
Abstract
Many species of marine bacteria elicit a weak immune response. In this study, the aim was to assess the immunomodulatory properties of Gram-negative Pseudoalteromonas strains compared with other marine Gram-negative bacteria and to identify the molecular cause of the immunomodulation. Using murine bone-marrow derived dendritic cells (DCs), it was found that Pseudoalteromonas strains induced low cytokine production and modest up-regulation of surface markers CD40 and CD86 compared with other marine bacteria and Escherichia coli LPS. Two strains, Ps. luteoviolacea and Ps. ruthenica, were further investigated with respect to their immunomodulatory properties in DCs. Both inhibited IL-12 and increased IL-10 production induced by E. coli LPS. LPS isolated from the two Pseudoalteromonas strains had characteristic lipid A bands in SDS-PAGE. Stimulation of HEK293 TLR4/MD2 cells with the isolated LPS confirmed the involvement of LPS and TLR4 and established Pseudoalteromonas LPS as TLR4 antagonists. The isolated LPS was active in the endotoxin limulus amoebocyte lysate assay and capable of inducing increased endocytosis in DCs. This study highlights that antagonistic LPS from Pseudoalteromonas strains has potential as a new candidate of therapeutic agent capable of modulating immune responses.
Introduction
Marine bacteria and other marine microorganisms offer a rich source of useful biomolecules, such as membrane lipids, proteins, complex polysaccharides, pigments, nucleic acids and a range of bioactive secondary metabolites.
1
These bioactive molecules hold potential for pharmaceutical and biotechnical applications.
1
The genus Pseudoalteromonas comprises Gram-negative marine bacteria belonging to the γ-proteobacteria and is present globally in marine waters, constituting 0.5–6% of the total bacterial plankton.
2
Marine bacteria produce extracellular polysaccharides (EPS) and various low molecular mass (LMM) compounds as a strategy for growth, adhesion to solid surfaces and survival in adverse conditions, for example by biofilm formation.3,4 In most Gram-negative bacteria, LPS is a very potent virulence factor that activates innate immune cells primarily through TLR4.
5
Upon CD14 binding of LPS, LPS is transferred to MD2, which then forms complexes with TLR4 leading to TLR4 dimerization and initiation of the MyD88- or TRIF-dependent pathway.6,7 Lipid A is the active component of the LPS that stimulates the TLR4 receptor complex. Lipid A structures exhibit considerable variability between bacterial species, but have little intra-genus variability.1,8 The structural differences, in particular the degree of acylation, can influence their pro-inflammatory capacity. In general, highly acylated (hexaacylated) lipid A, for example from Escherichia coli constitute potent inflammatory stimulators, whereas less acylated lipid A is less stimulatory or even displays antagonistic properties.8,9 As an example, the tetra-acylated lipid A from Yersinia pestis is a weak immunostimulator in human macrophage U937 cells.
10
Likewise, LPS from Gram-negative marine bacteria often elicits a weak immune response only.
1
Lipid A from Gram-negative marine bacteria may, instead, act as putative antagonists of LPS from E. coli and thereby represent potential candidates in the development of drugs.
11
So far, LPSs from five different Pseudoalteromonas strains have been characterized and they have all been found to be pentaacylated.12–16 LPS is composed of a hydrophilic heteropolysaccharide (formed by core oligosaccharide and an O-chain polysaccharide moiety) linked covalently to the lipophilic lipid A domain. The polysaccharide (PS) moiety is linked to the lipid A part by 3-deoxy-
Dendritic cells (DCs) represent a group of innate immune cells centrally involved in the initiation of both innate and adaptive immunity for immediate and long-term protection respectively. Pattern recognizing receptors (PRRs) expressed on DCs and other immune cells contribute to the specific recognition of pathogens and microbial ligands. 5 Recognition of an antigen by DCs involves receptor–ligand interaction, mediating intracellular signalling cascades leading to DC maturation, including production and release of cytokines, which initiates recruitment of pro-inflammatory cells and determine the resulting T cell repertoire.5,21 Hitherto, the immunostimulatory effects of Pseudoalteromonas and other Gram-negative marine bacteria have not been studied in DCs.
The immunostimulatory potential of LPS is dependent on the structure of the lipid A part. In the monocytic THP-1 cell line tetra- and pentacylated LPS were weak immune stimulators with antagonistic properties. 22 Hexa-acylated lipid A from marine Vibrio cholerae, a known human pathogen, was able to induce TLR4 activity in HEK293 cells transfected with TLR4/MD-2 and co-receptor CD14. This study also found that loss of hexa-acylation reduced TLR4 activation. 23 Moreover, it has been demonstrated that LPS may act as an antagonist through competition for the TLR4/MD-2 receptor complex in human monocyte-derived DCs. 24 Recently, a study by Carillo et al. 16 characterized the structure of lipid A from Pseudoalteromonas haloplanktis TAB 23. They found that this penta-acylated lipid A was able to inhibit E. coli LPS-induced production of TNF-α and IL-6 in human monocytic THP-1 cells. 16 As all LPSs so far isolated from Pseudoalteromonas are penta-acylated,12–14,16 we hypothesized that most, if not all, Pseudoalteromonas strains are low immunostimulators with TLR4 antagonist potential.
In this study, we therefore aimed to assess the immunostimulatory properties of several Pseudoalteromonas strains compared with other marine Gram-negative bacteria and to identify molecules involved in the observed weak stimulation.
Materials and methods
Generation and stimulation of bone marrow-derived DCs
Bone marrow cells were isolated and cultured as described previously.
25
Briefly, femurs and tibias were removed from C57BL/6 mice and stripped of muscles and tendons, and the marrow was flushed. The resulting cell suspension was washed and cells were re-suspended in RPMI 1640 (Lonza, Walkersville, MD, USA) supplemented with 4 mM
In experiments involving stimulation of DCs with spent medium (SM) from bacterial cultures, a fixed volume of SM was added. We estimated that when stimulating with SM (dry mass around 38–39 mg/ml) diluted 1:36, this would equal approximately 1 mg/ml.
Bacterial cultures
Bacterial strains S2604, S2753, S2052, 234.10, 1926, 3655, S3258, S4054, S1189 and S4498 were collected during the Galathea 3 marine research expedition (Figure 1C) and grown in 50 ml marine broth (MB) 2216 solution (37.4 g/l; Difco, Detroit, MI, USA) and on marine agar (MA) 2216 (55.1 g/l; Difco). Alternatively, strains were also grown in autoclaved sea salt solution (SSS) (Sigma S9883; 30 g/1) to which 4 g/l mannose (Fluka), and 0.3 g/l casamino acids (223050; Difco) were added from sterile, filtered stock solutions at 20% v/w. If necessary, the substrate was supplemented with a 1.0% Bacto agar (BDAA503-957; Becton Dickinson, Franklin lakes, NJ, USA). Cultures were grown with agitation for 2 d at 25°C and centrifuged at 10,000 g. Bacterial pellets were washed twice with ice cold sterile PBS (Lonza) and re-suspended in 10 ml PBS. Bacteria were irradiated in a laboratory reactor (CL-1000 UV crosslinker; UCP, Cambridge, UK) at 25,000 μJoule/cm2 for 2 × 6 s at a monochromatic wavelength of 254 nm. Viability was checked by plating onto MA plates. The suspensions were aliquoted into sterile Eppendorf tubes and stored at −80°C until use.
Induction of cytokines and cell surface marker up-regulation in DCs by Gram-negative marine bacteria (1–100 µg/ml) including five different Pseudoalteromonas strains. (A) Supernatants from bacteria-stimulated DCs were harvested after 20 h and concentrations of IL-10, IL-12, TNF-α and IL-6 were measured by ELISA. Data are presented as means of triplicate cultures ± SD. Results are representative of one of three independent experiments. (B) Expression of surface markers CD40 and CD86 on DCs upon exposure to ultraviolet-killed 1 µg/ml Ps. ruthenica (S3258), Ps. luteoviolacea (S4054), R. mobilis (1926) and LPS from E. coli. DCs were stimulated with 1 µg/ml (black line). Results for unstimulated DCs are also depicted (filled histogram). The light grey histogram indicates the unspecific binding of isotype Abs. The numbers in the upper right corner of each histogram represent the geometric MFI obtained with 1 µg/ml of stimulus or isotype staining. Data are representative of one of at least two independent experiments. (C) Bacteria used in the experiment.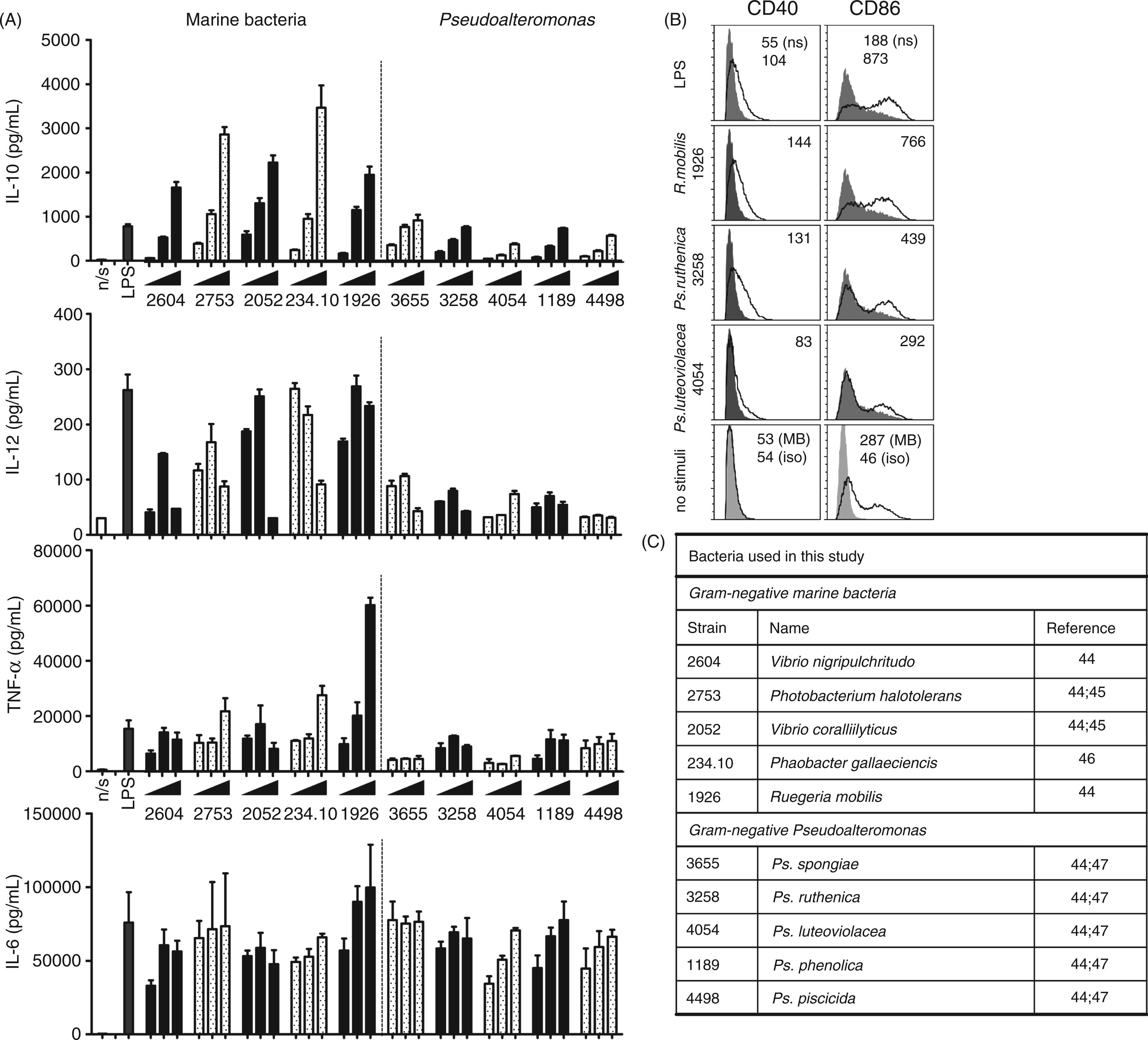
The supernatant or SM from 2-day-old bacterial cultures was collected, aliqouted and stored at −80°C. Selected SM samples were processed for size separation using PD-10 columns (GE Healtcare, Uppsala, Sweden) giving rise to a high molecular mass (HMM) fraction (>5000 u). Briefly, PD-10 columns were washed with phosphate buffer (pH 7.5) and 2.5 ml SM from bacterial cultures were added to the columns. The macromolecular fraction was collected by adding 3.5 ml of elution buffer and collecting the flow-through. The suspension of macromolecules from SM was diluted by a factor of 1.4.
Flow cytometry analysis
Stimulated mature DCs were harvested and re-suspended in ice-cold PBS containing 1% (v/v) FBS and 0.15% (w/v) sodium azide (PBSAz). Cells were incubated with anti-mouse FcγRII/III for 10 min (BD Biosciences, San Jose, CA, USA) to block non-specific binding of Ab reagents. The cells were stained with the following Abs for 45 min: Allophycocyanin (APC)-conjugated anti-mouse CD86, phycoerythrin (PE)-conjugated anti-mouse CD40, (both from eBioscience, San Diego, CA) FITC-conjugated anti-mouse CD11c (BD Pharmingen, San Diego, CA, USA). Non-specific binding was evaluated by including matched isotype controls for all the Abs used. DCs were analysed using a BD FACSCanto II flow cytometer (BD Biosciences) based on counting 10,000 cells. The level of expression was expressed as the geometric mean fluorescence intensity (MFI) and percentage of events.
ELISA
Quantitative assays for IL-12(p70), IL-10, IL-6, TNF-α and IL-8 in the culture supernatants were performed using ELISA kits (R&D Systems, Minneapolis, MN, USA) according to the manufacturer's instructions. Results were analysed as the mean concentrations ± SD in triplicate culture supernatants.
Macropinocytosis assay
Cells (4 × 105) were seeded into 96-well plates and pre-incubated at 37°C with medium or stimuli for 30 min in final concentrations as indicated. Cells were incubated for 10 min with 200 µg/ml FITC-dextran (10 ku, Sigma) at 37°C. Macropinocytosis was stopped by 2 washes in ice-cold PBS containing 0.15% (w/v) sodium azide and 1% (v/v) FCS (PBS-Az), and immediately hereafter cells were fixed in PBS-Az containing 1% (v/v) formalin and analysed on a BD FACSCanto II (BD Biosciences) based on counting 10,000 cells.
Isolation and characterization of LPSs
LPSs from Ps. luteoviolacea S4054 and Ps. ruthenica S3258 grown in MB media, as described earlier, was obtained with a hot phenol–water mixture according to the protocol by Hassan et al. 26 Briefly, cells were collected by centrifugation, washed with water and dried with cold acetone for 3 × 1 h. Dried cells were extracted three times with a mixture of aq. 90% phenol/chloroform at 65°C. After cooling, LPS was recovered in the water phase by centrifugation. After 48 h of dialysis (16 mm Servapor; Serva, Electrophoresis, Heidelberg, Germany) against sterile Milli Q water the obtained LPS was lyophilized and resolved in sterile Milli Q water to obtain the concentration of 5 mg/ml.
Mild hydrolysis of purified LPS fractions was performed according to the protocol of Chhibber et al. 27 Briefly, 1000 μl of purified LPS from Ps. luteoviolacea S4054 and Ps. ruthenica S3258, respectively (5 mg/ml in water), were mixed with 1000 μl 2% acetic acid, and incubated for 2 h in a boiling water bath in closed tubes. After storage at 4°C overnight (20 h), the tubes were centrifuged for 15 min at 1073 g (MiniSpin Eppendorf). Each supernatant was extracted with an equal volume of a 2:1 mixture of chloroform and methanol. This was done three times—collecting the upper (water) phase each time. The final phase was then freeze-dried yielding a white powder that was dissolved to approximately 5 mg/ml in water for further analysis.
SDS-PAGE of LPS and endotoxin assay
LPS samples from Ps. ruthenica S3258, Ps. luteoviolacea S4054 and E. coli O26:B6 (product number: L2654, Sigma) were mixed (1:4) with NuPage LDS sample buffer (4×) (Invitrogen, Carlsbad, CA, UK). The mixtures were heated in a 100°C hot plate for 5 min. Each sample was applied to 1.0 mm sample wells and subjected to electrophoresis in 10% Bis-Tris gels (Invitrogen) in NuPage MES-SDS running buffer (20× ) at 200 volt for 45 min. After electrophoresis, the gels were stained with coomassie or silver nitrate using a slightly modified protocol from Tsai et al. 28 The gels were fixed for 1 h in 30% ethanol–12% acetic acid at room temperature (20–22°C, RT). In order to oxidize samples, the gel was treated with periodic acid, making the silver stain capable of staining carbohydrate and protein content. 28 When the samples were assayed for protein content, pretreatment of the gel with periodic acid was omitted.
Selected samples were tested for endotoxin content by limulus amoebocyte lysate (LAL)-assay according to the manufacturer’s protocol (Lonza).
TLR4 activation
Transfected TLR4 HEK293 cells were generously donated by Søren Paludan (Århus University).7,29 The transfected cells were grown in DMEM (Lonza, Walkersville, MD, USA) supplemented with 10% FCS and antibiotics (500 µg/ml G418) (Roche, Basel, Switzerland). The transfected cells were seeded in 48-well tissue plates at a density of 4 × 104 cells per well and left overnight.
Next day, cells were stimulated with LPS (1 µg/ml) from E. coli O26:B6, 1–100 µg/ml of purified LPS from Ps. ruthenica S3258 or Ps. luteoviolacea S4054 for 24 h at 37°C in a 5% CO2 humidified atmosphere. For co-stimulatory experiments, purified Pseudoalteromonas LPS were added 30 min prior to stimulation with LPS from E. coli. Supernatants were harvested and stored at −80°C until IL-8 levels were measured by ELISA.
Statistical analysis
Data were analyzed for statistical significance (GraphPad Prism, version 5.0, GraphPad Software, San Diego, CA, USA) using one-way ANOVA and the Dunnett test [comparing individual treatments with that of a control treatment (LPS or medium)]. P values were <0.001, <0.01 or <0.05. Results are shown as the mean ± standard deviation of triplicates.
Results
Pseudoalteromonas induces weak IL-12 production in bone marrow-derived DCs
It has been reported previously that many marine bacteria show low immunostimulatory capacity. Thus, to establish the immunostimulatory capacity of the Pseudoalteromonas genus, five strains of Pseudoalteromonas (Ps. spongiae, Ps. ruthenica, Ps. luteoviolacea, Ps. phenolica, and Ps. piscicida) were added to bone marrow-derived DCs. To establish their relative capacity to mature DCs, we included five other marine bacteria representing four genera and LPS from E. coli in the analysis (Figure 1A). The production of IL-10, IL-12, IL-6 and TNF-α in DCs was measured after 20 h of exposure to the bacteria at different concentrations. Compared with the other marine bacteria, all Pseudoalteromonas strains only induced weak IL-12, IL-10 and TNF-α responses, while the IL-6 response was comparable to that induced by the other bacteria. When compared with LPS from E. coli the IL-6, IL-10 and TNF-α response to Pseudoalteromonas strains was comparable to that induced by LPS (1 µg/ml). The IL-12-induced response did not reach more than 100 pg/ml compared with the 240 pg/ml induced by E. coli LPS. Most of the other bacteria were capable of inducing an IL-12 level comparable to that induced by E. coli LPS. For most bacteria, including the Pseudoalteromonas strains, the level of IL-12 peaked at 1 or 10 µg/ml and then decreased again at higher bacterial concentrations. LPS from E. coli was included in 1 µg/ml. This was based on a titration study showing a slight increase in the induced IL-12 production in the range 1–30 µg/ml (data not shown).
In order to further analyse the maturation pattern of DCs exposed to different Gram-negative marine bacteria, the expression of CD86 and CD40 was measured by flow cytometry upon stimulation with two different Pseudoalteromonas strains (Ps. ruthenica S3258 and Ps. luteoviolacea S4054) representing low cytokine stimulatory bacteria and R. mobilis representing high cytokine stimulatory bacteria respectively (Figure 1B). While CD40 was only up-regulated in the presence of bacteria, even addition of MB (the bacterial growth medium) gave rise to up-regulation of CD86. LPS and R. mobilis were most potent stimulators of CD86 up-regulation, but Ps. ruthenica also stimulated a significant up-regulation, while, upon stimulation with Ps. luteoviolacea, the level corresponded to that induced by addition of MB. Ps. ruthenica and R. mobilis were the most potent inducers of CD40 and induced a level comparable to that induced by LPS from E. coli, while Ps. luteoviolacea induced a lower CD40 level, though it was still higher than in unstimulated DCs. The expression of CD86 was up-regulated further upon stimulation with a higher dosage of Ps. luteoviolacea (10 µg/ml) (data not shown) to levels similar to that of 1 µg/ml Ps. ruthenica.
These data show that all the five Pseudoalteromonas strains are weak inducers of IL-12, IL-10 and TNF-α, but induce IL-6 to similar levels of E. coli LPS-stimulated DCs and up-regulated the co-stimulatory markers CD40 and CD86.
Pseudoalteromonas inhibits IL-12 while increasing IL-10 production induced by LPS from E. coli
The poor IL-12-inducing capacity of the Pseudoalteromonas strains might be a result of Pseudoalteromonas LPS acting as a TLR4 antagonist, binding to TLR4 but only weakly stimulating the DCs. Alternatively, the presence of other microbial ligands might act contra-active and thus, indirectly, inhibit TLR4 signalling in DCs. To address a possible inhibitory effect, we added increasing concentrations of three strains of Pseudoalteromonas together with LPS from E. coli (1 µg/ml) to DCs. Ps. luteoviolacea, Ps. ruthenica and Ps. spongiae inhibited the LPS induced IL-12 production, while slightly enhancing the LPS induced IL-10 production (Figure 2A and B). In contrast, none of the three strains were able to significantly alter the LPS induced IL-6 or TNF-α cytokine production (data not shown). To rule out that the effect on IL-12 production was not caused by LPS desensitization, we stimulated DCs with two consecutive exposures of 1 µg/ml LPS E. coli (Figure 2C). In contrast to DCs exposed to Pseudoalteromonas prior to stimulation with LPS from E. coli, these DCs showed no reduction in IL-12 production. Furthermore, application of Pseudoalteromonas (10 µg/ml) after E. coli LPS stimulation did not result in a reduction in the IL-12 response.
Pseudoalteromonas strains inhibit E. coli LPS-stimulated induction of IL-12. (A, B) DCs were stimulated with whole bacteria alone or in co-incubation with 1 µg/ml E. coli LPS, as indicated. After 20 h supernatants were harvested and IL-10 and IL-12 were measured with ELISA. (C) DCs were stimulated with 1 µg/ml E. coli LPS for 30 min followed by a second exposure of 1 µg/ml E. coli LPS or 10 µg/ml Ps. ruthenica 3258. DCs were also stimulated with 10 µg/ml Ps. ruthenica 3258 for 30 min followed by exposure to 1 µg/ml E. coli LPS as in Figure 2A and B. After 20 h supernatants were harvested and IL-12 was measured with ELISA. Data represent the mean of measurements from triplicate cultures ± standard deviation, ***P < 0.001, **P < 0.01 indicate cytokine levels induced by the Pseudoalteromonas strains that are significantly different from unstimulated cells. ###P < 0.001, ##P < 0.01, #P < 0.05 indicate cytokine levels significantly different from the level induced by LPS alone. The data represent one of at least three independent experiments.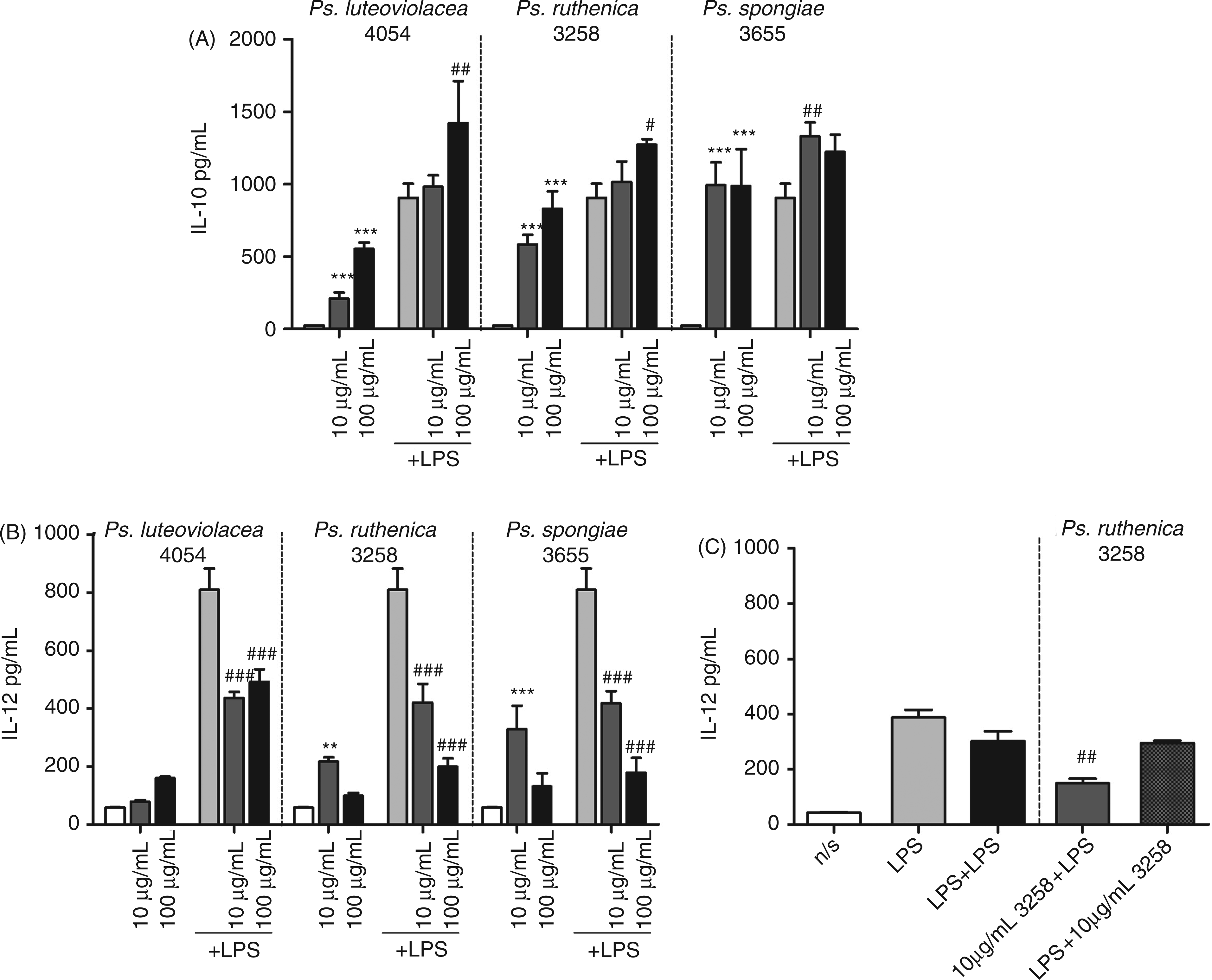
SM and secreted macromolecules from Ps. luteoviolacea hold low immunostimulatory properties and inhibit LPS-induced IL-12 production
LPS and other constituents on the bacterial surface may be present in the SM, as bacteria shed outer membrane fragments containing LPS during growth,
17
and some compounds, for example PS and LMM compounds, are actively secreted.
30
As an example of a secreted compound, strains of Pseudoalteromonas produce large quantities of EPS, presumably dependent on the growth conditions.4,31 To investigate the immunomodulatory activities of secreted molecules from Ps. luteoviolacea S4054, we examined whether the SM from the bacterial cultures was able to exhibit the IL-12 production inhibitory effects observed by the whole bacteria. As the growth medium contains yeast extract, which may hold similar immunomodulatory effects, we included fresh medium diluted in the SM as a control. As depicted in Figure 3A, fresh medium per se exhibited a dose-dependent down-regulation of the LPS-induced IL-12, but did not induce any cytokines when added to the cells alone. In contrast, the SM induced IL-10, IL-6 and TNF-α production by itself, but not IL-12 production. However, when added together with LPS, SM inhibited IL-12 (85% and 92% for the 108 - and 36-fold dilution) markedly more than fresh medium (a 38% and 70% inhibition for the 108 - and 36-fold dilution respectively). SM from Ps. luteoviolacea up-regulated IL-10 production, presumably owing to an additive effect of LPS and the immunomodulating compound(s) in the SM. An inhibitory effect on LPS-induced IL-6 production from both fresh medium and SM was also seen. Here, the inhibition was slightly stronger for the fresh medium than for SM (down to approximately 50 ng/ml and 60 ng/ml respectively). In contrast to the slight TNF-α down-regulation of the fresh medium, the SM from cultured Ps. luteoviolacea did not change the LPS-induced production of TNF-α.
SM from Pseudoalteromonas growth and macromolecules from the SM inhibit LPS induced IL-12 production. (A) Induction of cytokine production with stimulation of SM from Ps. luteoviolacea S4054 or blank marine broth ± 1 µg/ml LPS. Five hundred microlitres of DCs were added to 50 μl of SM (diluted 1:3) for 30 min, then 50μl LPS was added and incubated for 20 h. SM was added as dilution fractions instead of absolute concentrations giving final dilution concentrations of 1:36 and 1:108. (B) Molecules above 5 ku were isolated from SM by gel filtration from bacterial culture Ps. luteoviolacea S4054 grown in marine broth. During this process the HMM fraction was diluted approximately three times. DCs were stimulated with the HMM fraction in either 1/1500 or 1/750 dilutions (final dilutions) for 30 min followed by addition of 1 µg/ml LPS or medium and incubated for 20 h. Supernatants were harvested and cytokine concentrations of IL-10, IL-12, IL-6 and TNF-α were measured by ELISA. Error bars depict mean ± SD of triplicate determinations. The data represent one of at least two independent experiments. ***P < 0.001, **P < 0.01, *P < 0.05 indicate cytokine levels significantly different from the level induced by LPS alone.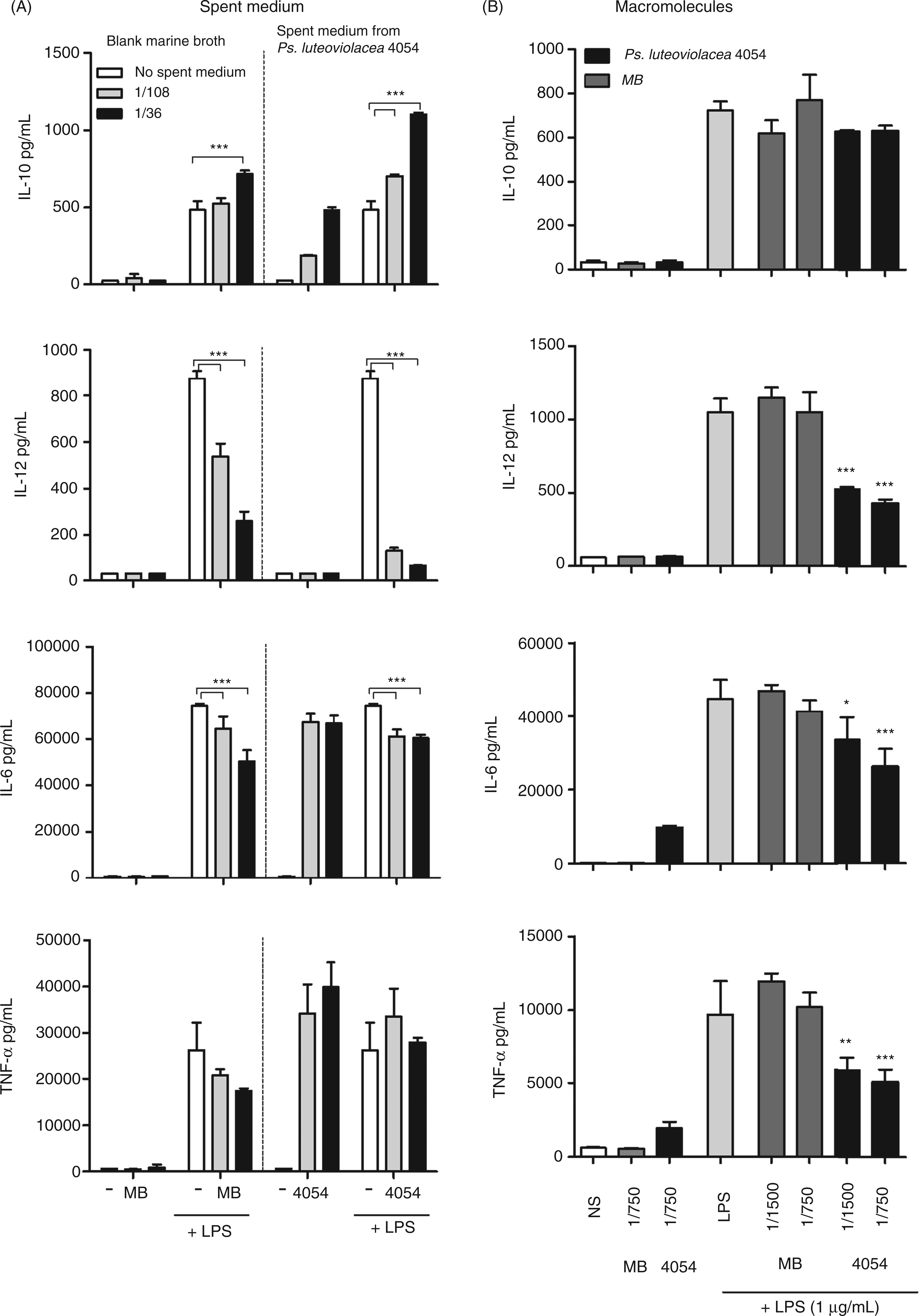
To exclude that the inhibiting effect was caused by LMM molecules, we separated compounds of a molecular mass of less than 5 ku from the rest of the constituents in the SM from the bacterium Ps. luteoviolacea S4054 and marine broth by size-exclusion chromatography (Figure 3B). The HMM fractions from fresh marine broth and SM from Ps. luteoviolacea 4054 were diluted to reach final concentrations of 1/750 and 1/1500 during DC stimulation. The macromolecules from Ps. luteoviolacea exhibited dose-dependent inhibitory effects on LPS-induced IL-12, TNF-α and IL-6. The HMM fraction from Ps. luteoviolacea SM per se stimulated the induction of IL-6 and TNF-α, but not IL-10 and IL-12 production. In contrast to the complete SM, we did not observe an increase in the IL-10 production after stimulation with LPS and the HMM fractions. The HMM fraction from marine broth did not have any significant effect on LPS-induced cytokine production nor was it able per se to induce cytokine production in DCs. Taken together these results show not only that the studied strains of Pseudoalteromonas are poor stimulators of DC cytokine production and inhibitors of IL-12 production, but also that shed or secreted HMM components in the SM hold such properties.
LPS from Pseudoalteromonas is a TLR4 antagonist and induces macropinocytosis in DCs
In order to establish whether the inhibitory effects were caused by LPS, we isolated LPS from Ps. ruthenica S3258 and Ps. luteoviolacea S4054 grown in MB and compared them with LPS from E. coli by SDS-PAGE (Figure 4A). LPS from E. coli exhibited the traditional ladder-like banding pattern, including a distinct band at 30 ku (Figure 4A), while LPS from Ps. ruthenica S3258 (lanes B, C and D) exhibited two distinct bands around 3–15 ku, indicating a composition of the LPS with LMM carbohydrates characteristic for smooth Gram-negative bacteria.
32
In contrast, LPS from Ps. luteoviolacea S4054 (lane E, F and G) gave rise to a band at 30 ku similar to that seen for E. coli LPS, but much stronger. In addition, all three LPS samples gave rise to the characteristic lipid A band at the bottom of the gel. The band in Pseudoalteromonas LPS were lowest and thereby indicative of a lower molecular mass than lipid A from E. coli.
LPS from Pseudoalteromonas shows strong presence of a lipid A band. (A) Purified LPS fractions visualized on SDS-PAGE with silver staining. Lane (A) Mark12TM unstained standard, (B) Ps. ruthenica S3258 grown in MB 56.3 µg, (C) Ps. ruthenica S3258 grown in MB 37.5 µg, (D) Ps. ruthenica S3258 grown in MB 18.8 µg, (E) Ps. luteoviolacea S4054 grown in MB 56.3 µg, (F) Ps. luteoviolacea S4054 grown in MB 37.5 µg, (G) Ps. luteoviolacea S4054 grown in MB 18.8 µg, (H) LPS from E. coli 11.3µg, (I) LPS from E. coli 7.5 µg and (J) LPS from E. coli 3.8 µg. (B) DCs were stimulated with purified Pseudoalteromonas LPS (0.1–100 µg/ml) for 30 min followed by co-stimulation with 1 µg/ml LPS from E. coli or medium for 20 h. Supernatants were harvested and cytokine concentrations of IL-10, IL-12, IL-6 and TNF-α were measured by ELISA. Error bars depict mean ± SD of triplicate determinations. The data represent one of at least two independent experiments. ***P < 0.001, **P < 0.01, *P < 0.05 indicate cytokine levels significantly different from the level induced by LPS alone. (C) Macropinocytosis in DCs upon stimulation with 1 µg/ml of purifed LPS from Ps. luteoviolacea S4054, Ps. ruthenica S3258 or E. coli for 30 min followed by addition of FITC-dextran particles for 10 min. FITC-positive cells were measured by flow cytometry from triplicate determinations. ***P < 0.001 (unstimulated vs stimulated). The results are representative for one of three independent experiments.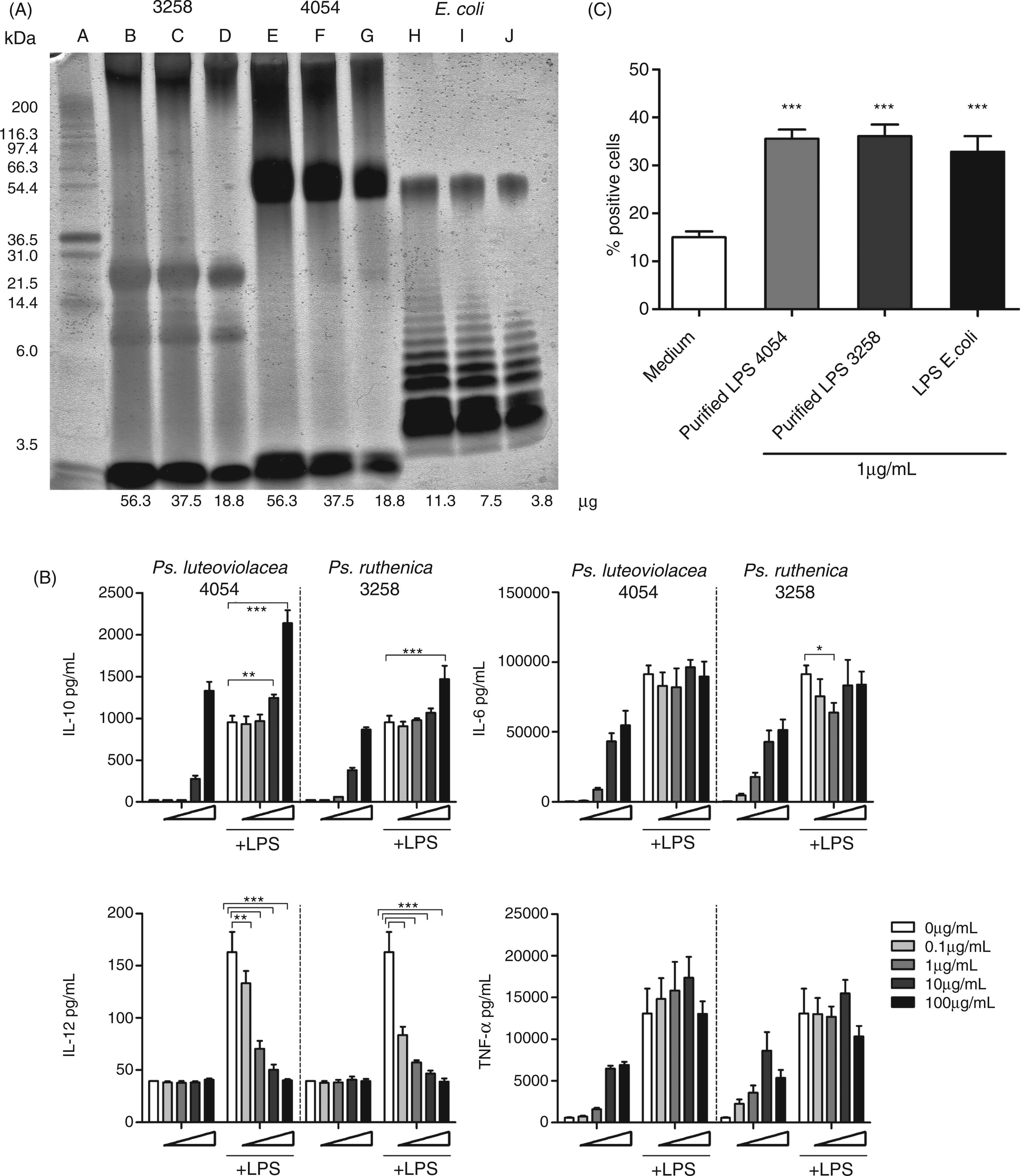
To assess the stimulatory and inhibitory activity of the isolated LPS, DCs were stimulated with 0.1–100 µg/ml of purified LPS from Ps. ruthenica S3258 and Ps. luteoviolacea S4054 for 30 min followed by addition of LPS from E. coli (1 µg/ml) or medium. LPS isolated from both Pseudoalteromonas strains were able to induce IL-10 production in DCs, but about a 100-fold higher concentration was required to induce the same up-regulation of IL-10 as 1 µg/ml of LPS from E. coli (Figure 4B). The isolated LPS also induced a dose-dependent production of TNF-α and IL-6, but even with addition of a 100-fold higher concentration, the production did not reach the concentrations produced upon addition of 1 µg/ml LPS from E. coli. As for the intact bacteria, the isolated LPS did not give rise to any IL-12 production. Incubation of the LPS from Pseudoalteromonas, together with the E. coli LPS, revealed that despite no IL-12 inducing capacity per se, the LPS isolated from Pseudoalteromonas were potent IL-12 inhibitors, decreasing the IL-12 production significantly even when added at a 10-fold lower concentration than the E. coli LPS (Figure 4B). The IL-10 production upon co-stimulation increased approximately corresponding to the production induced by the Pseudoalteromonas LPS per se, while addition of Pseudoalteromonas LPS did not affect the E. coli LPS induced production of TNF-α and IL-6.
LPS from E. coli has been shown to induce an increased endocytosis of dextran particles in DC. 33 To investigate the endocytosis-inducing capacity of the LPS purified from Pseudoalteromonas we stimulated DCs with 1 µg/ml of LPS from Ps. luteoviolacea, Ps. ruthenica or E. coli for 30 min prior to addition of FITC-conjugated dextran particles (Figure 4C). All three LPS preparations were equally potent in increasing the endocytosis of the dextran particles. Taken together, these results show that despite their action as antagonist in regard to LPS-induced IL-12 production, LPS preparations from Pseudoalteromonas are fully capable of inducing enhanced endocytosis of dextran.
LPS is generally regarded to stimulate cells through TLR4. In cooperation with MD2, TLR4 binds LPS, dimerizes and transduces the activation signal to the internal of the cells, which initiates the activation pathway.
7
To examine if the inhibitory effect of Pseudoalteromonas LPS is mediated through TLR4, we stimulated HEK293 cells stably expressing TLR4, together with MD2/CD14 and using IL-8 as reporter system, with increasing amounts of LPS purified from Ps. luteoviolacae or Ps. ruthenica. Neither of the two purified LPS preparations nor the TLR2 ligand Pam3Csk4 were capable of inducing IL-8, while E. coli LPS induced high IL-8 production (Figure 5A). Co-incubation of HEK293 TLR4 cells with Pseudoalteromonas LPS and E. coli LPS gave a dose-dependent inhibition at Pseudoalteromonas LPS doses 10- to 100-fold higher than the E. coli LPS added (Figure 5B). Pam3Csk4 did not change the E. coli LPS induced production of IL-8. This indicates that LPS from Pseudoalteromonas binds to TLR4 but is unable to stimulate through TLR4 and, hence, acts as a true TLR4 antagonist.
Activation of TLR4 in TLR4-stimulated HEK293 cells with purified LPS from marine bacteria. (A) HEK293-TLR4/MD2 cells were stimulated with purified 1–100 µg/ml LPS from Ps. luteoviolacea, Ps. ruthenica, E. coli (1 µg/ml) or TLR2 ligand Pam3Csk4 (1 µg/ml) for 30 min. (B) Hereafter, pre-stimulated cell were co-incubated with 1 µg/ml LPS from E. coli or medium as indicated. Supernatant was harvested after 24 h of incubation and IL-8 cytokine production was measured by ELISA. Data are presented as means of triplicate cultures ± SD. ***P < 0.001 indicates cytokine levels significantly different from the level induced by LPS alone. Results are representative for one of three independent experiments.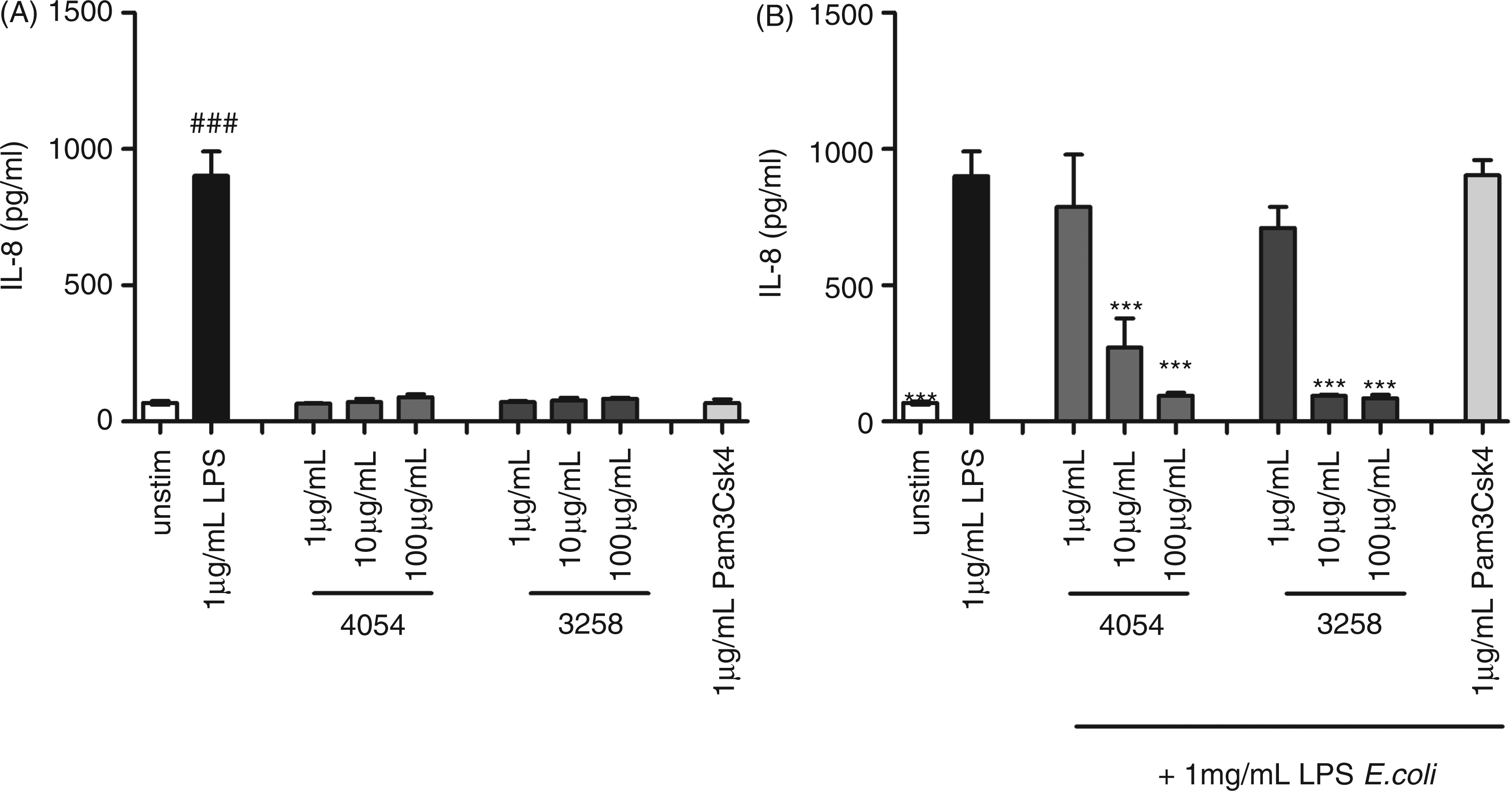
SM from Ps. luteoviolacea is able to inhibit an E. coli LPS-induced TLR4 activation owing to the presence of lipid A
To confirm that the IL-12 inhibitory effect of the SM was due to direct inhibition of the TLR4 receptor, HEK293 cells were incubated with SM from Ps. luteoviolacea and Ps. ruthenica in the presence or absence of LPS from E. coli (Figure 6A). SM from neither Ps. luteoviolacea nor Ps. ruthenica S3258 induced TLR4 activation in HEK293 TLR4 cells. Upon co-incubation with LPS from E. coli, SM from Ps. luteoviolacea showed a strong inhibition of the TLR4 activation (93%), whereas SM from Ps. ruthenica S3258 exhibited a weak, though still significant (reaching around 60%), inhibition at the highest concentration (1/12). Fresh medium did not induce TLR4 activation, but, when added together with E. coli LPS, we found significant inhibition (40%) of TLR4 activity, which was lower than the strongest inhibition seen for Ps. ruthenica SM (Figure 6A).
Activation of TLR4 in HEK293 cells TLR4 stimulated with SM from bacterial cultures. (A) HEK293-TLR4/MD2 cells were stimulated with SM from marine bacterial cultures grown in MB at dilutions; 1/108-1/12 for 30 min, followed by stimulation with 1 µg/ml LPS from E. coli or medium as indicated. Supernatant was harvested after 24 h of incubation and IL-8 production was measured by ELISA. (B) HEK293- TLR4/MD-2 cells were stimulated with PS fractions (5 and 50 µg/ml) from Ps. luteoviolacea and Ps. ruthenica, obtained by treating marine LPS with mild acidic hydrolysis. Cells were stimulated with marine LPS 50 µg/ml followed by stimulation with 1 µg/ml LPS from E. coli or medium. Supernatant was harvested after 24 h of incubation and IL-8 production was measured by ELISA. Data are presented as means of triplicate cultures ± SD. ***P < 0.001 indicates cytokine levels significantly different from the level induced by LPS alone. Results are representative for one of two independent experiments.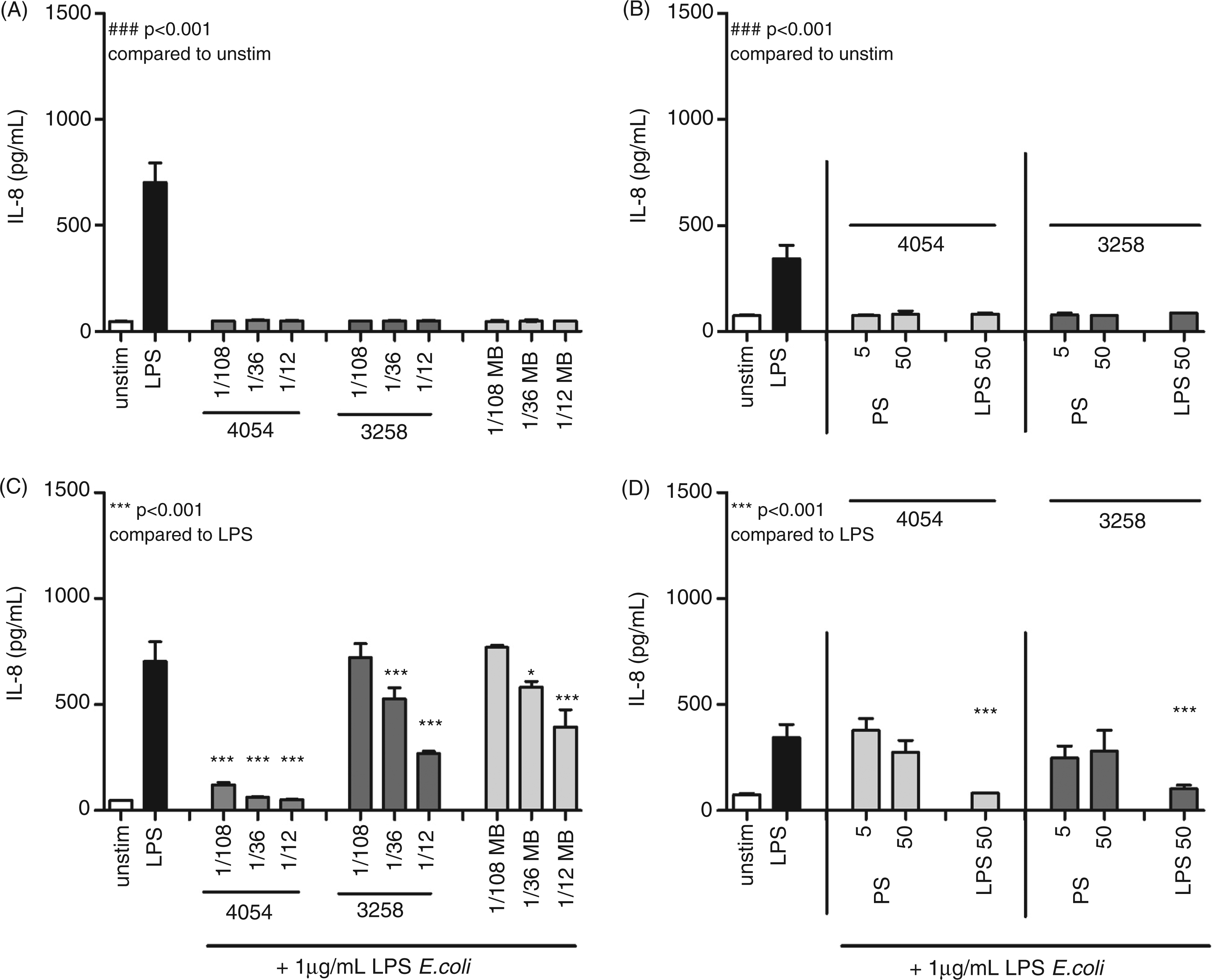
Endotoxin content.
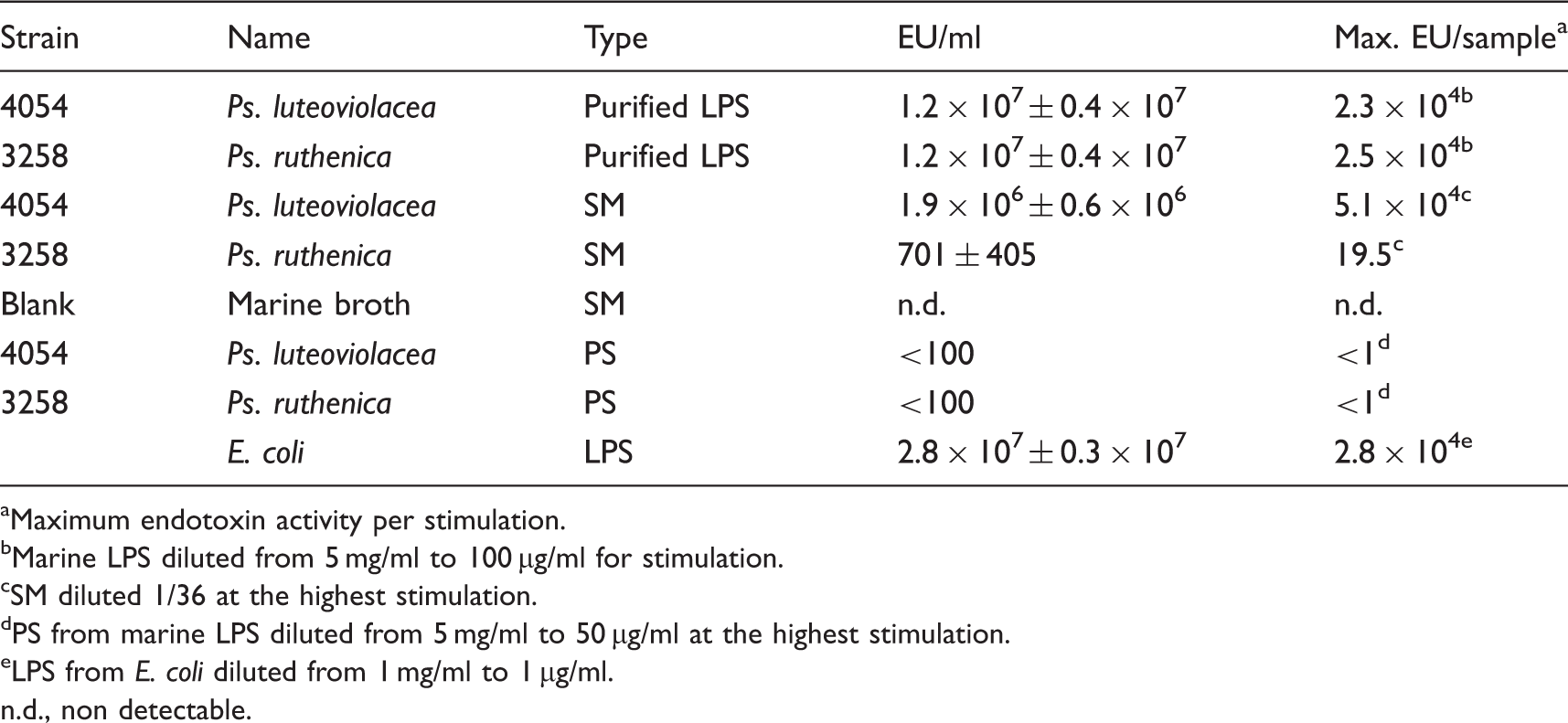
Maximum endotoxin activity per stimulation.
Marine LPS diluted from 5 mg/ml to 100 µg/ml for stimulation.
SM diluted 1/36 at the highest stimulation.
PS from marine LPS diluted from 5 mg/ml to 50 µg/ml at the highest stimulation.
LPS from E. coli diluted from 1 mg/ml to 1 µg/ml.
n.d., non detectable.
Finally, we separated the lipid A part from LPS by a mild acidification and obtained PS with very low endotoxic activity (<100 EU/ml) for both strains, indicating a pure PS fraction. When tested in the TLR4 HEK293 cells (Figure 6B) they did not induce TLR4 activity nor did they show any significant ability to decrease E. coli LPS-induced activation. The various purified fractions in Table 1 were run on a SDS-PAGE followed by silver staining (data not shown). As found previously, we observed that LPS from both Ps. luteoviolacea and Ps. ruthenica had a characteristic lipid A band at the bottom of the gel. This characteristic lipid A band was also found for SM from Ps. luteoviolcea (data not shown) confirming the data from the LAL assay. In accordance with the data from the LAL-assay, SM from Ps. ruthenica, MB and the two PS fractions did not show any lipid A like-bands (data not shown).
Taken together, these results demonstrate the presence of lipid A in the SM of Ps. luteoviolacea, as the cause of the antagonistic effect on TLR4 activation and the IL-12 inhibitory effect in E. coli LPS-stimulated DCs.
Discussion
Marine microorganisms represent a novel source of bioactive molecules that may be exploited for pharmaceutical purposes.1,34 With this in mind, in the present study we studied the immunomodulatory properties of the marine bacteria genus Pseudoalteromonas in murine bone marrow-derived DCs. In comparison with a selection of other bacteria of marine origin, such as Vibrio and Ruegeria, and E. coli-derived LPS, the Pseudoalteromonas strains exhibited poor immunostimulatory activity. This could be ascribed to their LPS, which, apart from the poor immunostimulatory capacity, acted as antagonists of E. coli-derived LPS. The LPS was, however, still capable of inducing increased endocytosis in the DCs and activating the proteases in the LAL assay.
The poor immune stimulating properties of the intact bacteria studied here were evident as they only induced a weak production of the IL-12, IL-10 and TNF-α and, additionally, a weak induction of the surface markers CD40 and CD86, involved in T-cell activation. IL-6 production, however, was induced to the same level as in DCs stimulated with E. coli LPS or the other marine bacteria tested. The lack of immune stimulation coincided with a potent inhibition of E. coli LPS-induced IL-12, indicating that microbial compounds present on the bacteria were able to directly or indirectly abrogate the activation of the DCs by E. coli LPS. Some pathogenic bacteria have been shown to be able to escape the immune recognition by repression of the immune response through the binding of carbohydrate moieties to receptors, typically of the C-type lectin class, thus inducing a response counteracting the immune-stimulating effect of LPS.35,36 Moreover, it has been shown that LPS from some Gram-negative species are poor immune stimulators.22,37 When testing the immune-modulating activity of the medium harvested upon growth of Ps. luteoviolacea, is was evident that the fresh medium contained compounds with immune suppressing activity, but without immune stimulatory capacity, while the SM held immune-suppressing, as well as activating, activity. It is well established that yeast extract present in the growth medium contains β-glucans and we have previously shown that yeast β-glucan is capable of inhibiting the E. coli-induced IL-12 but unable to stimulate DC activation per se. 38 Of note, in the SM, however, compounds capable of slightly stimulating IL-10, IL-6 and TNF-α production were present. These were HMM compounds and exhibited IL-12 inhibiting activity, greatly exceeding that of fresh medium.
LPS is known to be shed from Gram-negative bacteria into the media and LPS is most often the main ligand in the recognition of Gram-negative bacteria by immune cells. 17 By isolating LPS from both the whole bacteria of two Pseudoalteromonas strains, Ps. luteoviolacea and Ps. ruthenica, we established LPS as the cause of inhibition of the E. coli LPS-induced IL-12. This was further substantiated by the use of the TLR4 transfected HEK cells, where the involvement of other PRRs could be ruled out and the role of LPS confirmed. The TLR4 binding lipid A part of LPS is a highly diverse molecule, with variations in both the number and the length of fatty-acid side chains, and in terminal phosphate residues; 39 these variations are critical for TLR4 signalling. The hexa-acylated lipid A from E. coli with side chains of 12–14 carbon atoms seems to be optimal for stimulation, while tetra- and penta-acylated lipid A are often poorly recognized by the MD2-TLR4-complex and do not lead to TLR4 dimerization, which is central for the induction of down-stream processes in the cell leading to DC maturation and cytokine production.9,22 We did not characterize the structures of the LPS or lipid A molecules from the Pseudoalteromonas strains tested in this study; however, the available structures of LPS molecules from five other Pseudoalteromonas strains, Ps. haloplanktis TAC 125, Ps. haloplanktis ATCC 14393, Ps. nigrifaciens IAM 13010, Ps. tetraodonis and Ps. haloplanktis TAB 23, show that they all contain penta-acylated lipid A.12–14,16 Additionally, a recent study showed that LOS from Ps. haloplanktis TAB 23 is penta-acylated and exhibits antagonistic activity towards the E. coli LPS-stimulated human monocytic cell line, THP-1. 16 Thus, it is likely that the LPS from the Pseudoalteromonas strains investigated in the present study are penta-acylated and that this renders them low stimulators and antagonists. Strikingly, we did not find that LPS from Pseudoalteromonas inhibited induction of TNF-α upon stimulation with E. coli in a dose-dependent manner, indicating that either other components in E. coli induce the majority of the TNF-α production or that LPS induces the TNF-α production through other pathways. We have obtained very similar results by stimulation of DCs with E. coli and LPS from E. coli (data not shown) and, moreover, as we show in TLR4-transformed HEK cells, that TLR4 ligation by E. coli LPS is antagonized by the Pseudoalteromonas LPS, we find it likely that E. coli LPS may induce TNF-α production in DCs through other pathways than through TLR4.
Despite the very similar antagonistic nature of the purified LPS from Ps. luteoviolaceae and Ps. ruthenica, respectively, the molecular mass patterns revealed by SDS-PAGE were highly distinct. While the Ps. luteoviolacea LPS exhibited a marked band around 30 ku, indicative of a high molecular mass O-antigen moiety, the Ps. ruthenica LPS showed lower MM bands, but only two as compared with a high number in E. coli LPS. However, both Pseudoalteromonas LPS showed marked lipid A bands of comparable size and concentration that were markedly different from that of E. coli LPS. This indicates that the type of carbohydrate of the O-antigen, and possibly also the core carbohydrate, is of minor, or any, significance for the immune stimulating and antagonistic properties.
Remarkably, despite the lack of immune stimulating activity, the isolated Pseudoalteromonas LPS was as potent as E. coli LPS in inducing increased endocytosis in the DC. It was shown recently that the binding of LPS to CD14 is required for induction of endocytosis and, hence, uptake of dextran particles. 40 The lipid A part is recognized by CD14, which directs the further binding of the LPS molecule to MD2. 41 To this end it is also worth noticing that all Pseudoalteromonas stains exhibited smooth colony morphology when grown on agar plates. It has previously been shown that rough LPS of E. coli and Salmonella may bind to TLR4/MD2 and activate TLR4 dimerizaton without the assistance of CD14, while CD14 is indispensable in the induction of a TLR4-mediated response to smooth LPS. 42 Thus, even though the Pseudoalteromonas LPS is not able to bind to MD2/TLR4 in a way that leads to TLR4 dimerization, it may still bind to CD14 and thus induce endocytosis and the transfer of LPS to the MD2 protein.
It is also noteworthy that the Pseudoalteromonas LPS was able to activate the enzymatic process in the LAL assay. The endotoxin activity determined by the LAL assay corresponded nicely to the antagonistic capacity of the same samples, for example compared with SM from Ps. luteoviolaceae. Ps. ruthenica SM exhibited a substantially weaker antagonistic activity both in DC and in the HEK cells. This corresponds well with the much lower concentration of LPS in the Ps. ruthenica SM. The activation of the serine protease factor C is dependent on the lipid A moiety in LPS; 43 hence, it must do so in a way that does not differentiate between structural differences, for example differences in acylation, of the lipid A moiety.
We used DCs to investigate the stimulatory and antagonistic properties of Pseudoalteromonas LPS. By ligation of innate receptors, DCs become activated and link the innate and adaptive immunity, Although DCs are likely to play only a minor role in septic shock and upon LPS stimulation display a cytokine profile distinct from that of LPS stimulated monocytes or neutrophils, we believe that all endocytosing cells expressing TLR4 may be equally valuable in assessing putative TLR4 antagonists.
In conclusion, we have shown that two strains of Pseudoalteromonas isolated from marine habitats owing to the structure of the LPS are poor immunostimulators with antagonistic properties. The LPS is still capable of inducing endocytosis in DCs and in activating the LAL endotoxin assay. LPS molecules from Pseudoalteromonas might, by their antagonistic activity, find use as inhibitors of LPS induced sepsis. However, in order to establish such potential, studies of the effects on human phagocytes, in particular of neutrophils, as well as in vivo effects of the Pseudoalteromonas LPS, need to be done.
Footnotes
Funding
This study was supported by the Programme Commision on Health, Food and Welfare under the Danish Council for Strategic Research. The present work was carried out as part of the Galathea 3 expedition under the auspices of the Danish Expedition Foundation. This is Galathea 3 contribution no. P92.
Acknowledgements
We acknowledge Abdellatif El Ghazi, Heidi Gertz Andersen and Henriette Vorsholt for help with LPS purification and SDS-PAGE analyses; and Søren Paludan for his kind donation of transfected TLR4 HEK293 cells. Also, huge thanks to Anni Mehlsen and Marianne Petersen for help with the experiments; and Cisse Hedegaard Prosby, Matthias Wietz and Jette Melchiorsen for helping with the marine bacteria.