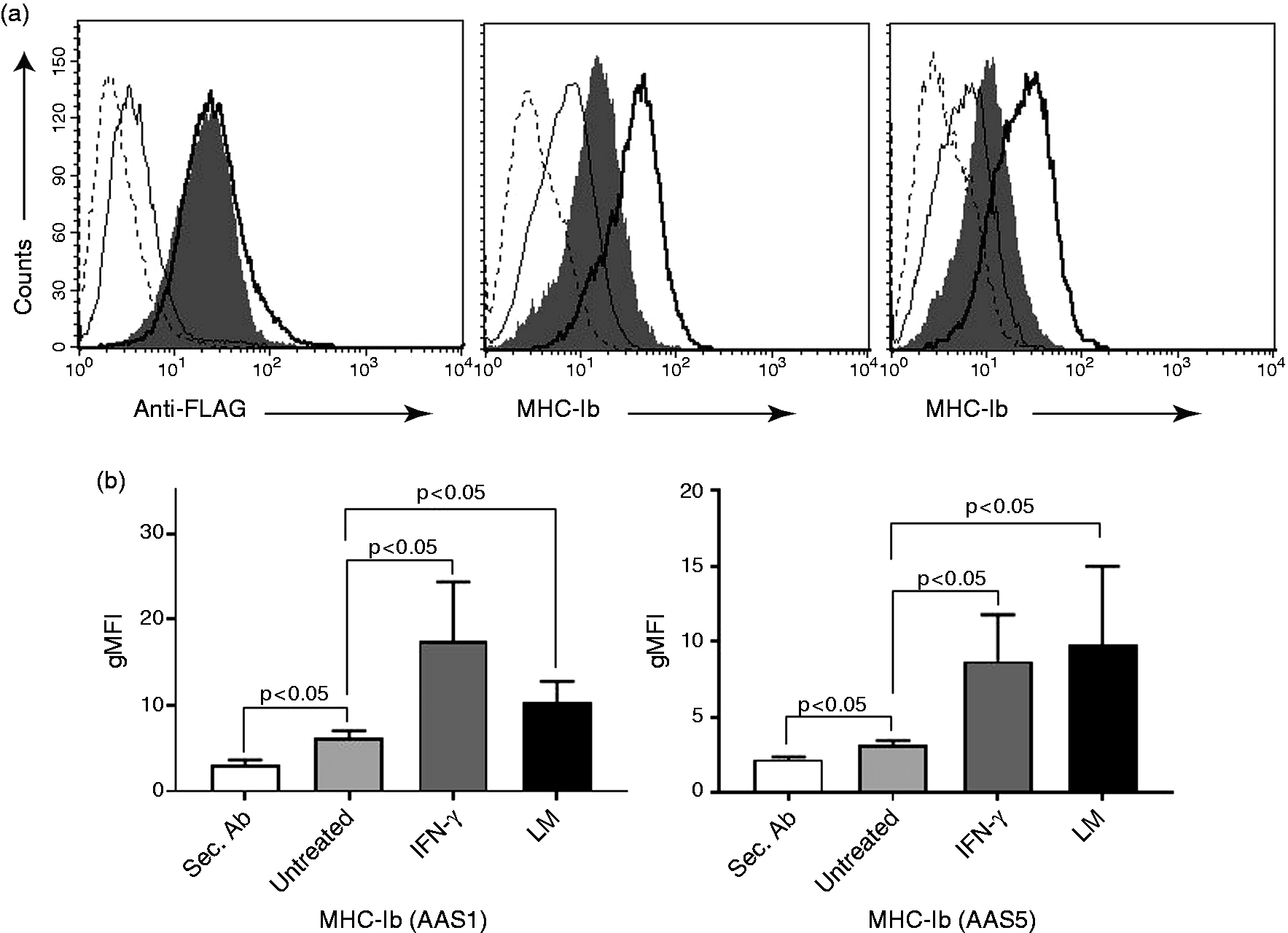
Ly49s4 recognizes the non-classical class I molecule CE16d, and mediates a stronger response to CE16d upon LM infection
We have previously shown that the activating Ly49s4 receptor recognizes ligand(s) encoded within the non-classical region of the RT1-CEd haplotype.
18
It is unknown if the Ly49s4 receptor is able to recognize all molecules encoded within the CEd region, and whether these ligands behave similarly during infection. To address whether the CE16d molecule is specifically recognized by Ly49s4 receptor, we utilized the BWZ reporter cell expressing Ly49s4 and the signaling adapter molecule, DAP12. Untransfected RAW cells, and RAW-CE16d cells either untreated, IFN-γ stimulated or LM-infected, were incubated with the Ly49s4 reporter cells for 18 h. The Ly49s4 reporter cells did not elicit any response against untransfected RAW cells, either untreated, IFN-γ stimulated or LM-infected, compared with an Ionomycin-treated positive control (Figure 3a). In contrast, the Ly49s4 reporter cells responded to the untreated and uninfected RAW-CE16d transfectant, indicating that Ly49s4 recognizes the CE16d molecule. This recognition was significantly enhanced for LM-infected CE16d expressing cells, but not with IFN-γ treated RAW-CE16d cells (Figure 3a).
Ly49s4 recognizes the non-classical class I molecule CE16d and mediates a stronger response to CE16d upon LM infection. (a) BWZ-Ly49s4 reporter cells were incubated with untransfected RAW cells (all in beige bars) or RAW-CE16d cells; uninfected (white bars), treated with IFN-γ (light gray bars)or infected with LM (black bars). For the blocking experiments, cell cultures were incubated with 3 µg/well of purified mAbs, or 10 µl of Ab-containing hybridoma supernatants. Stimulation of reporter cells with Ionomycin served as a positive control (dark gray bar). Responses were measured in β-galactosidase activity. All experiments were performed in triplicates and quantitative data of five independent experiments are presented. P < 0.5, P < 0.01 with Student’s t-test. (b) A representative visualization of BWZ-Ly49s4 reporter cell responses against RAW-CE16d displaying color development in microwells is shown.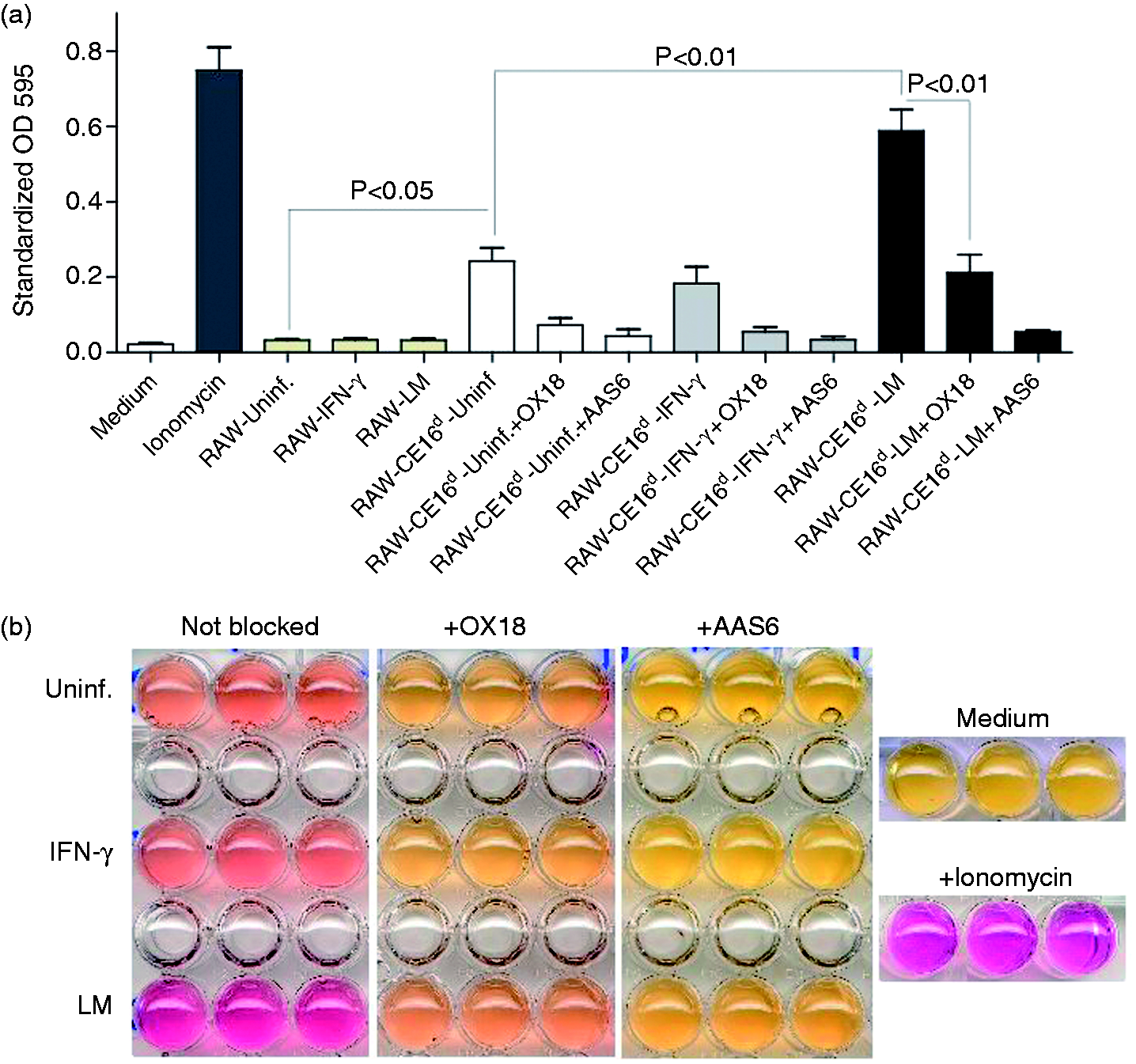
The increased activation of Ly49s4 reporter cells by LM-infected RAW-CE16d cells was clearly concomitant with the higher CE16d expression (Figure 2a,b). Interestingly, IFN-γ treatment of RAW-CE16d cells, which also induced up-regulation of CE16d expression and to a level that exceeded that as a result of LM infection, did not induce a similar increase in activation of Ly49s4 reporter cells (Figure 3a). The activation of reporter cells was substantially if not completely blocked, depending on the experimental group, verified with the mAbs OX18 [(which binds to MHC-Ia,b molecules), or AAS1 (not depicted) and completely with AAS6 (Figure 3a,b). All of these Abs have in common the ability to recognize rat MHC-Ib molecules, with recognition by AAS1 and AAS6 limited exclusively to rat MHC-Ib molecules in haplotype d. The blocking by these mAbs indicates that the responses were indeed mediated by the rat CE16d molecule expressed by the transfected RAW cells. A representative experiment is shown in Figure 3b. Together, the data demonstrate that CE16d is a ligand for the Ly49s4 receptor. The results suggest that the Ly49s4 receptor more effectively recognizes CE16d in the context of LM infection than from IFN-γ stimulation.
To exclude that stimulation of reporter cells was not due to cytokines released by the LM-infected cells, and to confirm the specificity of the response, we used the Ly49s5 reporter cells that recognize ligands of the RT1-Eu haplotype.19,22 These reporter cells differ only in expression of the activating Ly49 receptor. RAW-CE16d cells were therefore co-incubated with Ly49s4 or Ly49s5 reporter cells. Regardless of IFN-γ treatment or LM infection of the RAW CE16d-expressing cells, Ly49s5 reporter cells did not respond to RAW-CE16d cells compared with Ly49s4 reporter cells (Figure 4a). The Ly49s5 reporter cells did respond to Ionomycin treatment, indicating that they are capable of responding, ruling out a signaling deficiency as an explanation for its lack of responsiveness to CE16d. In addition, unlike live LM, heat-killed LM (HkLM) did neither enhance the CE16d expression (not shown) nor reporter cell stimulation (Figure 4b). Collectively, these data reinforce the specific interaction between Ly49s4 and CE16d, and exclude the stimulation of reporter cells due to cytokines that may be released by LM-infected RAW cells. Furthermore, experiments with HkLM demonstrate that Ly49 recognition of CE16d is dependent on the viability of infecting LM.
Ly49s4, but not Ly49s5 recognizes CE16d, and Ly49s4 recognition of CE16d is enhanced with live but not heat-killed LM treatment of CE16d expressing cells. (a) A representative experiment of RAW-CE16d cells in triplicates co-incubated with Ly49s4 reporter cells (white bars) or Ly49s5 reporter cells (black bars). Stimulation with Ionomycin (positive control), uninfected (negative control), IFN-ɣ treatment and LM infection are indicated and identical to those in 3 a. (b) A representative experiment of BWZ-Ly49s4 reporter cells in triplicates with heat-killed HkLM treated RAW-CE16d cells (gray bars) compared with uninfected (white bars), IFN-γ-treated (light gray bars) or live LM-infected (black bars) is shown. Data shown are representative of three independent experiments.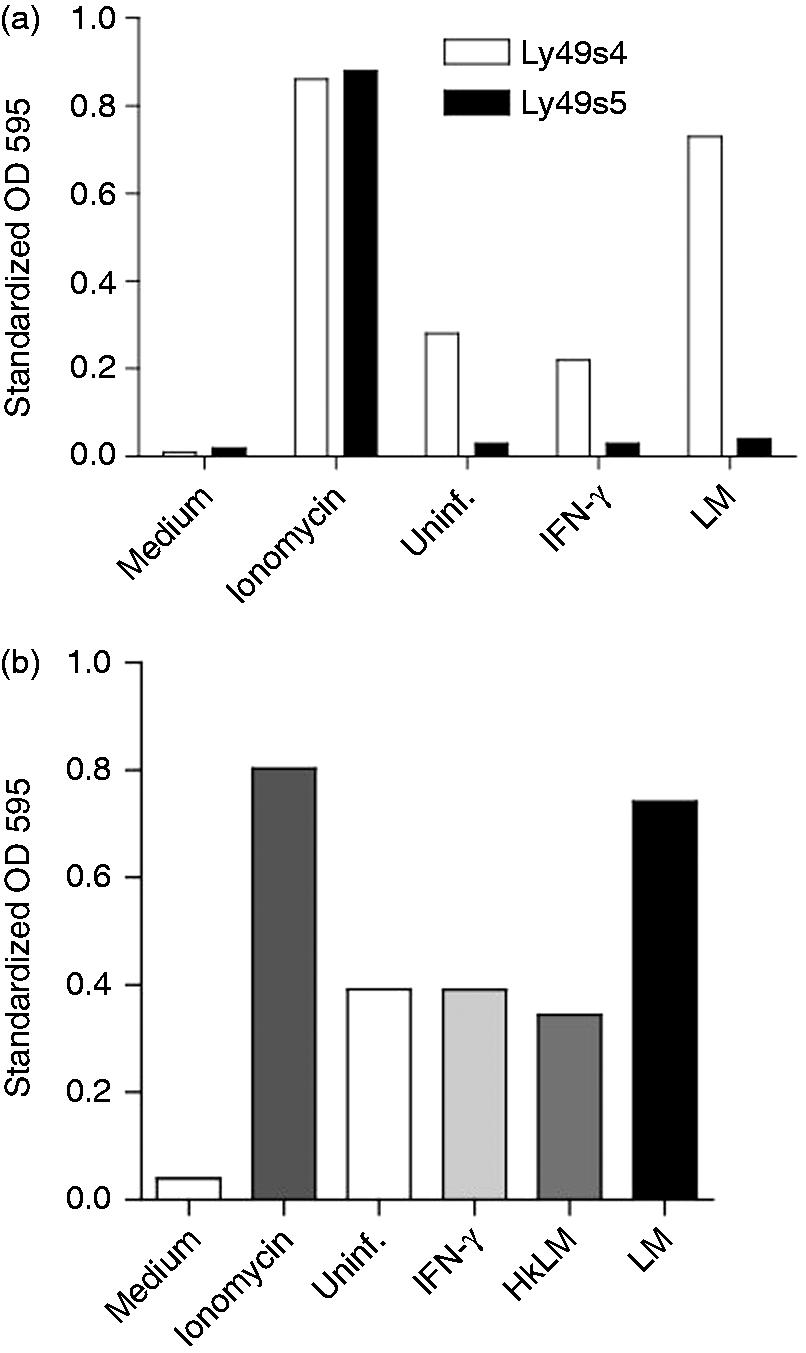
The ectodomain of the Ly49i4 inhibitory receptor recognizes CE16d
We next asked whether the inhibitory receptor Ly49i4, like Ly49s4, is capable of recognizing the CE16d molecule. To approach this, we utilized the BWN-Ly49i4 reporter cell that expresses the transmembrane and ligand-binding domain of Ly49i4, fused to the inverted intracellular domain of the CD3ζ-chain. Recognition of a Ly49i4 ligand will subsequently induce EGFP expression by the reporter cells. The reporter cells are capable of a strong signaling response, as treatment with Ionomycin resulted in a majority of the reporter cells expressing EGFP (Figure 5a, upper row). Reporter cells expressing chimeric Ly49i4-CD3ζ, incubated with RAW-CE16d cells, produced EGFP development, although modest, it still indicated recognition of CE16d (second row). The low Ly49i4 reporter cell response, compared with Ly49s4 reporter cells, was in line with our previous results with R2 cells (expressing all RT1d molecules).
18
Subsequently, this recognition could be reduced in the presence of anti-MHC-I (OX18) or Abs specifically targeting non-classical MHC-Ib molecules (AAS1 and AAS6) (Figure 5a and b). Similar to Ly49s4, recognition of CE16d by the Ly49i4 ligand recognition domain was not enhanced by IFN-γ (third row) or HkLM (fourth row) treatment of the RAW CE16d-expressing cells (Figure 5a). Infection with LM of the RAW-CE16d cells more than doubled the reporter cell response to CE16d (Figure 5a, fifth row), suggesting that LM infection enhances recognition of CE16d by Ly49i4. The specificity of the LM-augmented recognition is shown with MHC-I specific blocking of the reporter cell response, and similar to results with Ly49s4 reporter cells, HkLM did not enhance Ly49i4 reporter cell stimulation (Figure 5a and b).
The ectodomain of the Ly49i4 inhibitory receptor recognizes CE16d. (a) Dot plots of the Ly49i4-CD3ζ chimera-expressing reporter cells treated with medium (negative control) or Ionomycin (positive control) (upper row), co-incubated with RAW-CE16d cells (first panel), that were uninfected (second row), treated with IFN-γ (third row), exposed to HkLM (fourth row) or infected with live LM (fifth row). Parallel co-incubations performed with the indicated MHC-specific blocking Abs to define the ligand specificity of the response included: 10 µl of Ab-containing hybridoma supernatants OX18 (second panel column), 3 µg/well of purified AAS1 (third panel column) and 3 µg/well of purified AAS6 (fourth panel column). Responses were measured in EGFP fluorescence. The plots and values shown are representative of four independent experiments. (b) Statistical significance between Ly49i4 reporter cells in co-culture with CE16d expressing cells uninfected (white bars), treated with IFN-γ (light gray bars), infected with heat-killed LM (dark gray bars) or live LM (black bars) was determined with a two-tailed unpaired Student’s t-test (P < 0.02). Data, summarized from four independent experiments, represent means ± SEM of standardized reporter cell EGFP expression.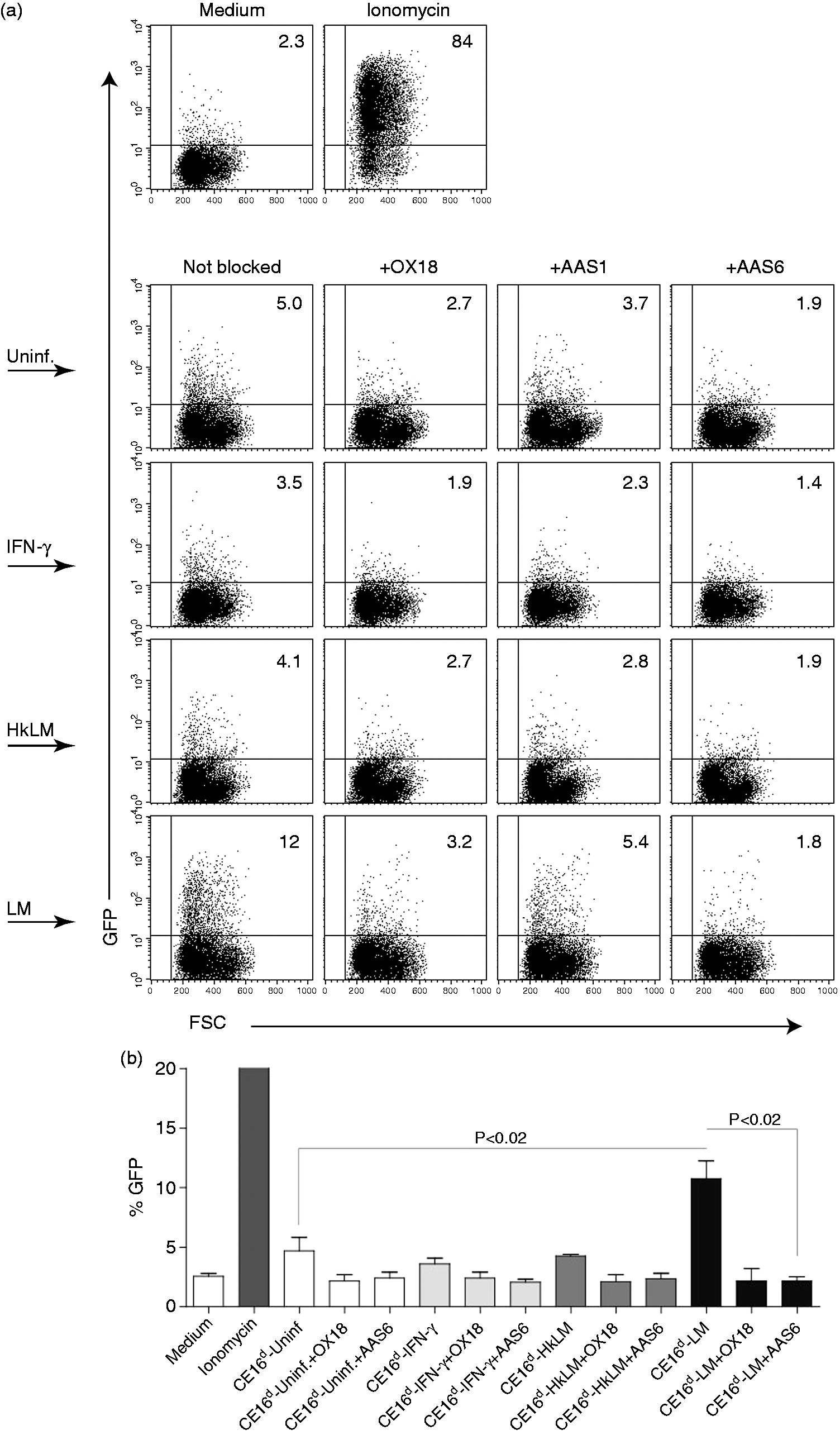
Taken together, the preceding results demonstrate that the CE16d molecule is recognized by the paired Ly49s4 and Ly49i4 receptors. Although both IFN-γ treatment and LM infection of the RAW-CE16d cells similarly up-regulated the CE16d, Ly49s4 and Ly49i4 reporter cell responses were significantly enhanced only by LM infection. Finally, the HkLM experiments indicate that infection with live LM is necessary for the augmentation of CE16d recognition by Ly49s4 and Ly49i4 ectodomain-expressing reporter cells.
Discussion
The RT1 region contains a large and dense area of non-classical Ib genes with mostly unknown function. 10 We have previously shown the important role of Ly49:RT1-CE in early defense against LM.18-20,23 Here, we report that the last gene in this region, CE16d, is a ligand for the paired Ly49s4 and Ly49i4 receptors and CE16d expression is as a consequence of LM infection. 18 More importantly, only LM-induced (but not IFN-γ-induced) enhancement of CE16d expression was concomitant with increased recognition by both activating Ly49s4 and inhibitory Ly49i4 receptors. Up-regulation of CE16d and its recognition, particularly by the activating Ly49s4, may explain NK cell protection in response to LM infection. Given that the responses were largely if not completely due to recognition of CE16d, as evidenced by our MHC-I-specific Ab blocking experiments, our observations imply that the CE16d expressed on LM-infected cells differs from that on IFN-γ-treated cells and is more efficiently recognized by Ly49 receptors. There may be post-translational differences between CE16d expressed on LM-infected cells compared with CE16d expressed on IFN-γ-treated cells. The most likely possible difference is in the repertoire of peptides bound to CE16d in the two circumstances, although the peptide-binding characteristics of CE16d are presently not known.
Ly49 receptors often do not show exquisite peptide specificity, but peptides may stabilize the MHC-I molecule that is important for sufficient surface expression. 24 However, some Ly49 receptors, including mouse Ly49C 25 and Ly49I 26 and rat Ly49i2, 27 may be peptide selective in their recognition of MHC-I-peptide complexes. This implies that some MHC-I-bound peptides, but not all, support receptor recognition. Consistent with these possibilities, our results open the possibility for contemplation that LM-derived peptides bound to CE16d consequently enhance Ly49:CE16d recognition. An alternative, or an additional, influence is that LM infection alters the repertoire of host cell-derived peptides bound by CE16d, mediating enhanced Ly49 recognition.
It remains unknown whether all CE molecules are similarly affected by LM infection in terms of expression and interaction with Ly49 receptors. In vitro data show that LM may influence expression of MHC-I in different ways. While some cell lines up-regulate their MHC-I in response to LM,18,19 the MHC-I remained unaffected on other cell types.34 The various MHC-I alterations may be haplotype and tissue dependent. HIV up-regulates HLA-E, while down-regulating classical MHC-I. 15 A thought-provoking speculation could be that LM non-uniformly modulates the genes located in the RT1A and -CE regions. As demonstrated by crystallography studies, Ly49 recognition of MHC-I molecules occurs at a site underneath the peptide-binding groove, 24 a region that can be affected by peptide anchor residues, possibly influencing the topology of the MHC-I site for Ly49 association. In fact, Ly49 recognition can be determined/influenced by peptide residues bound in specific buried locations in the peptide-binding groove, and this may be a fundamental feature of Ly49 recognition of MHC-I ligands. 28 Our results also indicate that whatever mechanism enhances Ly49 recognition of CE16d, it is dependent on the viability of infecting LM, as HkLM did not enhance recognition by Ly49s4 and Ly49i4. It may be that the LM or its products do not gain access to relevant intracellular compartments to affect CE16d unless the LM is alive. Alternatively, only live LM alters the cellular machinery that favors changes in CE16d composition, such as bound peptides, and/or expression resulting in enhanced recognition by Ly49 receptors.
It is not known whether Ly49s4 and Ly49i4 are expressed on the same NK cells or on different NK cell subsets. This distinction has clearly been shown for another rat Ly49 receptor pair, Ly49s5 and Ly49i5, that are expressed on distinct NK cell subpopulations. 22 In the case of expression on separate NK cell subsets, enhanced Ly49s4 engagement may overcome inhibitory receptors other than Ly49i4 on the Ly49s4+ subset. By contrast, if Ly49s4 and i4 are expressed on the same NK cell subset, then it is conceivable that CE16d-bound peptides play a role. This variable in the context of LM infection could be important in determining the relative engagement of the activating and inhibitory Ly49s4 and i4, favoring Ly49s4, albeit relative receptor densities and signaling strength might also play a role. Insight into the relative ability of Ly49s4 and i4 to engage CE16d molecules is difficult to obtain using receptor reporter systems, and in our case two different reporter systems were used for the activating and inhibitory receptors, respectively. Future studies using surface Plasmon resonance, 29 and other techniques, may clarify differences in ligand preference and possibly affinity, and this would be particularly relevant if these two receptors are found to be expressed on the same NK cell subset(s).
The existence of activating and inhibitory Ly49 receptors capable of binding a common or related ligand during microbe infection, as found here with LM-infected cells, has been observed earlier, during MCMV infection. In that case, the MCMV-encoded m157 MHC decoy binds inhibitory Ly49C and I receptors, and to the activating Ly49H receptor.30,31 The MCMV expresses m157 to evade NK cell destruction of cells infected with the virus, by engaging the inhibitory receptors. 30 However, Ly49H, expressed on NK cells of the C57BL/6 mouse strain, by engaging m157 and activating the Ly49H+ NK cell subset, offers protection from MCMV. 30 By analogy, the LM bacterium may attempt to protect LM-infected cells from NK cell destruction by attempting to enhance recognition by the inhibitory Ly49i4 receptor, but Ly49s4 recognition may be more effective and trigger NK cell responses that offer protection from LM. A major and distinctive difference in LM infection compared with MCMV infection is that the ligand recognized by the activating and inhibitory Ly49 receptors is a host-expressed non-classical molecule, instead of a pathogen-encoded ligand.
In conclusion, our data provide direct evidence for a specific non-classical MHC-I, CE16d, serving as a ligand for the paired activating Ly49s4 and inhibitory Ly49i4 receptors, and these interactions can be modified substantially by LM infection of host cells. Ly49:MHC-I interaction in an infection model presented here may facilitate further research to gain knowledge into the complex mechanism(s) of NK cell-mediated resistance to bacterial infections.
Footnotes
Acknowledgments
Generous technical support of the people in the Bent Rolstad’s lab is gratefully acknowledged. This work was supported by grants from the Norwegian Cancer Society, the Research Council of Norway, Anders Jahre’s Foundation for the promotion of Science and the University of Oslo. The authors regretfully acknowledge that B. R. is deceased. This article is dedicated to the memory of B. R.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.