
Abstract
Ovulation is a critical inflammation-like event that is central to ovarian physiology. IL-1β is an immediate early pro-inflammatory cytokine that regulates production of several other inflammatory mediators, such as cyclooxygenase 2 (COX)-2 and IL-8. NS-398 is a selective inhibitor of COX-2 bioactivity and thus this drug is able to mitigate the COX-2-mediated production of downstream prostaglandins and the subsequent inflammatory response. Here we have investigated the action of NS-398 using a human ovarian granulosa cell line, KGN, by exploring IL-1β-regulated COX-2 and IL-8 expression. First, NS-398, instead of reducing inflammation, appeared to further enhance IL-1β-mediated COX-2 and IL-8 production. Using selective inhibitors targeting various signaling molecules, MAPK and NF-κB pathways both seemed to be involved in the impact of NS-398 on IL-1β-induced COX-2 and IL-8 expression. NS-398 also promoted IL-1β-mediated NF-κB p65 nuclear translocation but had no effect on IL-1β-activated MAPK phosphorylation. Flow cytometry analysis demonstrated that NS-398, in combination with IL-1β, significantly enhanced cell cycle progression involving IL-8. Our findings demonstrate a clear pro-inflammatory function for NS-398 in the IL-1β-mediated inflammatory response of granulosa cells, at least in part, owing to its augmenting effect on the IL-1β-induced activation of NF-κB.
Introduction
Inflammation plays an important role in the host defense system that occurs in response to internal or external stimuli and has the function of counteracting the insults exerted by these stimuli.1,2 As a fine-tuned acute inflammation consequence lasting for a short period of time, it is responsible for innate immunity and/or humoral immunity, both of which have therapeutic significance to the host; when inflammation becomes chronic and lasts for a long period of time, the outcome may result in pathogenic consequences to the body. 3
Ovulation is a key event in ovarian cycle that involves a number of regulatory networks. It has been well recognized that several aspects of ovulation resemble an inflammation-like reaction. Accumulated studies have demonstrated that various inflammatory factors are involved in ovulation. For example IL-1β, cyclooxygenase-2 (COX-2) and IL-8 have all been proposed to be of particular significance during ovulation.4–6 In the COX-2 knockout animals that are unable to ovulate, the administration of gonadotropin and IL-1β is able to restore the ability to ovulate to a level comparable with that of wild type animals. 5 The direct injection of IL-1β intra-follicularly in equine-dominant follicles is able to induce the ovulation process. 4 In addition, IL-1β has been reported to up-regulate the expression and production of several other inflammatory mediators, including COX-2 and IL-8.7,8 The critical role of COX-2 in ovulation is well established and this protein is known to regulate the rate-limiting step of ovulation, namely the production of prostaglandins (PGs), which are central to the ovulatory process. 5 IL-8 is a chemotactic cytokine that helps to recruit and activate neutrophils. IL-8 was detected in the follicular fluid obtained from in vitro fertilization patients. Furthermore, the secretion of IL-8 from cultured granulosa cells has been shown to be enhanced by IL-1β.9,10 When the immature rats were injected with pregnant mare serum gonadotropin followed by the administration of human IL-8, the exogenous IL-8 was able to induce an increase in follicular growth that was able to reach the same level as that produced by the luteinizing hormone surge. 11 The administration of human chorionic gonadotropin (hCG) to induce ovulation in rabbits has also been demonstrated to be significantly attenuated by treatment with anti-rabbit IL-8 antiserum. 6 Thus, IL-8 also has begun to be regarded as an important player in ovulation.6,7,9–11 In addition, studies have also proposed that IL-1β may be able to induce the inharmonious expression/secretion of growth factors such as COX-2, various PGs and IL-8 in a micro-environmental manner; such an effect may lead to various ovarian diseases. 12
A variety of different types of non-steroid anti-inflammatory drugs (NSAIDs) have been developed and these are commonly used to treat inflammatory disorders; they do this by blocking the COX activity of the various COX enzymes, which then prevent the synthesis of PGs. 13 However, several studies have indicated that regular NSAID use may increase the risk of gastrointestinal ulceration owing to its non-selective inhibition of COX activity. 14 COX controls the rate-limiting step that catalyzes the biosynthesis of various PGs from the arachidonates, which are then released from the plasma membrane. 15 In addition to the constitutively expressed isoform of COX, namely COX-1, there is also COX-2, which is an inducible isoform, and this enzyme is usually not detectable in most tissues. The expression of COX-2 can be dramatically induced by a variety of stimuli; these include various cytokines, various growth factors and LPS.16–18 NS-398 is a COX-2-selective inhibitor with IC50 values for human recombinant COX-1 and COX-2 of 75 and 1.77 µM, respectively, and IC50 values for ovine COX-1 and COX-2 of 220 and 0.15 µM, respectively.19,20 In addition to being able to inhibit COX-2 bioactivity and thus block the production of PGs, NS-398 also has been reported to suppress cell proliferation and to promote apoptosis in vascular smooth muscle cells and colorectal carcinoma cells.21–23 If we examine ovarian physiology in relation to ovulation, the effect of the in vivo administration of NS-398 following an ovulatory dose of hCG in the proestrous rats has been found to result in reduced PGE2 production, as well as a decreased ovulation rate. 24 Another study in cattle that had received ultrasound-mediated intra-follicular injection of NS-398 demonstrated a reduction in gonadotropin-releasing hormone injection-mediated intra-follicular PGE2 synthesis and follicle rupture. 25
The aim of this study was to decipher how NS-398 impacts on the various IL-1β-mediated inflammatory cascades in human granulosa cells by monitoring the COX-2/PGE2 and the IL-8 pathways, which are two critical components associated with inflammation and are also involved in ovarian physiology. Furthermore, we also aimed to clarify the signaling mechanisms that are targeted by NS-398 and are involved in the IL-1β-regulated production of COX-2/PGE2 and IL-8.
Materials and methods
Reagents
FBS was obtained from HyClone (Logan, UT, USA). The COX-2 selective inhibitor NS-398 was purchased from Sigma Chemicals (St. Louis, MO, USA). Reverse transcriptase and Taq polymerase were purchased from Promega (Madison, WI, USA). The IL-8 ELISA kit and IL-8 Abs were purchased from R&D Systems (Minneapolis, MN, USA). The mouse anti-COX-2 mAbs were purchased from Cayman Chemical (Ann Arbor, MI, USA). The PGE2 enzyme immunoassay kit was obtained from Assay Designs (Ann Arbor, MI, USA). Rabbit polyclonal anti-phospho-p38 (Thr180/Tyr182) Abs, rabbit polyclonal anti-p38 Abs, rabbit anti-phospho-JNK (Thr183/Tyr185) Abs, rabbit monoclonal anti-JNK Abs, rabbit polyclonal anti-phospho-p44/42 (Thr 202/Tyr204) Abs, rabbit anti-p44/42 Abs, anti-p21waf-1/cip-1 Abs and anti-p27kip-1 Abs were obtained from Cell Signaling (Danvers, MA, USA). Rabbit polyclonal Abs against human NF-κB p65 were purchased from Thermo Scientific (Fremont, CA, USA). Mouse monoclonal Abs against human histone H1 were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Mouse anti-α-tubulin mAbs were purchased from Sigma Chemicals. The alamarBlue assay kit was purchased from Invitrogen (Carlsbad, CA, USA). Unless otherwise specified, all the other chemicals and reagents used in this project were purchased from Sigma Chemicals.
Cell culture
The human ovarian granulosa cell line KGN was maintained in a 1:1 mixture of DMEM and nutrient mixture F-12 (Ham) (DMEM/F-12) with 10% FBS, 2 g/l sodium bicarbonate, 100 U/ml penicillin and 100 µg/ml streptomycin; the cells were cultured in an atmosphere of 5% CO2 at 37℃. 26 The cells were plated on the previous day to allow a 16–18 h (overnight) seeding period prior to the various treatments, which were carried out the following day.
Western blotting
Overnight-plated KGN cells were treated with IL-1β (10 ng/ml) in the absence or presence of NS-398 (5, 10, 25 µM) for 24 h. The protein concentrations of the harvested total cell lysates, the nuclear proteins and the cytosolic proteins were determined using Bio-Rad protein assay reagent (Bio-Rad, Hercules, CA, USA). Total protein concentrations were adjusted with SDS-PAGE loading buffer and this was followed by heating to 100℃ for 10 min. The samples were then subjected to regular Western blotting to determine the expression level of the various proteins of interest. Specifically, samples containing equal amounts of proteins (50 µg) were separated by 10% SDS-PAGE, and the proteins were then transferred onto a nitrocellulose membrane, blocked with 5% skimmed milk for 1 h and finally incubated overnight with various specific Abs. The probe Abs consisted of anti-COX-2 Abs (1:1000), anti-IL-8 Abs (1:500), anti-phospho-p38 Abs, anti-total-p38 polyclonal Abs (1:1000), anti-phospho-JNK Abs, rabbit anti-total-JNK Abs (1:1000), anti-phospho-ERK Abs, rabbit anti-total-ERK polyclonal Abs (1:2000), anti-phospho-Akt Abs, anti-total-Akt mAbs (1:1000), anti-NF-κB/p65 (Rel A) polyclonal Abs (1:500), anti-histone H1 mAbs (1:500), anti-p21 Abs, anti-p27 Abs (1:1000) and mouse anti-α-tubulin mAbs (1:5000). After probing with these Abs individually, this was followed by incubation for 2 h with the corresponding secondary Abs, such as sheep anti-mouse IgG HRP-coupled Abs (1:5000) and donkey anti-rabbit IgG HRP-coupled Abs (1:2500). After this, each membrane was developed, which was followed by exposure to film and then the bands of interest on the film were quantified using ImageQuant 5.2 software (Molecular Dynamics, Sunnyvale, CA, USA).
Measurement of PGE2 and IL-8 production
Overnight-plated KGN cells were treated with IL-1β (10 ng/ml) in the absence or presence of NS-398 for 24 h. The cultured medium was then collected and the IL-8 concentration was analyzed using a commercial available ELISA kit according to the instructions from the manufacturer (R&D Systems). Similarly, the PGE2 concentration was determined using an EIA kit (Assay Designs).
Semi-quantitative RT-PCR and quantitative real-time PCR
Overnight-plated KGN cells were treated with IL-1β (10 ng/ml) in the absence or presence of NS-398 for 6 h, which was followed by extraction of total cellular RNA using Tri-Reagent (Sigma Chemicals), according to the manufacturer’s instructions. The isolated RNA samples were re-suspended in RNase-free diethylpyrocarbonate-treated water and then each sample underwent two-step semi-quantitative RT-PCR in order to measure the levels of mRNAs encoding COX-2, IL-8, GAPDH, and β-actin. The primer sequences used in PCR were: IL-8 sense 5’-ACTTCCAAGCTGGCCGTGGCT-3’, antisense 5’-TCACTGGCATCTTCACTGATT-3’ to give a 345-bp product; COX-2 sense 5’-GAA TGGGGTGATGAGCAGTT-3’, antisense 5’-CAGAAGGGCAGGATACAGC-3’ to give a 561-bp product; GAPDH sense 5’-ATCACCATCTTCCAGGAGCG-3’, antisense 5’-CCTGCTTCACCACCTTCTTG-3’ to give a 574-bp product; and β-actin sense 5’-GGCACCACACCTTCTACAAT-3’, antisense 5’- CGTCATACTCCTGCTTGCTG-3’ to give an 834-bp product. In brief, 1 µg total RNA from each sample was used to perform the reverse transcription. In order to detect the cDNA content of each sample, 2 µl cDNA from the RT-PCR reaction was added into the PCR reaction tube and mixed with 10× PCR buffer, 0.5 mM dNTP, 0.5 µM forward and reverse primers (MDBio Inc., Taipei, Taiwan), and 0.2 U Taq DNA polymerase. Amplification was carried out using a Program Temp Control System PC 818 (Astec Technology, Fukuoka, Japan). The amplification of the DNA products started with incubation at 95℃ for 2 min in order to denature the DNA present; this was followed by 35 cycles (30 cycles for GAPDH and β-actin) of PCR, namely 95℃ for 30 s to denature the template, 57℃ for 30 s to allow annealing of the primers to the single-stranded DNA template and 72℃ for 1 min to elongate the DNA product. The reaction ended with a final extension for 10 min at 72℃. The PCR products were analyzed by electrophoresis using a 2% agarose gel containing 1 µg/ml ethidium bromide. The final cDNA yields were then determined from the amplified DNA signals by comparing them against the internal control, GAPDH or β-actin. The DNA signals were captured and analyzed by ImageQuant 5.2 software. In parallel, quantitative real-time PCR was also performed by Microarray & Gene Expression Analysis Core Facility of the National Yang-Ming University VGH Genome Research Center using an ABI Prism 7900HT System and a TaqMan Gene Expression Assay (Applied Biosystems, Carlsbad, CA, USA). This used a 96-well micro-fluidic card preloaded with optimized probes and matching primers from Applied Biosystems; these primers targeted human COX-2 (Hs01573469_m1) and human IL-8 (Hs00174103_m1). All samples were analyzed in triplicate and endogenous human GAPDH was used as the internal control.
Determining cell proliferation and cell cycle distribution
KGN cells plated at 10,000 cells/well in a 96-well plate format overnight and were then washed three times with serum-free medium, which was followed by treatment with IL-1β (10 ng/ml) in the absence or presence of NS-398 for 24 h or 48 h. Cell proliferation was assessed by the alamarBlue assay (Invitrogen), which involves the addition of a fluorogenic redox indicator to the cell culture. Via the reduction reactions of metabolically active cells, non-fluorescent resazurin is reduced to bright-red fluorescent resorufin. 27 Four h before termination of treatment, alamarBlue dye (Invitrogen) was added and the mixture was incubated for 4 h at 37℃. The culture medium containing the alamarBlue dye was then removed from wells and transferred into a black 96-well plate (OptiPlate-96F; Perkin Elmer, Waltham, MA, USA), and this was subjected to spectrofluorometry (Varioskan Flash Spectral Scanning Multimode Reader, Thermo Electron, Waltham, MA, USA) with excitation at 550 nm and emission at 590 nm.
To determine the cell cycle distribution profile, KGN cells were plated in six-well plate at a density of 4 × 105 cells/well overnight and then the cells were serum-starved for 24 h before the various treatments. After termination of treatment, the cells were harvested by trypsinization and then washed with cold PBS. Next, the cell pellet was fixed with 75% ethanol for 15 min, which was followed by the incubation with propidium iodide. For each treatment group, 10,000 cells were analyzed by Becton Dickinson FACSCalibur flow cytometer (BD Science, Franklin Lakes, NJ, USA) and the proportions of cells in G0/G1, S, and G2/M phases were estimated using CXP software analysis (Beckman Coulter, Brea, CA, USA).
Statistical analysis
Experimental data are expressed as the mean ± SEM. The results were analyzed by ANOVA, which was followed by the least-significant difference test in order to compare the differences between each of the treatment groups and the control group. Differences with a P-value < 0.05 were considered to be statistically significant.
Results
NS-398 augmentation of IL-1β-induced protein expression and mRNA expression of COX-2 and IL-8, as well as the secretion of IL-8
We first examined whether NS-398 affects the IL-1β-regulation of intracellular COX-2 and IL-8 expression, as well as PGE2 and IL-8 secretion. KGN human ovarian granulosa cells were exposed to IL-1β (10 ng/ml) in the presence or absence of NS-398 (5, 10, 25 µM) for 24 h. On treatment with IL-1β, a three-fold increase in COX-2 and an eight-fold increase in IL-8 expression were noted (Figure 1a, b). Interestingly, NS-398 at a dose of 10 or 25 µM dramatically enhanced the IL-1β-induced expression of COX-2 and IL-8 (Figure 1a, b). IL-1β resulted in a significant secretion of PGE2 and IL-8, namely a four-fold increase in PGE2 (∼1000 pg/ml; Figure 1c) and a 200-fold increase in IL-8 (∼100 ng/ml; Figure 1d). NS-398 at all concentrations (5, 10, 25 µM) was found to inhibit IL-1β-induced PGE2 secretion (Figure 1c), but NS-398 at 10 or 25 µM further enhanced IL-1β-mediated IL-8 secretion (Figure 1d). In addition, NS-398 treatment in the absence of IL-1β did not appear to affect significantly the expression of COX-2 and IL-8 or the secretion of PGE2 and IL-8 (Figure 1).
Augmentation of IL-1β-induced COX-2 and IL-8 expression and IL-8 secretion by NS-398 in KGN ovarian granulosa cells. Overnight-plated KGN cells were untreated, treated with NS-398 alone (25 µM) or treated with IL-1β (10 ng/ml) in the absence or presence of NS-398 (5, 10, 25 µM) for 24 h. The intracellular (a) COX-2 and (b) IL-8 protein expression levels and the resultant (c) PGE2 and (d) IL-8 concentrations in the cultured media were determined by Western blotting assay and ELISA, respectively. The results are expressed as mean ± SEM from four separate experiments. *P < 0.05 compared with control treatment; #P < 0.05 compared with IL-1β treatment.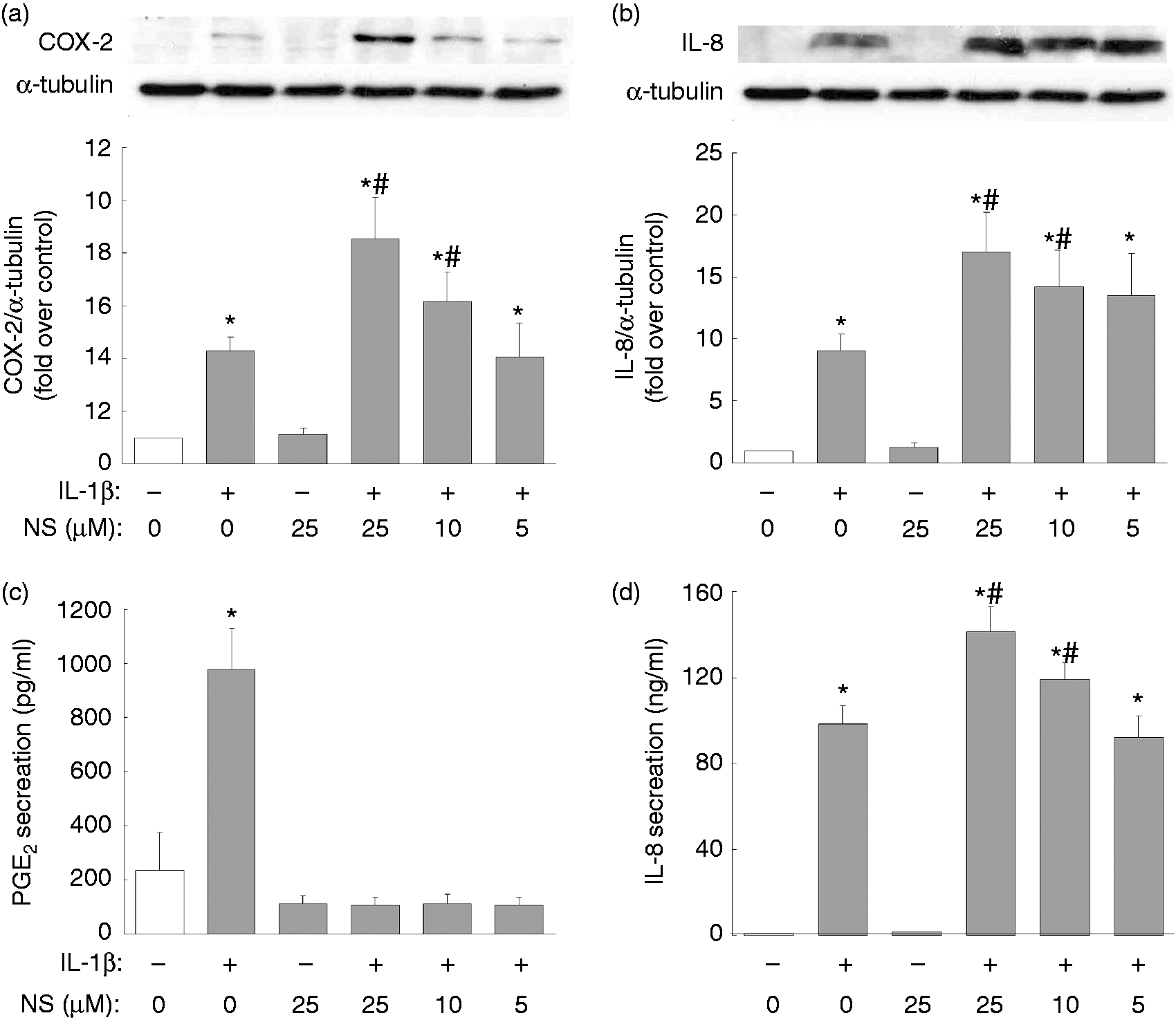
NS-398 promotes IL-1β-induced COX-2 and IL-8 expression at the mRNA level
NS-398 regulation of IL-1β-mediated protein expression of COX-2 and IL-8 may occur at either the mRNA or the protein level. We next evaluated whether NS-398 affects the protein stability of the IL-1β-induced COX-2 and IL-8 proteins. To do this, we used cycloheximide to block de novo protein synthesis and we found that NS-398 did not appear to have any effect on the stability of the COX-2 or IL-8 proteins (data not shown). Thus, in order to clarify whether regulation of COX-2 and IL-8 by IL-1β and NS-398 occurs at the transcriptional stage, we determined the mRNA expression levels of COX-2 and IL-8 after IL-1β (10 ng/ml) incubation with or without NS-398 (5, 10 or 25 µM) for 6 h by classical RT-PCR. Expression levels of COX-2 and IL-8 were found to be low in the control or the NS-398 alone treatment group, while IL-1β alone treatment resulted in an elevation of the COX-2 and IL-8 mRNA levels (Figure 2a, b). Furthermore, the addition of NS-398 at 10 µM or 25 µM produced a significant further increase in the IL-1β-induced expression of COX-2 and IL-8 mRNA (Figure 2a, b). In parallel, a real-time PCR analysis was also carried out and this resulted in a similar finding that the IL-1β-mediated elevation of both COX-2 and IL-8 mRNA was further enhanced by treatment with NS-398 (Figure 2c, d).
Enhancement of IL-1β-induced COX-2 and IL-8 mRNA expression by NS-398. Overnight-plated KGN cells were untreated, treated with NS-398 alone (25 µM) or treated with IL-1β (10 ng/ml) in the absence or presence of NS-398 (5, 10, 25 µM) for 6 h. (a, c) COX-2 and (b, d) IL-8 mRNA expression levels were determined by (a, b) RT-PCR and (c, d) quantitative real-time PCR. The results are expressed as mean ± SEM from (a, b) four or (c, d) three separate experiments. *P < 0.05 compared with control treatment; #P < 0.05 compared with IL-1β treatment.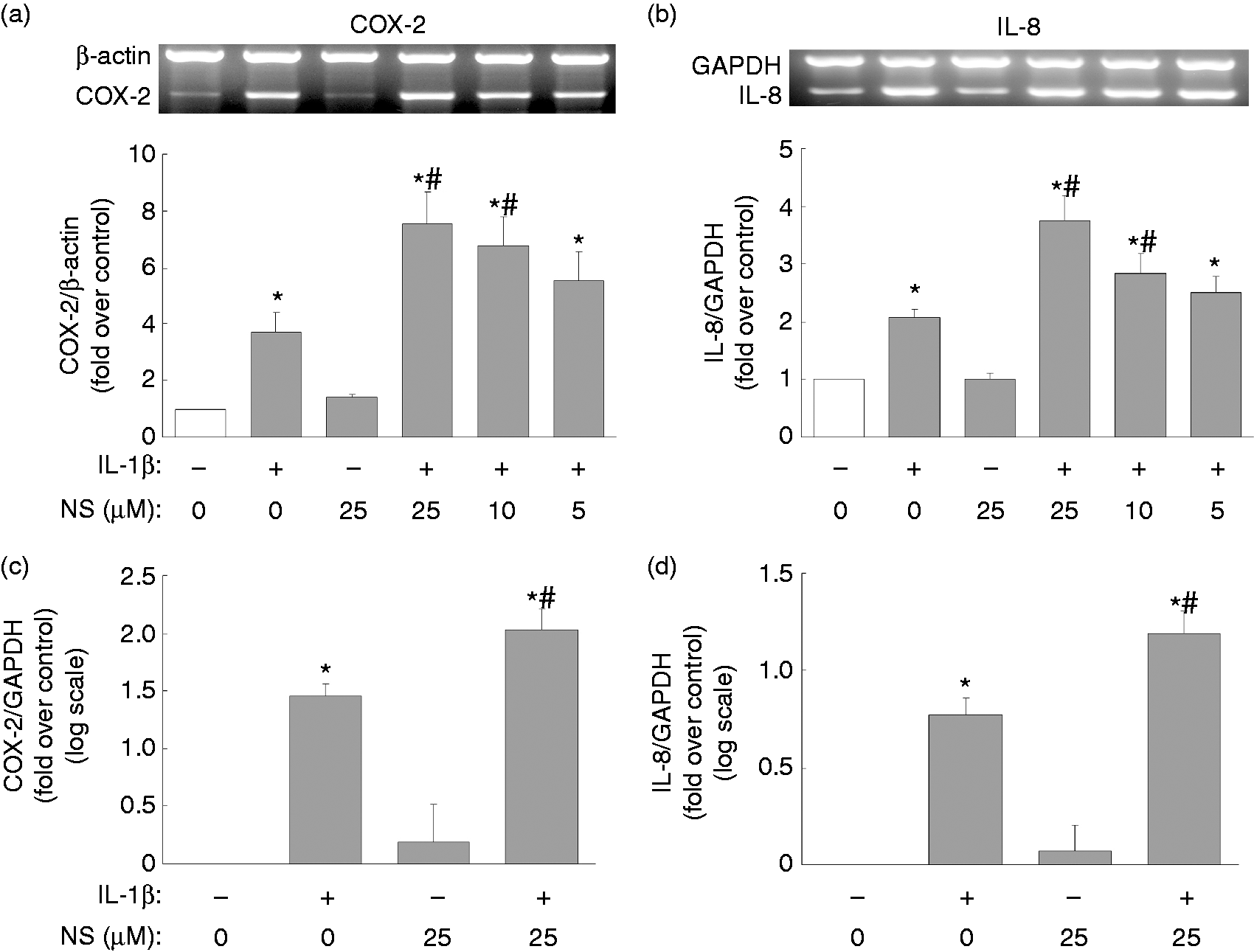
The promotion by NS-398 of IL-1β-induced NF-κB activation in relation to IL-1β-mediated COX-2 and IL-8 regulation
Previous studies have reported that MAPKs and NF-κB may act upstream and mediate expression of various inflammatory molecules.
28
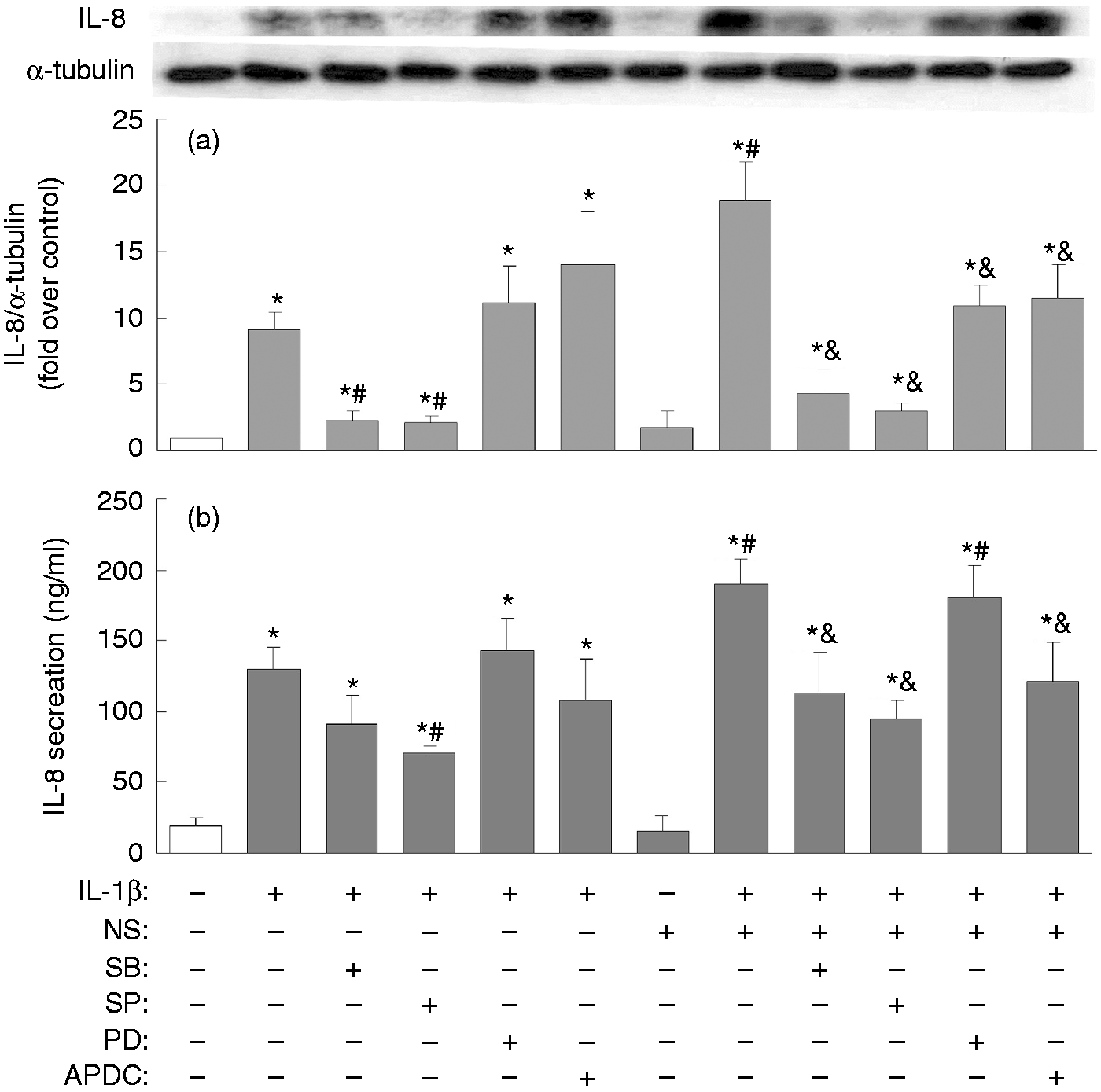
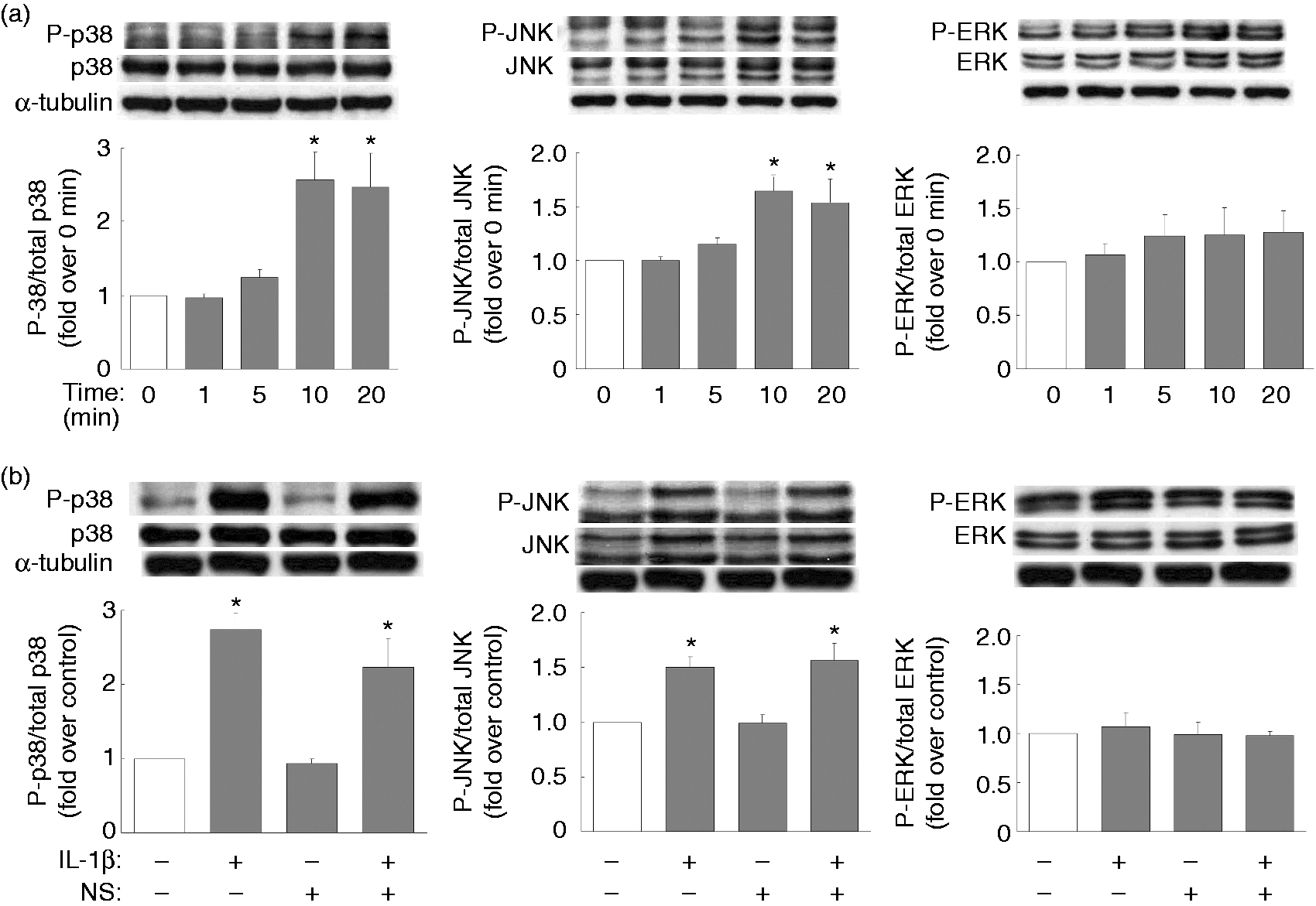
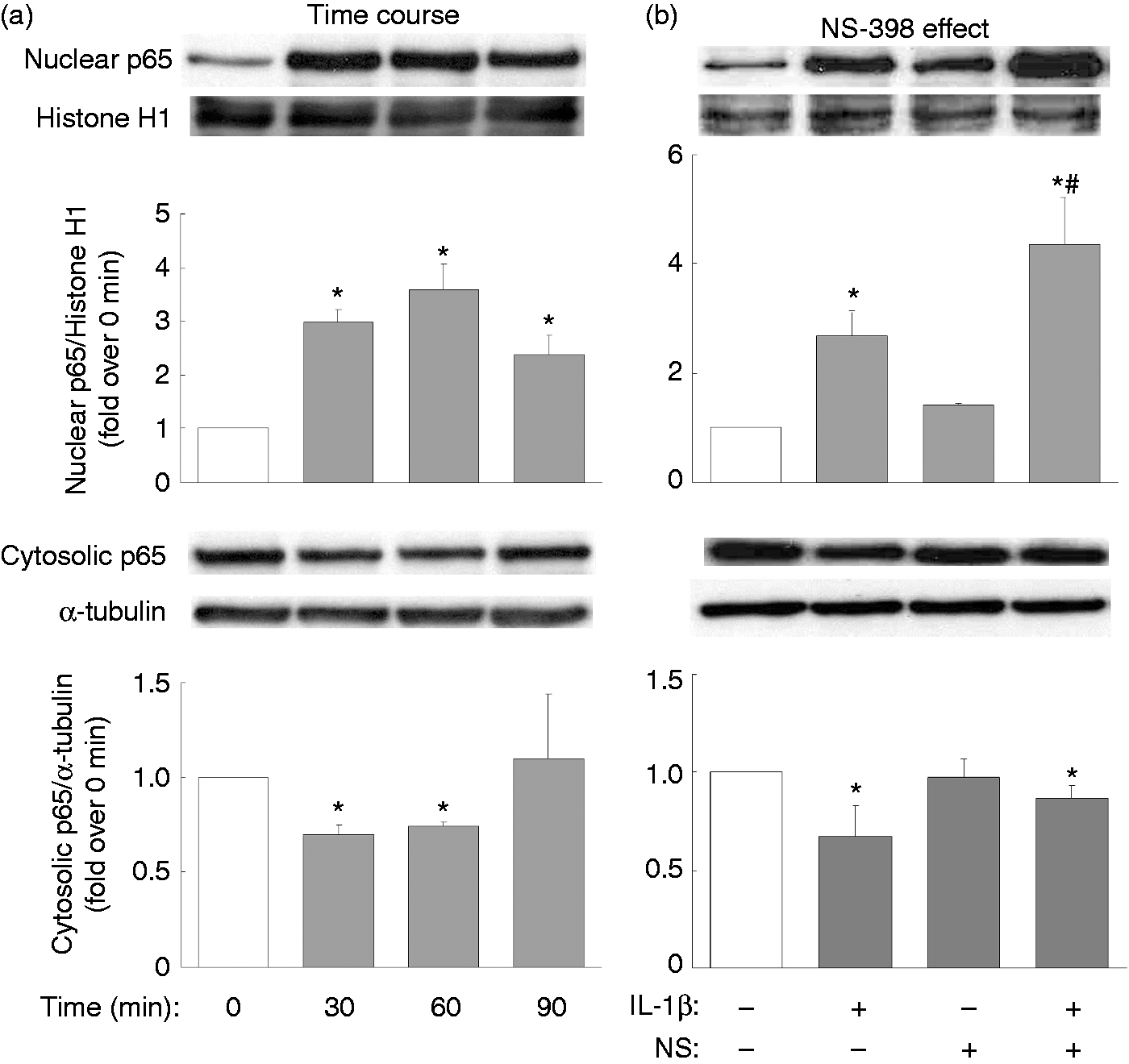
Thus, inhibitors of the various MAPKs (p38, JNK, ERK) and NF-κB were used to clarify in granulosa cells the signaling pathways post-IL-1β activation and the induction of COX-2 and IL-8. IL-1β-mediated COX-2 expression was found to be suppressed by inhibitors of p38 (SB2030580), of JNK (SP600125) and of NF-κB (APDC) (Figure 3); however, inhibitors of all MAPKs, as well as NF-κB, appeared to attenuate the combined IL-1β-mediated and NS-398-mediated COX-2 expression (Figure 3). Specifically, IL-1β-mediated IL-8 expression was attenuated by inhibitors of p38 and JNK, while IL-1β-mediated IL-8 secretion was only reduced by the JNK inhibitor (Figure 4). Furthermore, the inhibitors of all MAPKs, as well as of NF-κB, appeared to attenuate the combined IL-1β and NS-398 mediation of IL-8 expression, but only inhibitors of p38, JNK, and NF-κB were able to decrease effectively the combined IL-1β and NS-398 mediation of IL-8 secretion (Figure 4). It has been noted in many cell types that IL-1β is able to activate the MAPK signaling pathways, as well as the NF-κB signaling pathway, namely IL-1β-induced activation of MAPK (phosphorylation) and NF-κB (translocation from cytosol to nucleus), both of which then lead to induction of various downstream target genes.29,30 In this context, the effect of NS-398 on MAPK phosphorylation and on NF-κB translocation were monitored in KGN cells. We noted that IL-1β significantly induced p38 and JNK phosphorylation at 10 and 20 min (Figure 5a), and NS-398 did not appear to affect IL-1β-induced p38 or JNK or ERK phosphorylation (Figure 5b). However, IL-1β treatment for 30, 60 or 90 min strikingly increased translocation of p65 from cytosol to the nuclear compartment (Figure 6a); NS-398 alone caused a very minor p65 translocation into nucleus, but a combination of IL-1β and NS-398 was found to promote dramatically p65 nuclear translocation (Figure 6b).
Involvement of MAPKs and NF-κB in NS-398-regulated and IL-1β-regulated COX-2 expression. Overnight-plated KGN cells were pre-treated with various inhibitors of MAPKs, namely SB2030580 (SB, 20 µM) for p38, SP600125 (SP, 10 µM) for JNK, PD98059 (PD, 20 µM) for ERK and APDC (10 µM) for NF-κB for 1 h, followed by IL-1β (10 ng/ml) treatment or the combined IL-1β (10 ng/ml) and NS-398 (25 µM) treatment for an additional 24 h. This was followed by an analysis of the level of protein expression of COX-2 by Western blotting. The results are expressed as mean ± SEM from five separate experiments. *P < 0.05 compared with control treatment; #P < 0.05 compared with IL-1β treatment; &P < 0.05 compared with the combined IL-1β and NS-398 treatment. Involvements of MAPKs and NF-κB in NS-398-regulated and IL-1β-regulated IL-8 expression. Overnight-plated KGN cells were pre-treated with various inhibitors of MAPKs, namely SB2030580 (SB, 20 µM) for p38, SP600125 (SP, 10 µM) for JNK, PD98059 (PD, 20 µM) for ERK and APDC (10 µM) for NF-κB for 1 h, followed by IL-1β (10 ng/ml) treatment or the combined IL-1β (10 ng/ml) and NS-398 (25 µM) treatment for an additional 24 h. This was followed by analysis of the level of protein expression of IL-8 by (a) Western blotting and (b) the amount of IL-8 secreted into the cultured medium by ELISA. The results are expressed as mean ± SEM from five separate experiments. *P < 0.05 compared with control treatment; #P < 0.05 compared with IL-1β treatment; &P < 0.05 compared with the combined IL-1β and NS-398 treatment. NS-398 has no effect on IL-1β-induced MAPK phosphorylation. Overnight-plated KGN cells were (a) treated with IL-1β (10 ng/ml) for 0, 1, 5, 10, 20 min or (b) treated with IL-1β (10 ng/ml) or combined IL-1β (10 ng/ml) and NS-398 (25 µM) for 10 min. The phosphorylation patterns of MAPK p38, JNK or ERK were monitored by Western blotting. The results are expressed as mean ± SEM from (a) three or (b) four separate experiments. *P < 0.05 compared with (a) 0 min or (b) control treatment. Promotion of IL-1β-induced nuclear NF-κB translocation by NS-398. Overnight-plated KGN cells were treated with (a) IL-1β (10 ng/ml) for 0, 30, 60 or 90 min, or with (b) IL-1β (10 ng/ml) alone or combined IL-1β (10 ng/ml) and NS-398 (25 µM) for 60 min in order to monitor IL-1β-induced and NS-398-induced nuclear and cytosolic protein levels of the NF-κB p65 subunit by Western blotting. The results are expressed as mean ± SEM from (a) three or (b) four separate experiments. *P < 0.05 compared with (a) 0 min or (b) control treatment; #P < 0.05 compared with IL-1β treatment.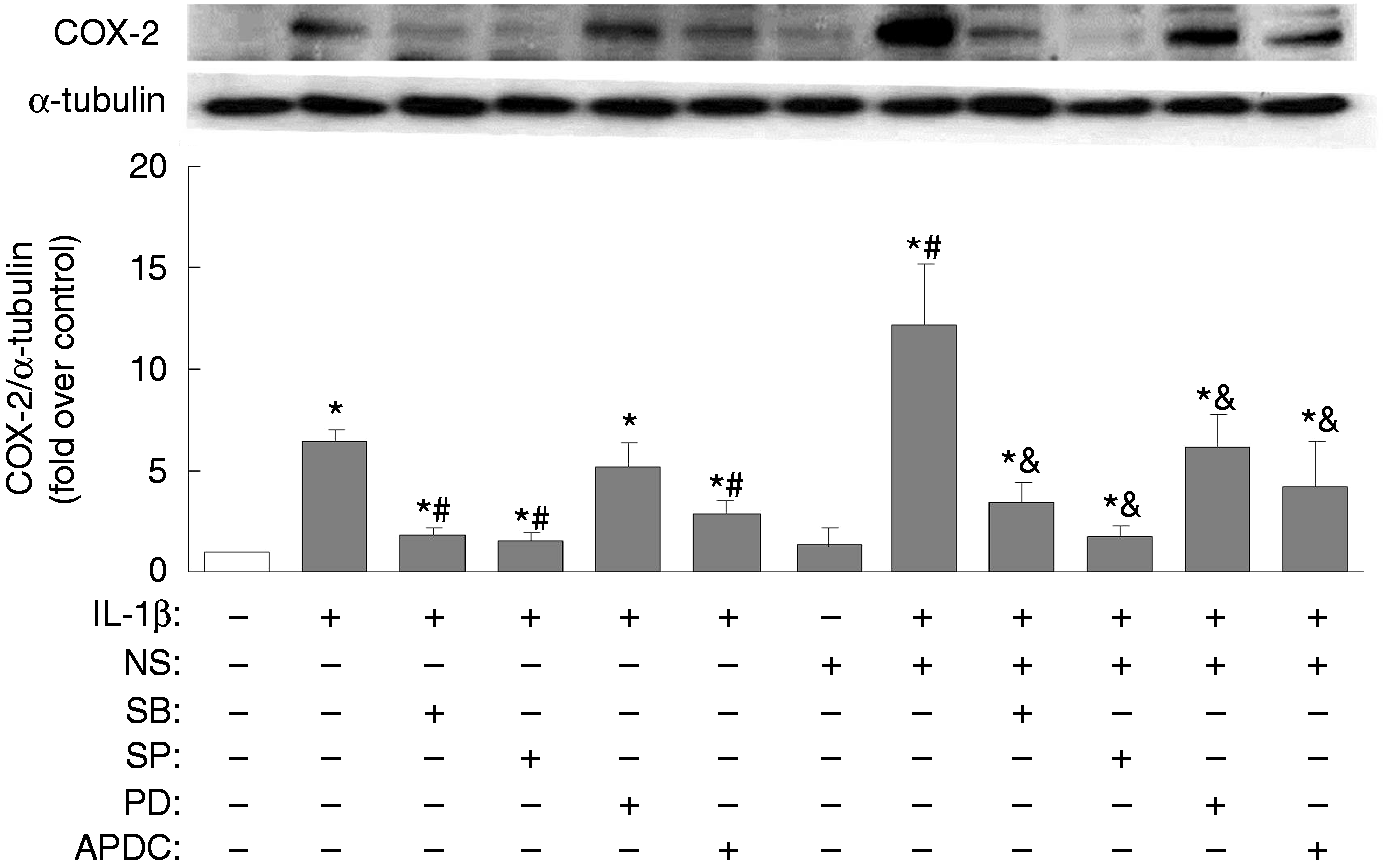
Promotion of cell cycle progression by combined IL-1β and NS-398 mediated IL-8 production
In addition to the induction of neutrophil recruitment, previous studies have demonstrated that IL-8 has the ability to promote cell proliferation.
31
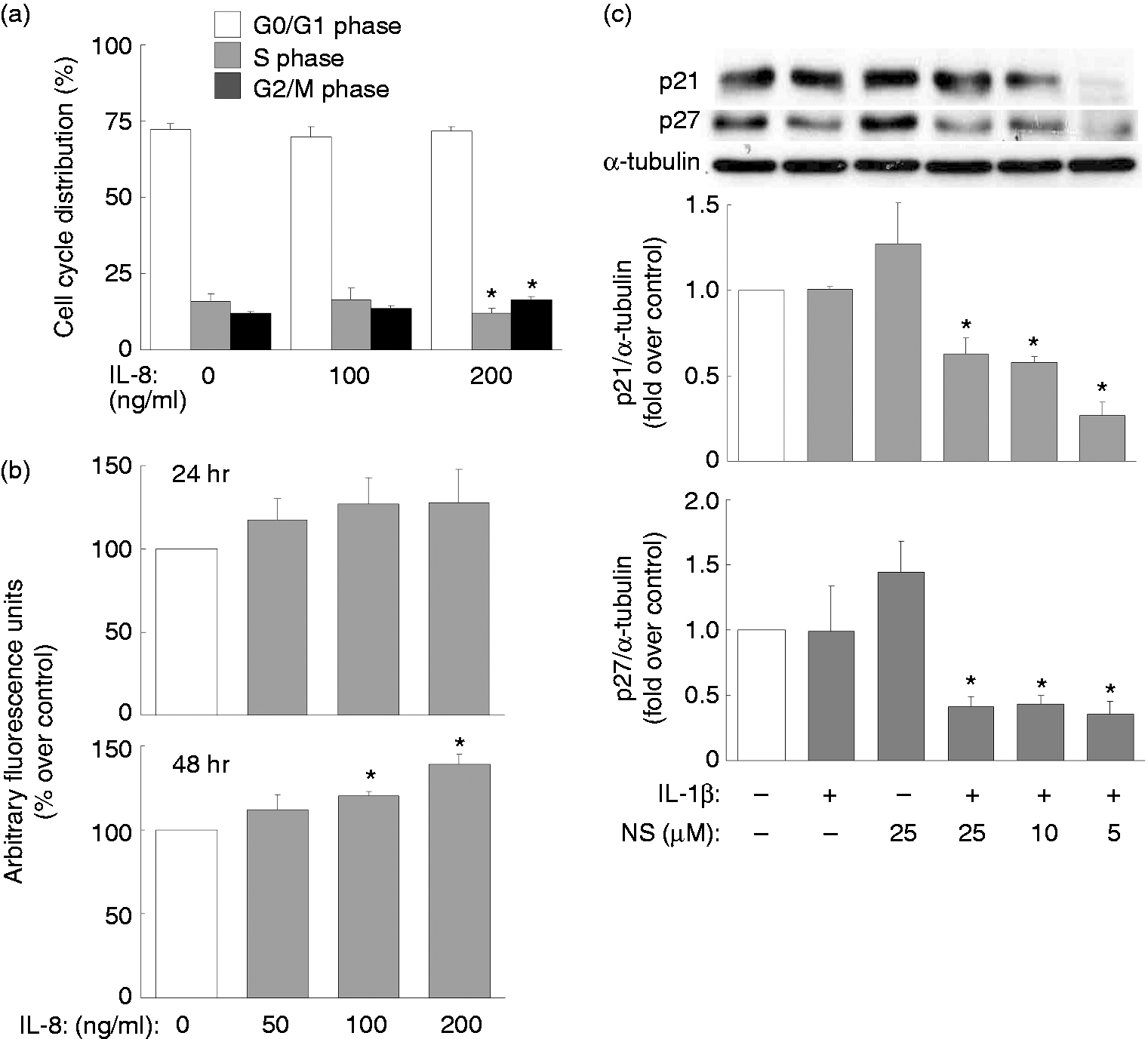
Thus, we next explored whether NS-398 is able to have an effect on cell proliferation under IL-1β stimulation. In order to do this the distribution of KGN cells across the various cell cycle stages were determined. A combination of IL-1β and NS-398 appeared to reduce the proportion of cells in S phase but to increase the proportion of cells in G2/M phase (Figure 7a). To further clarify the potential involvement of IL-8 in the combined IL-1β and NS-398 promotion of cell cycle progression, KGN cells were pretreated with control or specific IL-8 Abs before the addition of IL-1β and NS-398. Inclusion of the IL-8 Abs, but not the control Abs, effectively reversed the reduction in the proportion of cells in S phase and increased the proportion of cells in the G2/M phase brought about by the IL-1β and NS-398 combined treatment (Figure 7b). Furthermore, treatment with exogenous recombinant IL-8 was found to decrease the proportion of cells in S phase and to increase the proportion of cells in G2/M phase (Figure 8a), while IL-8 treatment (100 and 200 ng/ml) was found to induce KGN cell proliferation at 48 h (Figure 8b). In addition, previous studies have reported that the functions of p21waf-1/cip-1 and p27kip-1 involve the inhibition of cyclins and cyclin-dependent kinases, both of which are responsible for promoting cell cycle progression.
32
Thus, we went on to determine whether NS-398 in combination with IL-1β affects the expression of p21waf-1/cip-1 and p27kip-1. Indeed, NS-398 treatment at all doses (5, 10, 25 µM) in combination with IL-1β dramatically reduced the expression of both p21waf-1/cip-1 and p27kip-1, whereas NS-398 alone had no significant effect on the expression of either p21waf-1/cip-1 or p27kip-1 (Figure 8c).
Involvement of IL-8 in the combined IL-1β-mediated and NS-398-mediated cell cycle progression. Overnight-plated KGN cells were (a) untreated or treated with IL-1β (10 ng/ml) or NS-398 (25 µM) alone or in combination, or (b) treated with combined IL-1β and NS-398 in the presence of control or IL-8 Ab. After incubation, cells were incubated with propidium iodide, which was followed by flow cytometry to determination of cell-cycle distribution profiles. The results are expressed as mean ± SEM from four separate experiments. *P < 0.05 compared with control treatment within the same phase. Promotion of cell cycle progression and cell proliferation and the reduction of cell cycle regulators p21waf-1/cip-1 and p27kip-1 expression by combined IL-1β and NS-398 treatment. Overnight-plated KGN cells were treated with (a) IL-8 (0, 100, 200 ng/ml) for 24 h to determine the cell cycle distribution profile by flow cytometry, treated with (b) IL-8 (0, 50, 100, 200 ng/ml) for 24 or 48 h to determine cell proliferation index by alamarBlue assay, (c) untreated, or treated with NS-398 alone (25 µM) or with IL-1β (10 ng/ml) in the presence or absence of NS-398 (5, 10, 25 µM) for 24 h to monitor the expression of p21 and p27 by Western blotting. The results are expressed as mean ± SEM from (a, b) four or (c) three separate experiments. *P < 0.05 compared with control treatment.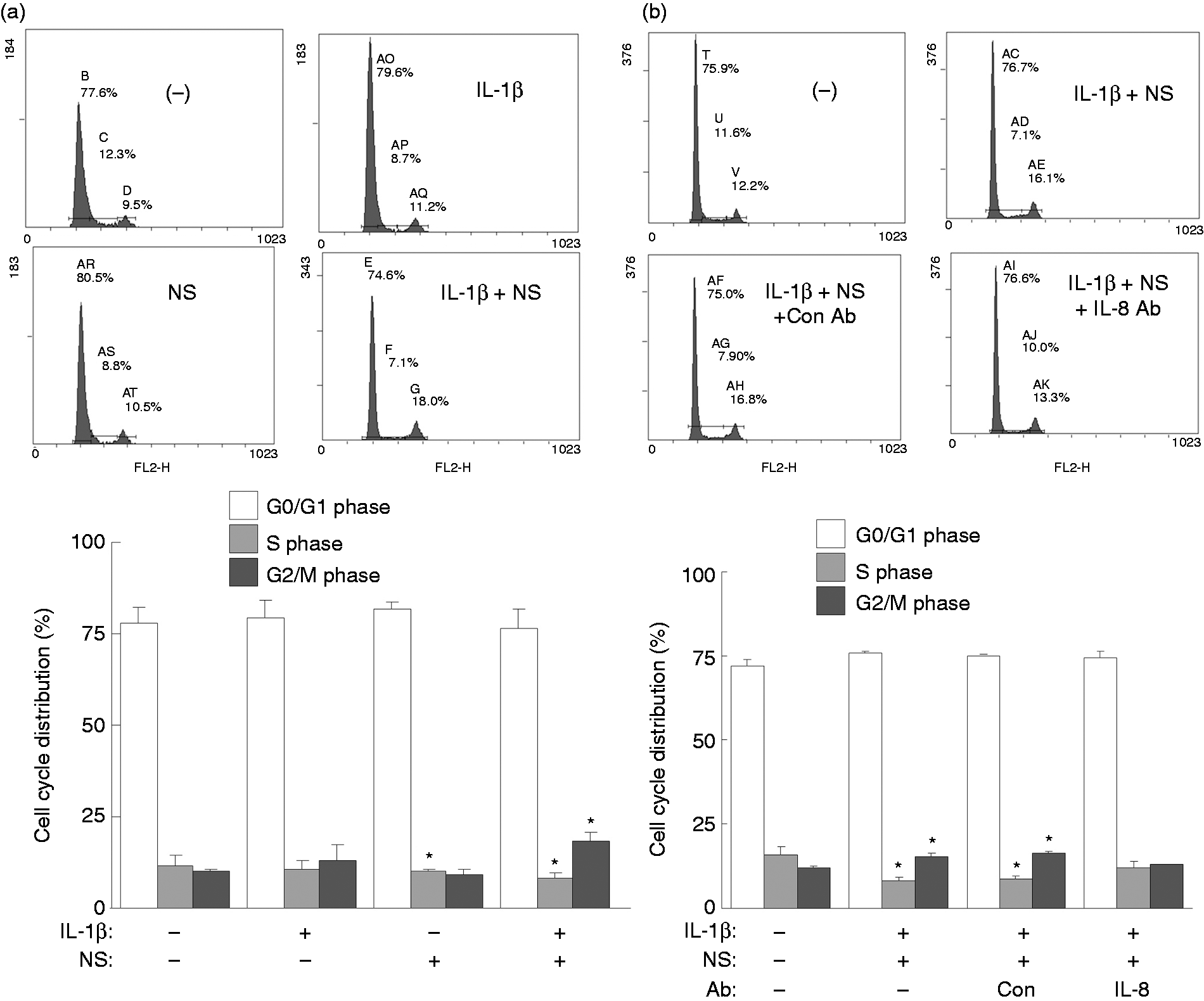
Discussion
Our results demonstrate that mRNA and protein expression of COX-2 and IL-8 genes are induced by IL-1β. NS-398, a selective COX-2 inhibitor, instead of attenuating the inflammation-inducing outcome of IL-1β as expected, further enhances the combined IL-1β and NS-398 mediation of IL-8 expression that leads to cell cycle progression and cell proliferation. Induction of COX-2 and IL-8 expression by IL-1β involves, at least in part, the MAPK pathways (p38, ERK and JNK) and the NF-κB pathway and NS-398 would seem to target the IL-1β-mediated activation of NF-κB, which subsequently up-regulates IL-1β-mediated COX-2 and IL-8 production.
In supporting the significance of inflammation to the ovarian system, our findings demonstrated in granulosa cells that IL-1β-induced IL-8 production is an important mediator of ovarian granulosa cell proliferation (Figures 7 and 8). This is consistent with previous studies demonstrating that IL-1β was able to induce IL-8 in human granulosa cells and to up-regulate a neutrophil chemoattractant IL-8 family CINC/GRO in rat granulosa cells.9,10,33 The cell cycle progression induced by a combination of IL-1β and NS-398 was significantly attenuated by anti-IL-8 Ab treatment (Figure 7), which strongly supports the hypothesis that the combined effect of IL-1β and NS-398 may induce IL-8 production by granulosa cells and that the subsequent increase in IL-8 then promotes granulosa cell proliferation, possibly in an autocrine manner. More importantly, in this study we demonstrated that when granulosa cells are treated with NS-398, a selective COX-2 inhibitor, although this drug essentially suppresses COX-2 activity and therefore blocks IL-1β-induced PGE2 secretion (Figure 1c), the expression of COX-2 at the mRNA and protein levels mediated by IL-1β is further enhanced by NS-398 (Figures 1 and 2). Notably, NS-398 alone did not affect either COX-2 or IL-8 expression (Figures 1 and 2), which suggests that NS-398 may target existing IL-1β-activated signaling pathways. This implies that, although the original definition of NS-398 was a drug with anti-inflammatory features, by inhibition of COX-2 bioactivity, this drug may have potentially unidentified effects involving amplification of the inflammatory process via other inflammatory molecules, such as COX-2 and IL-8.
When considering the cellular target sites by which NS-398 is able to modulate IL-1β-induced COX-2 expression and IL-8 expression and subsequent secretion, there is a panel of potential sites. For example, the interaction may occur at the cell surface where the agonist binds its receptor or, alternatively, within one or more intracellular compartments, with a range of steps, such as transcription, post-transcription, translation, post-translation and secretion as potential targets. As NS-398 can easily penetrate the cell membrane to target to COX-2, it seems very likely that it may interact with one or more intracellular molecules within cells. 34 In fact, previous studies have indicated that diverse pathways lead to COX-2 or IL-8 expression. Depending on the original extracellular signals and the cell type involved, a variety of distinct signaling molecules, including MAPKs, NF-κB, PKA and PKC, have been revealed to involve in COX-2 and IL-8 expression.9,16,35 We used a variety of inhibitors to block each of these pathways and found that MAPKs, as well as NF-κB, but not PKA or PKC (data not shown), are the possible pathways acting downstream of IL-1β to mediate COX-2 and IL-8 production in human granulosa cells (Figures 3 and 4). We also clarified that the intracellular signaling pathways mediating IL-1β-elicited COX-2 expression appears to depend on two of the MAPKs, p38 and JNK, as well as on NF-κB, while the combined IL-1β and NS-398 promotion of COX-2 expression seems to depend on all of the MAPKs, as well as the NF-κB pathways (Figure 3). Interestingly, IL-1β-mediated IL-8 expression/secretion seemed to depend on p38 and JNK, but not ERK or NF-κB (Figure 4); the combined IL-1β and NS-398 promotion of cellular IL-8 expression would seem to be dependent on all the MAPKs and NF-κB, but IL-8 secretion is only dependent on p38, JNK and NF-κB (Figure 4). Notably, the NF-κB inhibitor did not affect the IL-1β alone-mediated IL-8 expression and secretion, and this contrasted with the findings for combined NS-398 and IL-1β regulation of IL-8 expression and secretion (Figure 4); this suggests that NF-κB might be a target of NS-398. These findings also suggest that, in human granulosa cells, IL-1β-regulated or NS-398-augmented IL-1β-mediated COX-2 and IL-8 may involve different pathways. Regardless of this, we were able to define that the IL-1β-induced phosphorylation of p38 and JNK in granulosa cells was not affected by NS-398 (Figure 5). In addition, IL-1β did not appear to induce ERK activation (Figure 5), and this is in accordance with the finding that the ERK inhibitor did not affect IL-1β-induced COX-2 or IL-8 expression (Figures 3 and 4).
In this study, one unanswered question that remains is who exactly is the interacting player with NS-398 within cells. It has been shown previously that arachidonic acid, the substrate of COX-2, is involved in the regulation of COX-2 expression in vitro and in vivo.36,37 Fujishima et al. demonstrated that the cytosolic phospholipase A2 is critical for cellular production of arachidonic acid and the subsequent regulation of COX-2 expression. 36 In addition, another study further revealed that, in a paracrine manner, arachidonic acid produced from the endometrial epithelial cells is able to regulate COX-2 expression in stromal cells and that PG synthesis is not involved in this induction. 38 In our study, as NS-398 blocks the bioactivity of COX-2, there seems a possibility that it may also result in an accumulation of arachidonic acid, which consequently induces COX-2 expression. Where such a scenario would lead to in our system needs further investigation. The detailed mechanism of how NS-398 functions in our study is still not completely clear and, in fact, so far there is no directly interacting partner other than COX-2 that has been identified. However, we have revealed that IL-1β-induced nuclear NF-κB p65 translocation is dramatically enhanced by NS-398 (Figure 6). Similarly, it has been shown previously in colon cancer cells that NS-398 is able to increase the DNA binding activity of NF-κB molecules; however, it should be noted that no NF-κB nuclear translocation was noted in the colon cancer cells. For translocation to occur, a long treatment time was needed and translocation was not observed at 48 h after incubation, being only noted after 72 or 96 h after incubation. 23 In addition, a relatively high dose (50–75 µM) was used in this earlier study, compared with the dose of 5–25 µM used in our study. Interestingly, a previous study using human gingival fibroblasts demonstrated that NS-398 at 10 nM was able to inhibit IL-1β-induced PGE2 and IL-6 production by inhibiting the IL-1β-mediated phosphorylation of IκBα and nuclear p65 translocation. 39 Currently, it is not clear whether this discrepancy is a result of the difference between the cell types and/or due to the dose of NS-398 used. Thus, this inconsistency needs further investigation in order to solve this puzzle.
In the human ovary, the pro-inflammatory cytokine IL-1β has been well recognized as playing a vital role in mediating various ovarian functions such as oocyte maturation, cumulus expansion, and ovulation by an up-regulation of COX-2 and IL-8 expression.7,10,40,41 NSAIDs have been widely used to reduce pain and inflammation, and NS-398 has been demonstrated to block COX-2-mediated PGE2 production and thus inhibit ovulation.24,25,42 In the current study we found that NS-398, despite its ability to attenuate IL-1β-mediated PGE2 production, is able to further enhance IL-1β-induced COX-2 and IL-8 expression (Figure 1). When considering what the NS-398-promoted elevation in COX-2 and IL-8 production may lead to in terms of physiological impacts on cells, our findings showed that combined IL-1β and NS-398 treatment resulted in cell cycle progression (Figure 7), and this effect could be neutralized by the IL-8 Abs (Figure 7). Furthermore, the exogenous addition of IL-8 also caused cell cycle progression, as well as an increase in the cell proliferation index (Figure 8a, b). Consistently, two negative cell cycle regulators, p21waf-1/cip-1 and p27kip-1, were also noted to be down-regulated by the combined IL-1β and NS-398 treatment (Figure 8c). 43 These findings suggest a novel effect of NS-398 on cell proliferation in the presence of IL-1β that involves the amplification of IL-1β-induced IL-8 production. Similar to our findings on the correlation between p21/p27 and the G2/M phase (Figures 7 and 8), a novel NO-releasing derivative from oleanolic acid, NOAD, has been reported to produce G2/M arrest, as well as an up-regulation of p21 and p27 expression in a dose-dependent manner. 44 Linalool, a component from a Chinese medicine containing members of the Plantaginaceae, has been demonstrated to induce the expression of p21 and p27, to arrest the U937 cells at the G0/G1 phase and to arrest the HeLa cells at the G2/M phase. 45 Aniline, a toxic aromatic amine, has been shown to down-regulate p21 and p27 expression and to promote rat splenocytes to go through the G2/M transition. 46 Finally, the nucleosome assembly protein 1-like 1 has been reported to promote the proliferation of the induced pluripotent stem cells by acting on the G2/M transition, and this been shown to be mediated via the down-regulation of p21 and p27. 47 All the above, as a whole, strongly suggest that the G2/M phase is, indeed, affected by p21 and p27. In addition, previous studies have reported that COX-2 overexpression in the presence of platelet-derived growth factor activity is able to mediate cell cycle arrest by increasing the expression of regulator p21waf-1/cip-1 and p27kip-1 in a PGE2-independent manner.48,49 However, we found a decrease rather than an increase in p21 and p27 expression (Figure 8). In our study, whether the detected reduction in p21 and p27 expression is connected to the increased level of COX-2 was not clear and needs further investigation. In addition, NS-398 has been previously shown using ovarian cancer cells to exert an inhibitory effect on cell proliferation at concentrations ranging from 50 to 500 µM, which is much higher than the dose needed, namely 1 µM, for the inhibition of COX-2 activity and the PGE2 production. 50 At a concentration of 25 µM in our system, which is below the 50–500 µM range, NS-398 alone does not appear to affect cell cycle progression or the expression of p21 and p27 (Figures 7 and 8). This also supports the hypothesis that NS-398 needs IL-1β stimulation as a condition for it to augment the effects in granulosa cells on inflammation and cell proliferation/cell cycle progression.
In conclusion, our study demonstrates a novel action for NS-398 as a potential inflammation-enhancing molecule via the augmentation of cytokine IL-1β-induced inflammation in ovarian granulosa cells. These effects occur, at least in part, by targeting the IL-1β-induced activation of the NF-κB signaling pathway (Figure 9).
Outline of IL-1β regulation of COX-2 and IL-8 production, and the potential acting points of NS-398 in ovarian granulosa cells. IL-1β interacts with its cognate cell surface receptor to induce the intracellular signaling pathways, including the MAPKs (p38, JNK, ERK), as well as the NF-κB pathways, both of which would lead to transcription of COX-2 and IL-8, and consequently mediate the output of PGE2 and IL-8. NS-398 inhibits the bioactivity of COX-2 and thus blocks the PGE2 synthesis; NS-398 appears to augment the IL-1β-mediated NF-κB activation, and subsequently enhance the IL-β-induced COX-2 and IL-8 transcription, resulting in elevation of COX-2 and IL-8 protein levels and IL-8 secretion.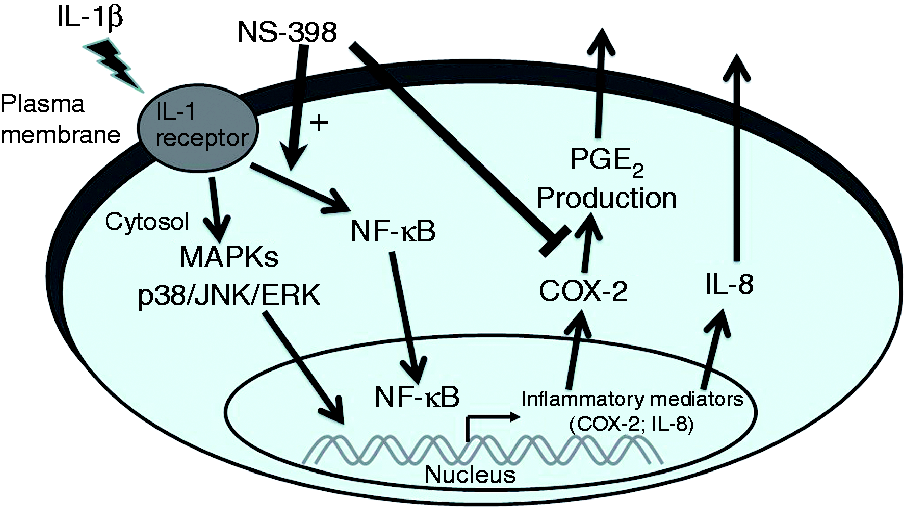
Footnotes
Acknowledgements
We thank Dr Ralph Kirby, Department of Life Sciences, National Yang-Ming University, for his help with language editing.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants from the Taiwan Ministry of Science and Technology (MOST 100-2320-B-010-018-MY3; MOST 102-2320-B-010-010-MY3), the Cheng Hsin General Hospital (103F003C03, 104F003C04, 105F003C04), and Taiwan Ministry of Education, Aim for the Top University Plan.