
Abstract
Heat-killed (HK) Mycobacterium obuense is a novel immunomodulator, currently undergoing clinical evaluation as an immunotherapeutic agent in the treatment of cancer. Here, we examined the effect of in vitro exposure to HK M. obuense on the expression of different categories of surface receptors on human blood myeloid (m) and plasmacytoid (p) DCs. Moreover, we have characterized the cytokine and chemokine secretion patterns of purified total blood DCs stimulated with HK M. obuense. HK M. obuense significantly up-regulated the expression of CD11c, CD80, CD83, CD86, CD274 and MHC class II in whole-blood mDCs and CD80, CD123 and MHC class II in whole-blood pDCs. Down-regulation of CD195 expression in both DC subpopulations was also noted. Further analysis showed that HK M. obuense up-regulated the expression of CD80, CD83 and MHC class II on purified blood DC subpopulations. TLR2 and TLR1 were also identified to be engaged in mediating the HK M. obuense-induced up-regulation of surface receptor expression on whole blood mDCs. In addition, our data demonstrated that HK M. obuense augmented the secretion of CCL4, CCL5, CCL22, CXCL8, IL-6, IL-12p40 and TNF-α by purified total blood DCs. Taken together, our data suggest that HK M. obuense exerts potent differential immunomodulatory effects on human DC subpopulations.
Introduction
Dendritic cells (DCs) are crucial modulators of the immune response whereby they act as a bridge between the innate and adaptive immune systems. 1 DCs are potent APCs that are in continuous surveillance for Ags while being in an immature state. Upon exposure to an Ag (self/foreign) or to a pro-inflammatory stimulus, DCs undergo a maturation process and migrate towards secondary lymphoid organs where they interact with T cells, consequently inducing an Ag-specific immune response or initiating immune tolerance.2,3 The hallmarks of DC maturation include up-regulation of co-stimulatory (CD80, CD86), Ag presentation (MHC class I and II) and maturation (CD83) receptors in addition to the release of pro-inflammatory cytokines (IL-6, IL-12).4,5 To date, DCs are broadly categorized into two phenotypically and functionally distinct subpopulations defined as the conventional or myeloid (m) and plasmacytoid (p) DCs. Both subpopulations exist in peripheral blood and they comprise <1% of PBMCs.6,7
The vital requirement for an effective immunotherapeutic approach for the treatment of various diseases has resulted in the development of novel strategies which exploit the biological properties of DCs.8,9 DCs are equipped with a specialized set of PRRs which identify specific microbe or damage associated molecular patterns (MAMPs or DAMPs). 10 Therefore, this unique and pivotal characteristic of DCs makes them a potential target for bacteria-based immunomodulatory agents which, upon binding to PRRs on DCs, trigger a chain of events that leads to transcriptional activation, alterations in DC cytokine/chemokine release, and a variation in surface receptor expression, thus affecting the interaction between DCs and other immune cells.11,12 Among the bacteria-based immunomodulators, heat-killed (HK) Mycobacterium vaccae preparation has demonstrated immunotherapeutic and vaccine potential when evaluated in different disease settings.13–17 In recent years, there has been a growing interest in assessing additional HK mycobacterial preparations, as potential immunotherapeutic agents, particularly in cancer. HK Mycobacterium obuense (NCTC13365) has been shown to be safe and well tolerated among patients with stage III/IV melanoma. 18 A recent randomized phase II study has demonstrated that the use of HK M. obuense as an adjunctive immunotherapeutic agent for treating advanced pancreatic cancer was well tolerated and led to a significant extension in the overall and progression-free survival of patients with metastatic disease. 19 Research into the mode of action of HK M. vaccae and M. obuense has demonstrated their abilities to directly affect the innate arm of the immune system. In a study in allergic mice, M. vaccae was reported to promote the development of CD11c+ APCs demonstrating a possible regulatory role. 20 Evidence from in vitro studies on human blood cells clearly indicated that both HK M. vaccae and M. obuense are able to modulate the expression of various cell surface receptors on monocytes and neutrophils, 21 and to stimulate cytokine release from type-1 mDCs, thus leading to the enhancement of the anti-tumor activity of γδ T cells. 22 Moreover, priming of monocyte-derived (Mo) DCs with M. vaccae was reported to correlate with a significant attenuation of Th type 2 (Th2) responses. 23
Despite the emerging clinical interest in HK M. obuense, little is known about the outcome of its interaction with human innate immune cells, particularly primary blood DCs. The purpose of our study was to analyze the effect of HK M. obuense stimulation on the expression of different categories of surface receptors on whole blood and purified blood DCs and to evaluate whether TLRs are involved in the M. obuense-induced up-regulation of surface receptor expression on whole-blood mDCs. In addition, we examined the secretion of cytokines and chemokines by purified total blood DCs stimulated with HK M. obuense. Our results showed that HK M. obuense differentially regulates the expression of surface receptors related to Ag presentation, co-stimulation and maturation on blood DCs. Furthermore, our data revealed that TLR2 and TLR1 contribute to the M. obuense-induced up-regulation of CD11c, CD80 and MHC class II expression on whole blood mDCs. Finally, HK M. obuense demonstrated an ability to induce cytokine and chemokine production by purified total blood DCs.
Materials and methods
Blood sampling
Peripheral whole-blood samples were drawn at the clinical laboratory department, Nini Hospital, Lebanon, from healthy donors and collected into K2-EDTA vacutainer tubes (BD Biosciences, San Jose, CA, USA). Written informed consent was obtained from all blood donors and the study procedure gained approval from the institutional review board at the University of Balamand and from the research ethics committee at the Faculty of Science, Engineering and Computing at Kingston University.
TLR blocking Abs and reagents
Anti-human monoclonal blocking Abs against TLR1 (clone H2G2), TLR2 (clone B4H2), TLR4 (clone W7C11), TLR5 (clone Q2G4), TLR6 (clone C5C8) and matching isotype control mouse IgG1 (clone T8E5) and human IgA2 (clone T9C6) Abs were purchased from Invivogen (Toulouse, France). Sterile vials of HK M. obuense (NCTC13365) suspended in borate-buffered saline (BBS; pH 8.0) at 50 mg/ml were manufactured by BioElpida (Lyon, France), whereby each 1 mg wet mass of bacilli is roughly equivalent to 109 HK organisms.
Immunophenotyping of DCs
Whole-blood cultures (total culture volume = 2 ml) were either left unstimulated or were stimulated with 30, 100 or 300 µg/ml of HK M. obuense for 3 and 6 h at 37℃ in a 5% CO2 humidified atmosphere. For purified total blood DCs, viable cells were re-suspended at 105 cells/ml in complete RPMI 1640 medium (Lonza, Slough, UK) supplemented with 10% heat-inactivated human AB serum and were stimulated with M. obuense for 6 h, as mentioned above. Whole-blood cultures or purified blood DCs incubated with equivalent amounts of BBS served as unstimulated control. Both mDC and pDC subpopulations were analyzed in whole blood and as purified cells for the expression of a group of cell-surface receptors. Whole blood (100 μl) was incubated for 25 min at room temperature (RT) in the dark with optimized concentrations of Ag-specific or matching isotype control monoclonal Abs (Supplementary Table S1) and red blood cells were then lysed by the addition of FACS lysing solution (BD Biosciences). After 15 min of incubation at RT, cells were pelleted and washed once with Cell Wash solution (BD Biosciences) and finally resuspended in Cell Fix solution (BD Biosciences). For the immunophenotyping of purified blood DCs, a similar procedure was adopted; however, the incubation and centrifugation steps were carried out at 4℃ and a Fc receptor blocking step with 10% heat-inactivated human AB serum (Lonza) was introduced (15 min incubation period) prior to incubating cells with Abs. Fixed cells were run on a FACSCalibur flow cytometer (BD Biosciences) and the generated data were acquired and analyzed using Cell Quest Pro software (BD Biosciences). A total of 2.5–3.0 × 105 leukocytes or 5 × 103 purified total blood DCs were acquired per sample. Single-color-stained controls were used to set compensation settings. Total blood DCs were identified as Lineage–/HLA-DR+, whereas blood mDCs and pDCs were identified as Lineage–/HLA-DR+/CD11c+/CD123− and Lin–/HLA-DR+/CD123+/CD11c–, respectively. The expression of each cell-surface receptor was indicated as the percentage of cells positive or a particular surface receptor within a gated cell population or as the raw geometric mean fluorescence intensity (MFI) of the receptor-positive cell population.
Isolation of CD4+ T cells and blood DCs
PBMCs were isolated from peripheral blood using the standard Ficoll-Paque gradient method, as previously described. 21 CD4+ T cells and total blood DCs were purified from PBMCs by negative selection using the Dynabeads untouched human CD4 T cells and human DC enrichment kits, respectively, and following the manufacturer's instructions (Thermo Fisher Scientific, Waltham, MA, USA). For the depletion of platelets, biotin-conjugated mouse anti-human CD41 Ab (clone M148; Leinco Technologies, St. Louis, MO, USA) was added to the Ab cocktail present in the DC enrichment kit. Isolated cells were checked for purity by flow cytometry and were consistently found to be >95% and ∼80% for CD4+ T cells and DCs, respectively. Cells were routinely analyzed for viability with trypan blue exclusion method and showed >90% viability.
Ag presentation assay
Freshly purified total blood DCs were seeded in a 96-well, flat-bottom plate at a density of 1 × 104 viable cells/well in complete RPMI medium supplemented with 10% heat-inactivated human AB serum and were either left unstimulated or were stimulated with 300 µg/ml of HK M. obuense in the presence or absence of 10 µg/ml Candida albicans soluble Ag (Greer Laboratories, Lenoir, NC, USA), for 7 d at 37 ℃ in a 5% CO2 humidified atmosphere. After 1 d in culture, DCs were co-cultured with 1 × 105 purified autologous CD4+ T cells to obtain a T-cell/DC ratio of 10:1. Co-cultures were maintained for a period of 6 d at 37 ℃ in a 5% CO2 humidified atmosphere. Cells were pulsed with 1 μCi/well tritiated thymidine (Perkin Elmer, San Jose, CA, USA) during the last 16 h of the co-culture period. Cells were then harvested onto glass-fiber filter disks (Connectorate AG, Dietikon, Switzerland) using a cell harvester (Inotech Biotechnologies, Basel, Switzerland). Proliferation of CD4+ T cells was assessed by measuring the radioactivity of incorporated methyl-tritiated-thymidine in a liquid scintillation counter (Perkin Elmer), obtained as cpm. Unstimulated (containing equivalent amounts of BBS as vehicle) DCs co-cultured with CD4+ T cells, as well as CD4+ T cells, without DCs, stimulated with HK M. obuense plus C. albicans served as negative controls. Stimulation index was calculated by dividing the mean cpm value of DC–CD4+ T cell co-cultures (triplicate) stimulated with HK M. obuense and/or C. albicans over the mean cpm value of unstimulated DC-CD4+ T cell co-cultures (triplicate).
TLR blocking
Whole blood was incubated with 10 µg/ml of cell culture grade, azide-free blocking monoclonal Abs against TLR1, TLR2, TLR4, TLR5 and TLR6 or isotype control Abs (mentioned above) for 1 h prior to stimulating whole blood with 300 µg/ml of HK M. obuense for 3 h. The efficiency of TLR blocking Abs has been previously validated. 21
Detection of chemokine and cytokine production
Purified total blood DCs (5 × 104 cells/ml) were either left unstimulated or were stimulated with 300 µg/ml of HK M. obuense for 24 h at 37 ℃ in a 5% CO2 humidified atmosphere. DC culture supernatants were collected after 24 h and were stored at −20℃ until further analysis. Levels of CCL4, CCL5, CCL22, CXCL8, IFN-α, IL-6, IL-10, IL-12p40 and TNF-α were determined in DC culture supernatants using commercially available sandwich ELISA kits (R&D Systems, Abingdon, UK) and performed according to the manufacturer’s protocol.
Statistical analysis
Statistical analysis of the data was carried out using GraphPad Prism software (version 6; GraphPad Software, San Diego, CA, USA). Data are presented as mean values ± SEM. Statistical differences were determined by paired t-test, Wilcoxon matched-pairs signed rank test or one-way ANOVA test followed by the Dunn’s post-hoc test and were considered to be statistically significant at P < 0.05.
Results
HK M. obuense modulates the expression of different surface receptors on whole blood mDCs and pDCs
Flow cytometric analysis of DC subpopulations in whole blood was done initially by gating viable leukocytes and excluding debris and dead cells on the basis of their FSC/SSC characteristics at 3, 6 and 24 h. Within this gate, total DCs were recognized as Lineage–/HLA-DR+ and the mDC and pDC subpopulations were further defined in the gated DC population as Lineage–/HLA-DR+/CD11c+/CD123– and Lineage–/HLA-DR+/CD123+/CD11c–, respectively (Figure 1a). Preliminary experiments over a 3-, 6- and 24-h time course indicated that blood DCs tend to lose their phenotype as Lineage–/HLA-DR+ cells in 24-h unstimulated and M. obuense-stimulated whole-blood cultures (Figure 1b). We performed a dose–response analysis to assess the effect of whole-blood stimulation with HK M. obuense for 3 and 6 h on the surface expression of MHC class II on blood DCs. HK M. obuense up-regulated the surface expression of MHC class II on blood DCs in a dose-dependent manner and this increase was apparently more prominent in mDCs than pDCs (Figure 1c). Based on these results, 300 µg/ml of HK M. obuense was selected as an optimal concentration for regulating surface receptor expression in blood DCs and was employed in subsequent experiments.
(a) Gating scheme for identification of mDCs and pDCs in whole blood. Viable leukocytes were gated based on forward and side scatter (FSC and SSC, respectively) properties. Among the gated leukocyte population, DCs were identified as Lineage–/HLA-DR+ and within this DC population, mDC and pDC subpopulations were identified as Lineage–/HLA-DR+/CD11c+/CD123– and Lineage–/HLA-DR+/CD123+/CD11c–, respectively. (b) Loss of flow cytometric profile of total DCs (Lineage–/HLA-DR+) in 24-h whole-blood cultures. Total DCs were identified in whole-blood cultures that were either left unstimulated or were stimulated with 300 µg/ml of HK M. obuense for 3, 6 and 24 h. Flow cytometry graphs are representative of three independent experiments. (c) Dose–response assessment for the effect of whole-blood stimulation with HK M. obuense for 3 and 6 h on the expression level of MHC class II on mDCs and pDCs. Whole blood was stimulated with various concentrations of HK M. obuense (30, 100 and 300 µg/ml) for 3 and 6 h and the expression level of MHC class II, presented as geometric MFI of MHC class II+ DCs, was measured on mDC and pDC subpopulations. Symbols and error bars indicate mean values and SEM, respectively (n = 4). Statistically significant differences were determined by one-way ANOVA test followed by the Dunn’s post-hoc test (*P < 0.05 vs. unstimulated).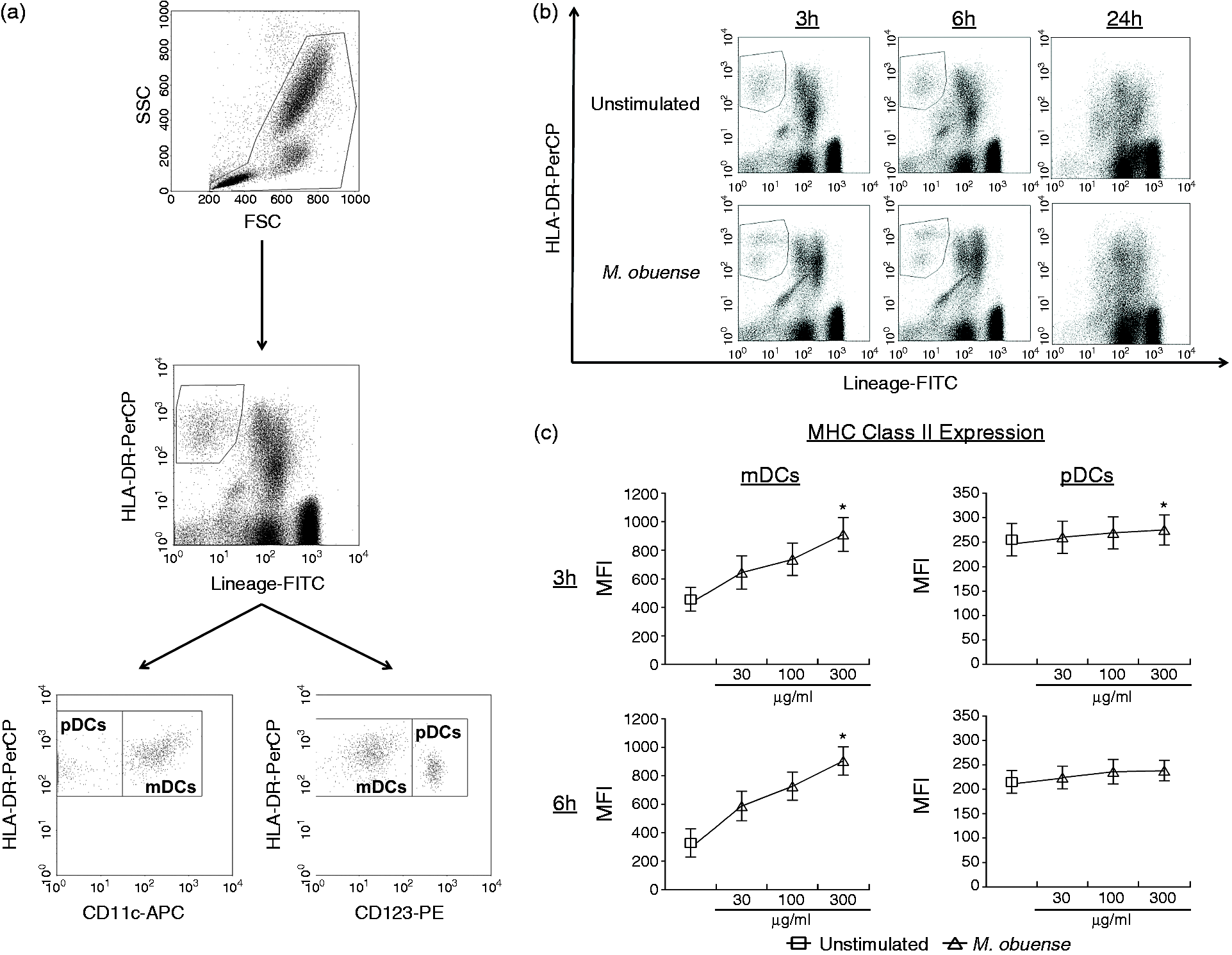
In order to determine whether in vitro whole-blood stimulation with HK M. obuense would influence the phenotype of whole-blood mDC and pDC subpopulations, we measured the expression level of different categories of surface receptors on both DC subpopulations following a 3- and/or 6-h stimulation period. Representative flow cytometry figures of the percentage or MFI of receptor-positive mDCs are shown in Figure S1. Among the Ag presentation receptors, only the MFI of MHC class II+ cells was significantly elevated (P < 0.05) on mDCs with a 1.75- and ∼2.5-fold change at 3 and 6 h after stimulation with HK M. obuense, respectively (Figure 2). mDCs responded to stimulation with HK M. obuense by significantly upregulating (P < 0.05) the expression of the B7 family receptors where the percentages of CD80+ and CD86+ mDCs were significantly increased 7-fold and 2-fold, respectively, after 3 h of stimulation as well as 38-fold and 2.5-fold, respectively, after 6 h of stimulation. However, a significant increase in the MFIs of CD80+ and CD86+ mDCs was noted only after 6 h of mycobacterial stimulation (Figure 2). The percentage and MFI of CD274+ mDCs were significantly induced (P < 0.05) after 3 h of stimulation, and these were even more evident at the 6 h time point (Figure 2). The expression of complement receptors was significantly enhanced on mDCs following stimulation with HK M. obuense. A significant up-regulation (P < 0.05) in the percentage of CD11b+ mDCs and in the MFI of CD11c+ mDCs was observed at 3 h and at both incubation time points after M. obuense stimulation, respectively (Figure 2). Analysis of the expression of cytokine and chemokine receptors revealed that in response to HK M. obuense stimulation, the MFI of CD116+ mDCs and the percentage of CD253+ mDCs were significantly upregulated (P < 0.05) at 3 and 6 h, respectively (Figure 2). However, the expression of CD195 was significantly (P < 0.05) down-regulated in mDCs at 3 and 6 h. Members of the PRRs demonstrated enhanced expression on mDCs after stimulation with HK M. obuense. The percentages of CD282+ and CD284+ mDCs were significantly enhanced at both stimulation time points, whereas only the percentage of CD206+ mDCs and the MFI of CD282+ mDCs were significantly enhanced at the 3-h time point (Figure 2). Whole-blood stimulation with HK M. obuense for 3 and 6 h also resulted in a marked increase (P < 0.05) in the expression of the DC maturation marker, CD83, on mDCs (Figure 2).
Whole-blood stimulation with HK M. obuense modulates surface expression of different receptors on mDCs. Whole-blood cultures were either left unstimulated or were stimulated with 300 µg/ml of HK M. obuense for 3 and 6 h and the expression level of different categories of cell-surface receptors was measured on mDCs. Column bars represent mean values of the percentage (%) and geometric MFI of receptor-positive mDCs of at least three donors. Error bars represent SEM. Statistically significant differences were determined by paired t-test (*P < 0.05 vs. unstimulated). NT: not tested.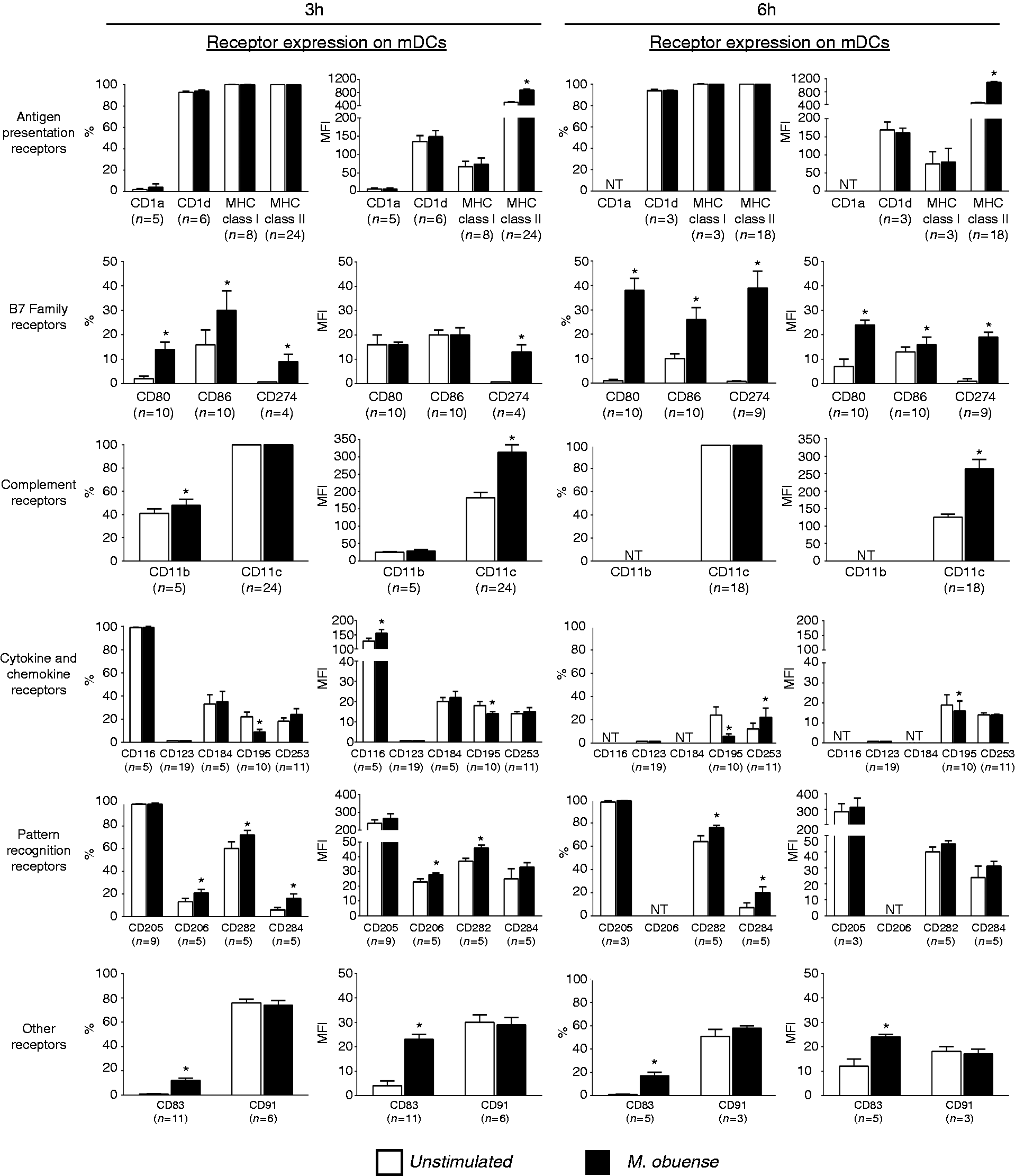
In response to whole-blood stimulation with HK M. obuense, pDCs exhibited a modest but statistically significant regulation in the expression of a restricted number of receptors belonging to the Ag presentation, B7 and cytokine and chemokine families of receptors, whereas no significant regulation in the expression of the studied complement receptor and PRR families was detected (Figure 3). A mild but significant (P < 0.05) increase in the MFI of MHC class II+ pDCs was observed after stimulation with HK M. obuense for 3 and 6 h (Figure 3). The percentage of CD80+ pDCs was slightly but significantly (P < 0.05) upregulated at 6 h after stimulation with HK M. obuense (Figure 3). Moreover, 3 and 6 h M. obuense-stimulated pDCs demonstrated a slight but significant increase in the MFI of CD123+ cells and a significant decrease in the percentage and MFI of CD195+ cells compared with unstimulated pDCs (Figure 3). We could not detect any spontaneous or HK M. obuense-induced expression of CD1d, CD83 CD91, CD282, CD284, and CD274 on pDCs (Figure 3).
Whole-blood stimulation with HK M. obuense modulates surface expression of different receptors on pDCs. Whole-blood cultures were either left unstimulated or were stimulated with 300 µg/ml HK M. obuense for 3 and 6 h and the expression level of different categories of cell-surface receptors was measured on pDCs. Column bars represent mean values of the percentage (%) and geometric MFI of receptor-positive pDCs of at least three donors. Error bars represent SEM. Statistically significant differences were determined by paired t-test (*P < 0.05 vs. unstimulated). NT: not tested.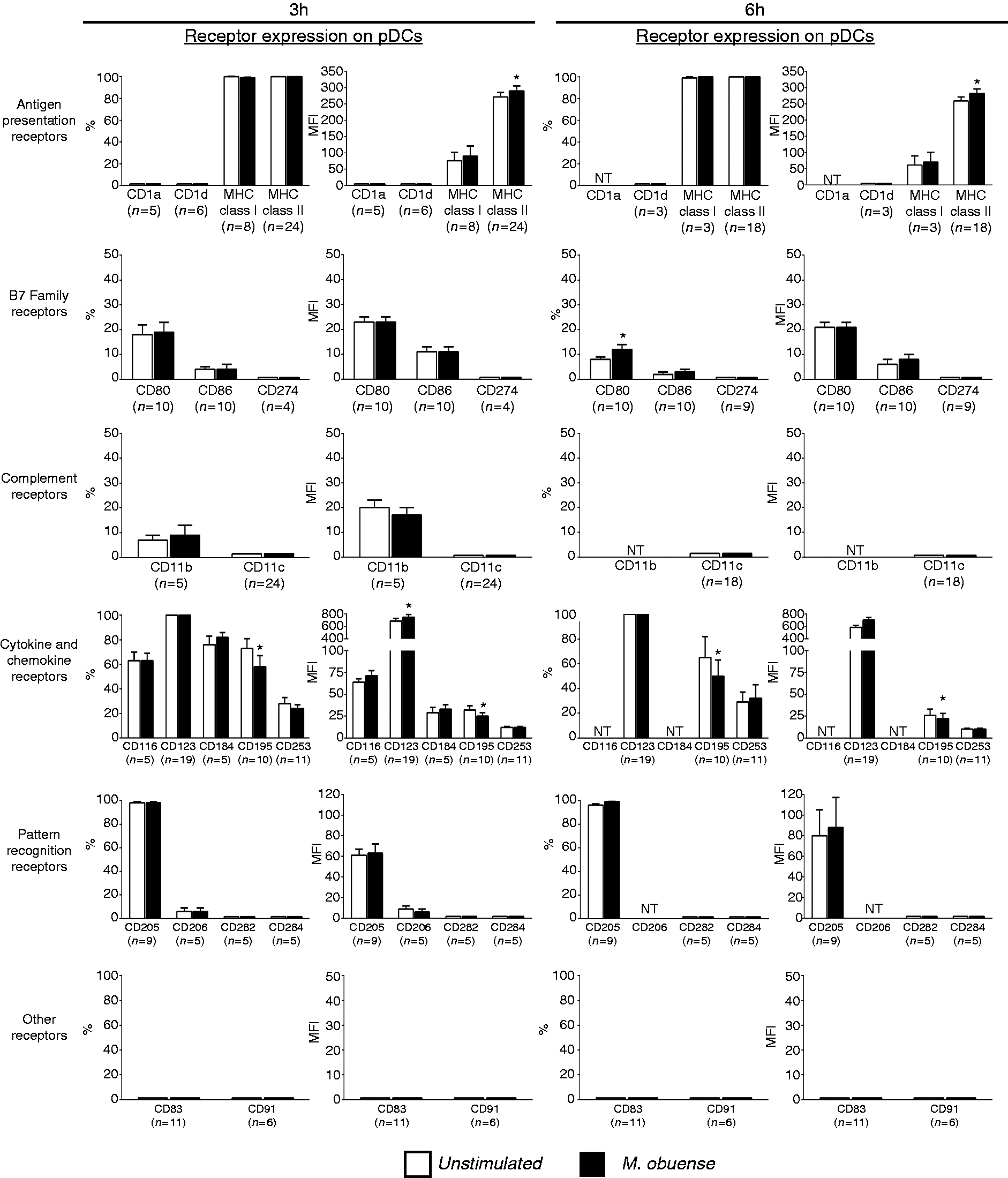
Additional receptors (CD40, CD197, CD209, CD273, CD275 and CD276) were also studied on both DC subpopulations and under similar conditions; however, their expression was neither constitutively expressed nor induced following mycobacterial stimulation. Taken together, our data reveal that whole-blood stimulation with HK M. obuense resulted in significant variations in the expression of receptors on mDC and pDC subpopulations; however, these variations were more prominent on mDCs. Based on the significantly up-regulated expression of CD80, CD83, CD86 and MHC class II and down-regulated expression of CD195 in M. obuense-stimulated mDCs, it can be inferred that HK M. obuense might drive mDCs towards acquiring a mature DC phenotype.
TLR2 and TLR1 blocking reduces the HK M. obuense-induced up-regulation of CD11c, CD80 and MHC class II expression on whole-blood mDCs
We have previously reported that TLR2 and TLR1 contribute to the M. obuense-induced up-regulation of surface receptor expression of CD11c and MHC class II on monocytes in whole blood.
21
We therefore attempted to determine whether TLR2 and TLR1 in addition to other TLRs (TLR4, TLR5 and TLR6) are also involved in mediating the M. obuense-induced expression of surface receptors on whole-blood mDCs. TLR blocking experiments were carried out using specific blocking Abs against TLR1, TLR2, TLR4, TLR5 and TLR6 whereby the effect of TLR blocking on the M. obuense-induced expression of selected receptors was studied. The selected receptors included: CD11c, CD80 and MHC class II, which represented three categories of surface receptors that demonstrated significant variations in expression (% or MFI) on mDCs following whole-blood stimulation with HK M. obuense (Figure 2). Both single and simultaneous blocking of TLR2 and TLR1 led to a significant reduction (P < 0.05) in the M. obuense-induced expression of CD80 (% of CD80+ cells) and MHC class II (MFI of MHC class II+ cells) on mDCs, whereas blocking of TLR2 and TLR2/1 resulted in a significant reduction (P < 0.05) in the M. obuense-induced expression of CD11c (MFI of CD11c+ cells) on mDCs (Figure 4). We did observe a trend of an enhanced blocking effect with combined TLR2/1 blocking vs. separate TLR blocking; however, this effect achieved statistical significance (P < 0.05) only against single TLR2 blocking in the case of MHC class II. Although there was a trend towards a decrease in the M. obuense-induced expression of CD11c and MHC class II in response to TLR1 and TLR2/6 blocking, respectively, this decrease did not attain statistical significance (Figure 4). Simultaneous blocking of TLR2 and TLR6 significantly reversed the M. obuense-induced expression of CD11c and CD80, an effect comparable to that observed with blocking TLR2 alone (Figure 4). Blocking of TLR4, TLR5 and TLR6 resulted in no significant reduction in the M. obuense-induced up-regulation of the three surface receptors (Figure 4).
Blocking of TLR2 and/or TLR1 reduces the M. obuense-induced up-regulation of CD11c, MHC class II and CD80 expression on mDCs in whole blood. Whole-blood cultures were pre-incubated with anti-(α)-TLR1, α-TLR2, α-TLR2/1, α-TLR6, α-TLR2/6, α-TLR4, α-TLR5 or isotype-matched control antibodies: IgA (for α-TLR2 and 5) or IgG (for α-TLR1, 4 and 6) for 1 h and then stimulated with 300 µg/ml HK M. obuense (Mob) for 3 h followed by the analysis of the expression level of CD11c, MHC class II and CD80 on mDCs. Column bars represent mean values of the geometric MFI of CD11c+ and MHC class II+ mDCs or the percentage (%) of CD80+ mDCs of three donors. Error bars represent SEM. Statistically significant differences were determined by Wilcoxon matched-pairs signed rank test (*P < 0.05 vs. Mob).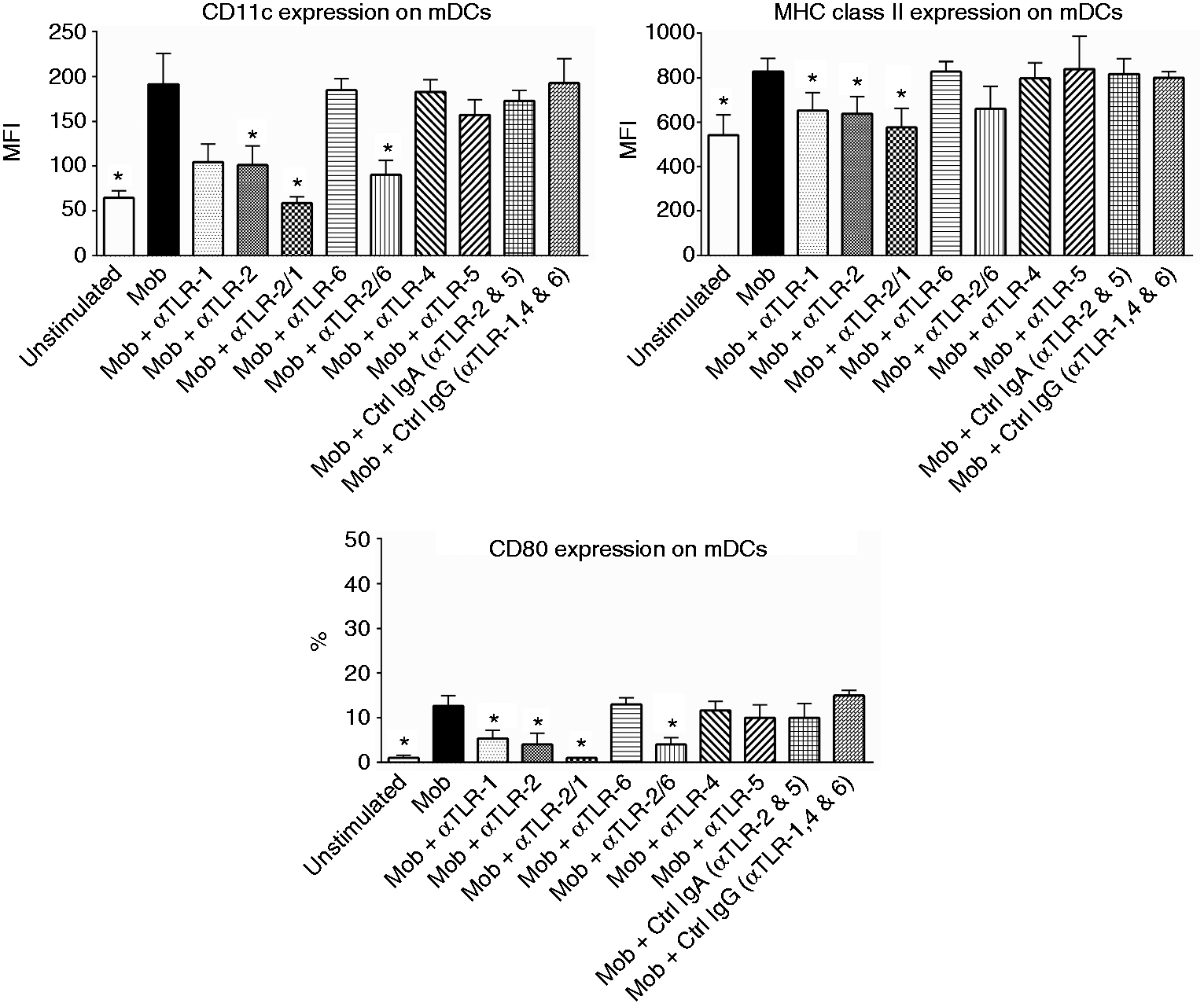
HK M. obuense modulates surface expression of different receptors on purified blood DCs
Our results have shown that whole-blood stimulation with HK M. obuense induced significant variations in the expression of different surface receptors on mDCs and pDCs in whole blood. This finding has prompted us to investigate whether the detected variations in receptor expression were merely related to a direct stimulatory effect induced by HK M. obuense on DCs or due to an indirect stimulatory effect induced by the whole-blood milieu (other blood cells, pro-inflammatory cytokines, etc.). To answer this question, we examined the effect of stimulating purified blood DCs with HK M. obuense on the expression of different receptors that were selected to represent those which showed significant variations on whole-blood DCs upon stimulation with HK M. obuense. Following 6-h stimulation with HK M. obuense, purified blood mDC and pDC subpopulations significantly upregulated (P < 0.05) the MFI of MHC class II+ cells (Figure 5). A significant (P < 0.05) ∼8.5-fold and 16.5-fold increase in the percentage of CD80+ cells was observed on M. obuense-stimulated purified blood mDCs and pDCs, respectively; however, only M. obuense-stimulated purified blood pDCs showed a significant 2.8-fold increase in the MFI of CD80+ cells (Figure 5). Unlike pDCs, M. obuense-stimulated purified blood mDCs revealed a significant elevation in the expression of CD274+ cells. M. obuense-stimulated purified blood mDCs and pDCs demonstrated, respectively, a significant up-regulation (P < 0.05) in the percentage of CD253+ cells and in the MFI of CD123+ cells (Figure 5). Both mDCs and pDCs enhanced the percentage of CD83+ cells in response to stimulation with HK M. obuense (Figure 5).
HK M. obuense modulates surface expression of different receptors on purified DCs. Purified total blood DCs were either left unstimulated or were stimulated with 300 µg/ml of HK M. obuense for 6 h and the expression level of different categories of cell-surface receptors was measured on the mDC and pDC subpopulations. Column bars represent mean values of the percentage (%) and geometric MFI of receptor-positive mDCs and pDCs of five donors. Error bars represent SEM. Statistically significant differences were determined by paired t-test (*P < 0.05 vs. unstimulated).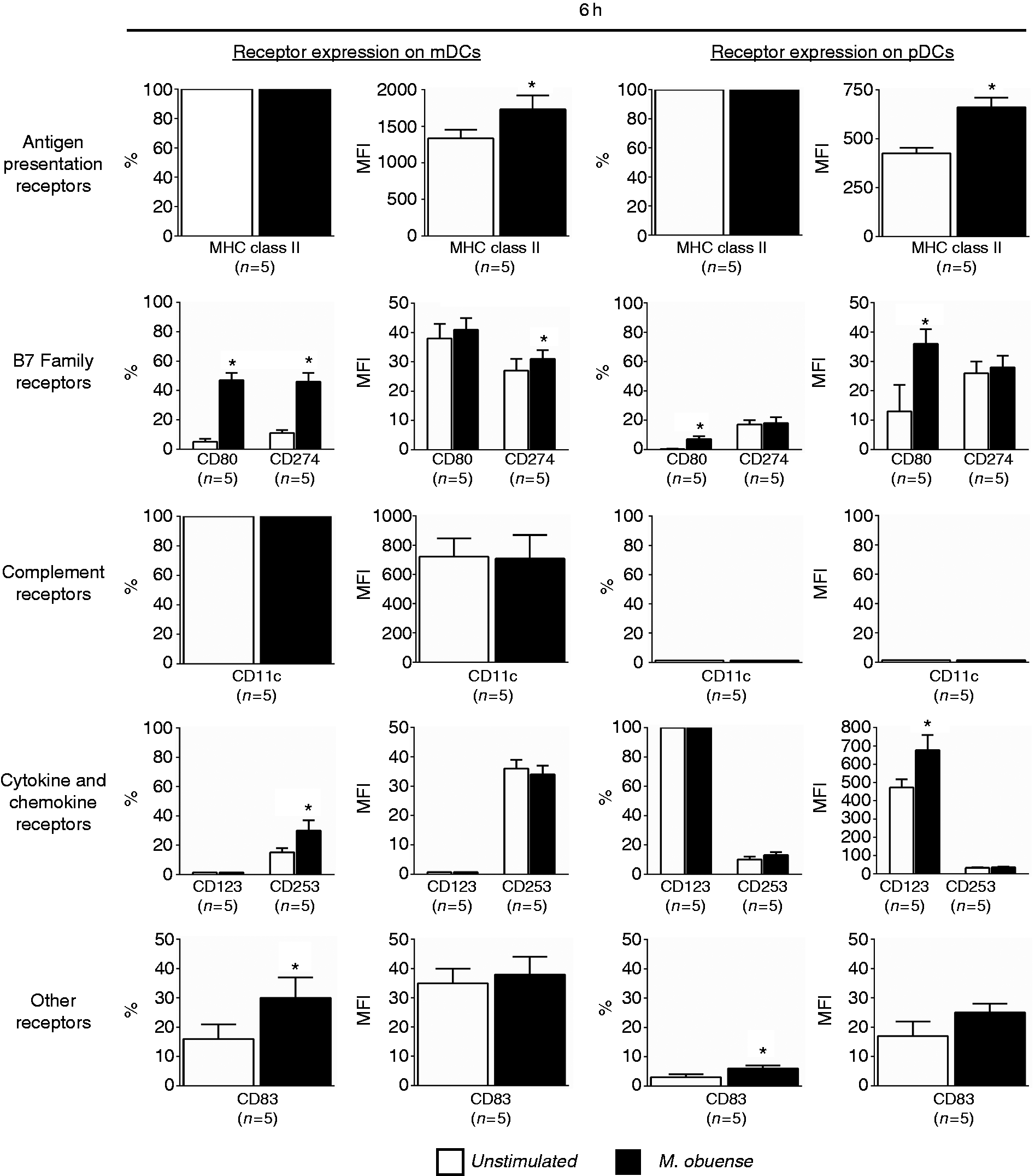
HK M. obuense induces cytokine and chemokine secretion by purified total blood DCs
We next examined the cytokine and chemokine secretion patterns of purified total blood DCs stimulated with HK M. obuense. Stimulation of DCs with HK M. obuense for 24 h was shown to substantially induce the release of CCL22 (1248 ± 324 pg/ml) and CXCL8 (2703 ± 443 pg/ml) (Figure 6). Moreover, M. obuense-stimulated DCs exhibited a significant (P < 0.05) ∼18 - and ∼2-fold increase in the production of CCL4 and CCL5, respectively, compared with unstimulated DCs (Figure 6). Stimulation with HK M. obuense also resulted in a weak, but significant (P < 0.05), secretion of IL-6, IL-12p40 and TNF-α by DCs (Figure 6). However, levels of IFN-α and IL-10 were not detected in supernatants of either unstimulated or M. obuense-stimulated DC cultures.
HK M. obuense alters the secretion of cytokines and chemokines by purified DCs. Purified total blood DCs were either left unstimulated or were stimulated with 300 µg/ml of HK M. obuense for 24 h. Levels of IL-6, IL-10, IL-12p40, TNF-α, IFN-α, CCL4, CCL5, CCL22 and CXCL8 were measured in DC culture supernatants. Column bars represent mean values of cytokine or chemokine concentration in DC culture supernatants of seven donors. Error bars represent SEM. Statistically significant differences were determined by paired t-test (*P < 0.05 vs. unstimulated).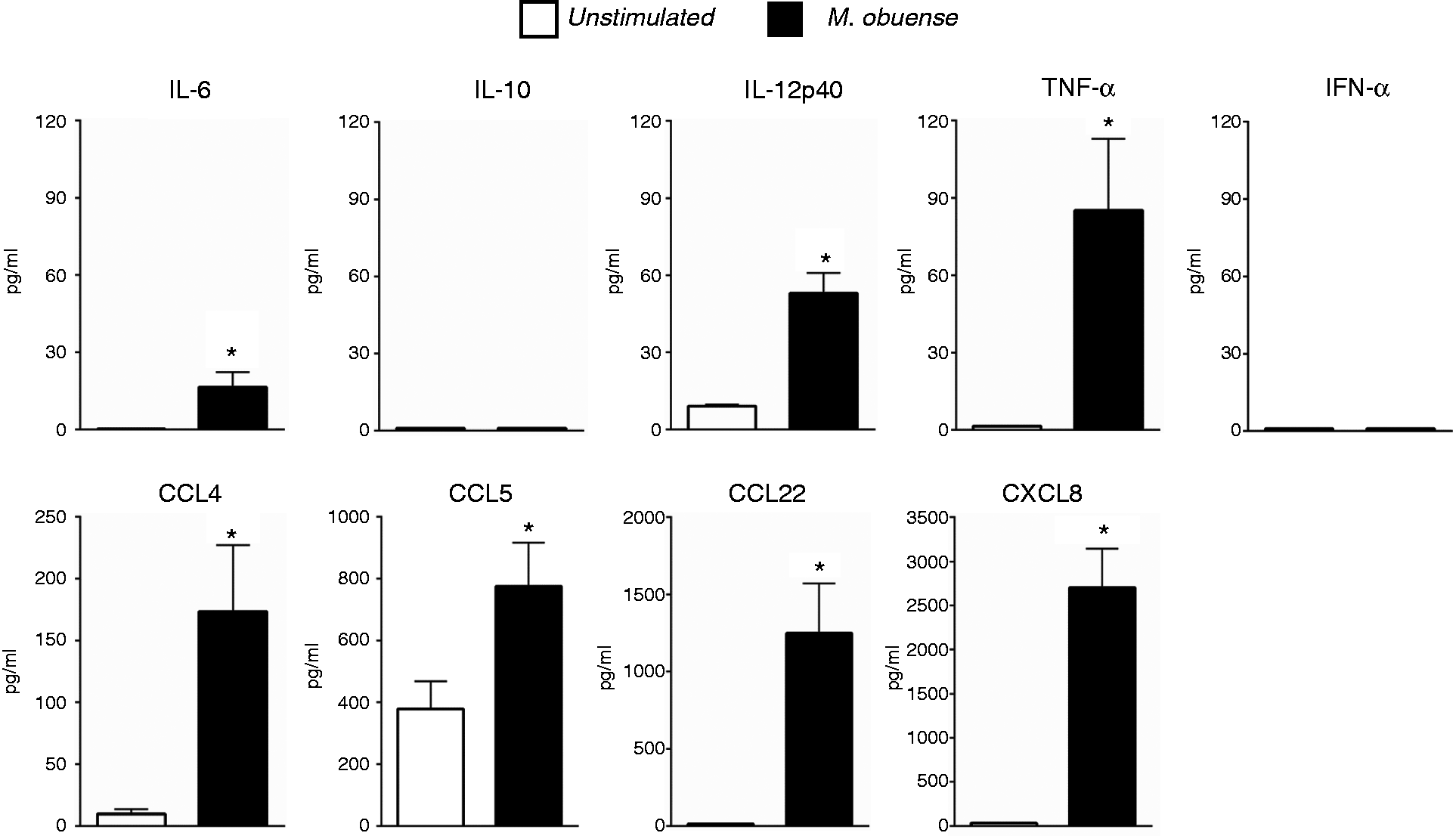
Effect of DC stimulation with HK M. obuense and/or C. albicans on CD4+ T-cell Ag-specific responses
We investigated the ability of purified total blood DCs to present HK M. obuense Ags and/or C. albicans-soluble Ag to autologous purified CD4+ T cells. DCs stimulated with HK M. obuense or C. albicans-induced comparable autologous CD4+ T-cell proliferation with stimulation indexes of 24 ± 14 and 25 ± 14, respectively. However, concomitant stimulation of DCs with HK M. obuense and C. albicans Ags elicited significantly (P < 0.05) higher CD4+ T-cell proliferative responses (stimulation index = 51 ± 26) than DCs stimulated separately with HK M. obuense or with C. albicans alone (Figure S2). Further analysis of our CD4+ T-cell proliferation data clearly demonstrated that the autologous CD4+ T-cell proliferative responses induced by DCs stimulated with HK M. obuense and C. albicans were not statistically different (P > 0.05) from the additive proliferative responses to each Ag separately. As expected, low CD4+ T-cell proliferation was noted when these lymphocytes were stimulated with both Ags, but without the presence of DCs (280 ± 66 cpm), or when co-cultured with DCs, but without the presence of Ags (504 ± 107 cpm).
Discussion
Shedding light on the outcome of interaction of an immunomodulator with immune cells is a vital step towards understanding the immunomodulator’s mode of action. Over the past few years, HK M. obuense, a novel systemic immunomodulator, has attracted the attention of cancer researchers;18,19 however, little is known about the consequences of its interaction with human innate immune cells. In this study, we investigated the effect of HK M. obuense on the phenotype and cytokine/chemokine secretion profile of human blood DCs, which are limited in number, 24 and represent a challenge in the study of human DC biology. To the best of our knowledge, this is the first study to describe the effect of HK M. obuense on human whole blood and purified primary blood DCs.
Our first approach was aimed at examining the effect of in vitro exposure to HK M. obuense on the expression of a group of receptors belonging to six different categories on mDC and pDC subpopulations in a human whole-blood model, which represents a physiologically relevant system that has been used extensively to assess the effect of several immunomodulators on the phenotype, function and responses of blood DCs.25–27 During initial time-course studies, we could not identify DCs either in unstimulated or in M. obuense-stimulated 24-h whole-blood cultures owing to the loss of their unique flow cytometric profile as Lineage–/HLA-DR+ cells. A previous study has also failed to measure intracellular cytokine production by human DC subpopulations in 24-h whole-blood cultures owing to an inability to reliably detect different DC subpopulations. 28 On the contrary, other studies have reported the detection of human DC subpopulations after a 20–24-h culture period of whole blood and were able to assess the phenotype and function of DCs.27,29 The reasons behind this discrepancy with our data are not clear.
In response to whole-blood stimulation with HK M. obuense, mDCs demonstrated a considerable increase in the expression of CD11c, CD80, CD83, CD86, CD274, CD282 (TLR2), CD284 (TLR4) and MHC class II with a concomitant decrease in the expression of CD195 (CCR5). However, pDCs showed a slight but significant increase in the expression of CD80 and MHC class II and a decrease in the expression of CD195. Based on the screened battery of surface receptors on mDCs, it was evident that whole-blood stimulation with HK M. obuense favored the up-regulation of a specific set of receptors (CD80, CD83, CD86, MHC class II), which are signature receptors correlating with the maturation status of DCs. 30 Similar induction of phenotypic maturation receptors has been reported on human Mo-DCs, phenotypically identical to mDCs, 31 by some mycobacterial species such as live or HK M. bovis, HK M. vaccae, and live, HK or γ-irradiated Mycobacterium tuberculosis,23,32–36 but not by HK Mycobacterium leprae. 34 Moreover, our data showed a significant down-regulation of CD195 (CCR5) expression on M. obuense-stimulated whole-blood mDCs and pDCs. A similar rapid down-regulation of CCR5 surface expression has been previously noted on Mo-DCs that are undergoing maturation. 37 Furthermore, HK M. obuense showed an ability to up-regulate the expression of CD274 (PDL-1) on whole-blood mDCs, a phenomenon which has been observed following stimulation of Mo-DCs with whole cell lysates of M. tuberculosis. 38
The critical role of TLRs in mediating the recognition and response to mycobacterial Ags in innate immune cells has been emphasized in several studies. TLR2 (as a heterodimer with either TLR1 or TLR6), TLR4 and TLR9 have been demonstrated to be engaged in sensing various mycobacterial cell wall components such as glyoclipids, glycoproteins, diacylated and triacylated lipoproteins, heat-shock proteins as well as unmethylated CpG motifs in mycobacterial DNA.39,40 An earlier study from our group has suggested a role for TLR2 and TLR1 in mediating HK M. obuense-induced up-regulation of CD11c and HLA-DR expression on monocytes in a whole-blood culture system. 21 The data presented here indicated that the M. obuense-induced up-regulation of CD11c, CD80 and MHC class II expression on whole-blood mDCs was mediated, at least in part, through the interaction of HK M. obuense with both TLR2 and TLR1. Using a panel of HEK293 cells expressing different human TLRs, previous studies have shown that cellular activation of HEK293 cells by HK M. vaccae or HK M. indicus pranii was mediated through interaction with TLR2 alone, with TLR2/TLR6 heterodimer and more potently with TLR2/TLR1 heterodimer.23,41 Our data also point to a putative interaction between HK M. obuense and surface TLR2/TLR1 and which could be mediated by interaction with triacylated lipoproteins, known to constitute part of the mycobacterial cell wall components.42–44 Moreover, mycobacterial triacylated lipoproteins have been reported to activate and to promote the maturation of human Mo-DCs.45,46
In this study, we also examined the effect of stimulating purified-blood DCs with HK M. obuense for 6 h on the expression of surface receptors on purified mDC and pDC subpopulations. Similar to the observed effects on whole-blood DCs, M. obuense stimulation of purified blood DCs induced a significant up-regulation in the expression of CD80 and MHC class II on both subpopulations of DCs and of CD83 and CD253 only on mDCs. Unlike whole-blood pDCs, purified-blood pDCs expressed low but detectable levels of CD83 and CD274 and up-regulated CD83 expression upon stimulation with HK M. obuense. Furthermore, in contrast to whole-blood mDCs, purified blood mDCs displayed no significant change in the expression of CD11c upon stimulation with HK M. obuense. These observed variations in M. obuense-induced surface receptor expression between whole blood and purified blood DCs might be a consequence of the DC enrichment process that eliminated other immune cells with potential contribution to the immunomodulatory effects of HK M. obuense on DCs. The differential regulation in expression of surface receptors between the two major DC subpopulations might be attributed to variations in the expression pattern of TLRs thus leading to differential recognition of HK M. obuense by mDCs and pDCs. Findings from this study, as well as from previous studies, have clearly indicated that human blood mDCs express TLR2, whereas blood pDCs lack TLR2 expression.47,48 Therefore, the absence of surface TLR2 on pDCs might explain the differential regulation in surface receptors between mDCs and pDCs in response to whole-blood stimulation with HK M. obuense. Despite the lack of TLR2 surface expression on blood pDCs, the involvement of other receptor(s) such as Dectin-2 49 and NOD250,51 in mediating the above-mentioned effects of HK M. obuense on blood pDCs should not be neglected. A putative role for human blood mDC-pDC cross-talk in mediating the observed pDCs responses to HK M. obuense, despite the absence of TLR2 expression, should be also taken into consideration. This mDC–pDC cross-talk has been reported to reverse the absence of responsiveness of purified human blood pDCs to recombinant bacillus Calmette-Guerin (rBCG) infection when cultured alone, but to allow the up-regulation of MHC class I, II, CD86 and CD123 expression when co-cultured with rBCG-infected mDCs.52–54
Cytokines and chemokines produced by DCs are critically essential in developing subsequent immune responses.55,56 In this study, we found that purified total-blood DCs stimulated with HK M. obuense mostly exhibited increased levels of secretion of several proinflammatory cytokines and chemokines, including IL-6, IL-12p40, TNF-α, CCL4, CCL5 and CXCL8. Our observation is in accordance with a previous report whereby treating Mo-DCs with HK BCG or HK M. bovis for 24 h stimulated their production of CCL5 and TNF-α. 57 Given our data showing that TLR1 and TLR2 are involved in mediating DC responses to HK M. obuense, a previous study has also demonstrated the ability of the specific TLR1/2 ligand, Pam3CSK4, to enhance the production of CXCL8, IL-6 and TNF-α by purified total blood DCs. 58 IFN-α, a cytokine reported to be mainly released in high amounts by pDCs upon activation of their TLRs, 59 was not detectable in supernatants from both unstimulated and M. obuense-stimulated purified total blood DC cultures. This observation may suggest a lack of pDC cytokine response to stimulation with HK M. obuense.
Our finding that HK M. obuense induces DC phenotypic changes consistent with a mature-like phenotype suggests that priming purified blood DCs with HK M. obuense might also result in an enhanced ability of DCs to present Ags and to activate T cells. Mo-DCs primed with HK M. vaccae have been shown to enhance autologous CD4+ T-cell responses to the recall Ag tetanus toxoid. 23 To address this hypothesis, we attempted to study the effect of priming purified blood DCs with HK M. obuense on the autologous CD4+ T-cell proliferative responses to the soluble recall Ag, C. albicans; however, our attempt was unsuccessful owing to the following technical difficulties. First, the number of purified blood DCs from 50 ml peripheral blood was low and not achieving >5 × 105 cells. This prohibited the ability of performing adequate bulk DC priming and comparing with unprimed DCs owing to cell loss upon centrifugation and washing. Second, the continued presence of HK M. obuense organisms with DCs even after bulk stimulation, centrifugation and washing. Third, the continued presence of HK M. obuense organisms in wells of cultured DCs (1 × 104 cells/well) owing to an inability to carry out vigorous washing dictated by the non-adherent nature of the purified blood DCs. Despite the above-mentioned limitations, we assessed the ability of purified blood DCs to present mycobacterial Ags and/or C. albicans Ag to autologous CD4+ T cells. We noted significant proliferation of the CD4+ T cells that were induced by M. obuense-stimulated DCs. This is in line with our previous report where we observed strong PBMC proliferative responses against HK M. obuense in healthy non-BCG-vaccinated individuals in Lebanon. 21 Our results also suggest no interference between the mycobacterial Ags and C. albicans Ag presentation mechanisms as evidenced by the additive CD4+ T cell responses induced by purified blood DCs simultaneously stimulated with HK M. obuense and C. albicans.
Collectively, our data provide a new insight into the outcome of interaction between HK M. obuense and human DCs and uncover the immunomodulatory effects of this mycobacterial preparation on DCs. Our findings might have significant implications in the design of immunotherapy protocols that could exploit the immunomodulatory properties of HK M. obuense to augment DC-mediated immune responses.
Footnotes
Acknowledgments
We would like to thank Miss Maha Said for her technical assistance.
Declaration of Conflicting Interests
The author(s) declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: SM and CA are unsalaried directors and shareholders of Immodulon Therapeutics, Ltd, UK. GMB is a member of the Scientific Advisory Board for Immodulon Therapeutics, Ltd, UK. The remaining authors declare no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by a special grant from the University of Balamand to GMB and by a grant from Immodulon Therapeutics Ltd, UK.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.