
Abstract
Human β-defensin3 (hBD3) and the cathelicidin LL-37 are host defense peptides (HDPs) that directly kill microbes and display immunomodulatory/wound-healing properties via the activation of chemokine, formylpeptide and epidermal growth factor receptors on leukocytes and epithelial cells. A C-terminal 14 amino acid hBD3 peptide with all Cys residues replaced with Ser (CHRG01) and an LL-37 peptide consisting of residues 17-29 (FK-13) display antimicrobial activity but lack immunomodulatory property. Surprisingly, we found that CHRG01 and FK-13 caused Ca2+ mobilization and degranulation in human mast cells via a novel G protein-coupled receptor known as Mas-related gene-X2 (MrgX2). At local sites of bacterial infection, the negatively charged LPS likely interacts with cationic HDPs to inhibit their activity and thus providing a mechanism for pathogens to escape host defense mechanisms. We found that LPS caused almost complete inhibition of hBD3 and LL-37-induced Ca2+ mobilization and mast cell degranulation. In contrast, it had no effect on CHRG01 and FK-13-induced mast cell responses. These findings suggest that HDP derivatives that kill microbes, harness mast cell’s host defense and wound-healing properties via the activation of MrgX2 but are resistant to inhibition by LPS could be utilized for the treatment of antibiotic-resistant microbial infections.
Introduction
Antibiotics have been used for the treatment of microbial infection since the early 1900s but the emergence of multidrug-resistant strains of microbes poses a tremendous public health concern globally. 1 Thus, there is an urgent need to develop novel therapy for the treatment of infectious diseases caused by antibiotic resistant organisms. Cationic antimicrobial peptides (AMPs), also known as host defense peptides (HDPs), such as the cathelicidin LL-37 and human β-defensins (hBDs), have the potential to be utilized as antimicrobial agents.2–4 In addition to their direct antimicrobial activity, LL-37 and hBDs display immunomodulatory and wound-healing properties via the activation of blood leukocytes, epithelial cells and endothelial cells.5–8 Most effects of hBD3 and LL-37 appear to be mediated via the activation of G protein-coupled formyl peptide receptor 2 (FPR2; also known as FPRL1) on monocytes, T cells and neutrophils. 7 These HDPs also activate chemokine receptor, CCR2 on macrophages, CCR6 on T cells, purinergic receptor P2X7 on fibroblasts and epidermal growth factor receptors in epithelial cells.5,9,10
Recent studies suggest that HDP’s immunomodulatory and wound-healing properties play greater roles in host defense against microbial infection than their antimicrobial activity.8,11,12 Not surprisingly, a number of laboratories have been engaged in identifying the structural components of hBD3 and LL-37 that promote antimicrobial and immunomodulatory properties. hBD3 is a 45-amino acid peptide that contains a three-stranded antiparallel β-sheet constrained by three intermolecular disulfide bonds. It has been shown that whereas Cys–Cys disulfide bridges in hBD3 are required for its chemotactic activity via CCR6, its bactericidal activity is unaffected in the absence of any disulfide bridge. 13 Furthermore, C-terminal 14-amino acid hBD3 peptides with all Cys residues replaced with Ser (CHRG01) display potent antimicrobial activity. 14 Li et al. 15 designed a series of LL-37 peptides to determine their structure/activity relationship. They showed that while an N-terminal fragment of LL-37 (sequence 1–12) has no antimicrobial activity, a peptide containing 13 amino acids within the central part of LL-37 (sequence 17-29; FK-13) is sufficient for killing microbes. Interestingly, peptides similar to FK-13 do not participate in the generation of chemokine from keratinocytes. 16 These findings suggest that distinct structural components of HDPs participate in antimicrobial and immunomodulatory activities.
Mast cells are multifunctional immune cells that are found in all vascularized tissues and are most common at sites that are exposed to the external environment, such as the skin, airway, oral mucosa and the intestine. Mast cells play a sentinel role in innate immunity and also contribute to wound healing, and these effects appear to be mediated, at least in part, by HDPs.17–22 Thus, murine mast cells express the human LL-37 counterpart, cathelicidin-related AMP (CRAMP), and its expression is up-regulated by LPS. 23 Furthermore, mast cells derived from CRAMP–/– mice display significant reduction in their ability to kill Gram-negative bacteria. 23 Exposure of human mast cells to pneumococci induces the release of LL-37 and a neutralizing Ab to LL-37 attenuates mast cell-dependent pneumococcal killing. 24 Enterococcus faecalis has emerged as an important cause of life-threatening multidrug-resistant bacterial infections in the hospital setting. Scheb-Wetzel et al. 25 recently showed that mast cells exert potent antimicrobial effect against this pathogen and that this effect is mediated via mast cell degranulation and the release of CRAMP. Furthermore, CRAMP has been shown to protect skin from necrotic skin infection and to promote healing. 26
HDPs activate a variety of signaling pathways in mast cells, including phospholipase C, the MAPKs (p38, ERK, JNK) for the induction of chemotaxis and mediator release.27–30 However, unlike the situation in other immune cells, the effects of HDPs on mast cells are not mediated via chemokine receptors, FPR2, P2X7 or epidermal growth factor receptors.31,32 We have recently shown that hBD3, LL-37 and other AMPs activate human mast cells via a novel G protein-coupled receptor, known as Mas-related gene-X2 (MrgX2).33–35 An important feature of MrgX2 that distinguishes it from other HDP receptors is that it is activated by a wide range of cationic amphipathic peptides.36–38 This raises the interesting possibility that hBD3 and LL-37-derived peptides such as CHRG01 and FK-13, which display antimicrobial activity, could activate mast cells via MrgX2.
In addition to immunomodulation and wound healing, HDPs display an anti-inflammatory effect via the inhibition of LPS-induced cytokine generation in monocytes and macrophages.39–41 It is therefore possible that negatively charged LPS interacts with cationic HDPs to inhibit their antimicrobial and immunomodulatory activities, thus providing a mechanism for Gram-negative bacteria to escape the host defense mechanisms. 42 The goals of the present study were to determine if CHRG01 and FK-13 activate mast cells via MrgX2 and to assess if LPS modulates mast cell activation by HDPs. The data presented herein demonstrate the novel finding that while HDPs and their peptide derivatives activate mast cells via MrgX2 their functions are modulated differently by LPS.
Materials and methods
Reagents
All cell culture reagents were purchased from Invitrogen (Gaithersburg, MD, USA). Native complement C3a was from Complement Technology (Tyler, TX, USA). DNP-BSA and DNP-specific mouse IgE (SPE-7) was purchased from Sigma-Aldrich (St. Louis, MO, USA). hBD3, LL-37, FK-13 and CHRG01 were purchased from Anaspec (Freemont, CA, USA). Escherichia coli LPS (serotype R515, LPS) was purchased from Alexis Biochemicals (San Diego, CA, USA) and Pseudomonas aeruginosa LPS (PSa-LPS) was obtained from Astarte Biologics (Bothell, WA, USA).
Culture of mast cell lines
Laboratory of Allergic Diseases 2 (LAD2) cells were maintained in complete StemPro-34 medium supplemented with 100 ng/ml human stem cell factor (hSCF).
43
Rat Basophilic Leukemia (RBL-2H3) cells were maintained as monolayer cultures in DMEM supplemented with 10% FBS,
Transfection of RBL-2H3 cells
RBL-2H3 cells were transfected with plasmids encoding hemagglutinin (HA)-tagged MrgX2 using the Amaxa nucleofector device and Amaxa kit V, according to the manufacturer’s protocol. Following transfection, cells were cultured in the presence of G-418 (1 mg/ml) and cells expressing equivalent receptors were sorted using an anti-HA specific Ab and FITC-conjugated anti-mouse IgG.
Calcium mobilization
Ca2+ mobilization was determined as described previously. 44 Briefly, cells (LAD2; 0.2 × 106, RBL-2H3 1 × 106) were loaded with indo-1 AM (1 µM) for 30 min at room temperature (25℃). Cells were washed and re-suspended in 1.5 ml of HEPES-buffered saline. Ca2+ mobilization was measured in a Hitachi F-2500 spectrophotometer with an excitation wavelength of 355 nm and an emission wavelength of 410 nm.
Degranulation
LAD2 cells (5 × 103) and RBL-2H3 cells (5 × 104) were seeded into 96-well plates in a total volume of 50 μl HEPES buffer containing 0.1% BSA and exposed to the indicated peptides. For total β-hexosaminidase release, unstimulated cells were lysed in 50 μl 0.1% Triton X-100. Aliquots (20 μl) of supernatant or cell lysate were incubated with 20 μl of 1 mM p-nitrophenyl-N-acetyl-β-
Results
CHRG01 and FK-13 activate mast cells via MrgX2
We have previously shown that hBD3 and LL-37 activate human mast cells via MrgX2.33,34 Comparison of amino acid sequences of hBD3, LL-37 and their smaller derivatives is shown in Figure 1. The C-terminal 14-amino acid peptide of hBD3 with the Cys residues replaced with Ser (CHRG01, net positive charge +8) display antimicrobial activity similar to hBD3 with LD90 (concentration of peptide at which 90% of bacteria are killed) values in the range of 1–6 µg/ml.
14
We therefore sought to determine if CHRG01 also activates mast cells using a human mast cell line (LAD2 cells) that endogenously expresses MrgX2.33,34,37 We found that CHRG01 caused a dose-dependent degranulation in LAD2 cells with an EC50 value of ∼10 µM (∼16 ng/ml) with a maximal release ∼40% of total granule content (Figure 2a). However, the potency and efficacy of CHRG01 for degranulation in LAD2 cells were much lower than those for hBD3 (Figure 2a). Nevertheless, the ability of CHRG01 to induce degranulation in LAD2 cells was associated with an increase in intracellular Ca2+ mobilization (Figure 2b, c).
Amino acid sequences of AMPs used in this study. Primary structures of hBD3, CHRG01, LL-37 and FK-13 with their net charges are shown. The positively charged residues in the peptides are underlined. hBD3, CHRG01, LL-37 and FK-13 cause degranulation and Ca2+ mobilization in LAD2 cells endogenously expressing MrgX2. (a, d) Cells were stimulated with different concentrations of peptides and percent degranulation (β-hexosaminidase release) was determined. Data are mean ± SEM of three experiments. Statistical significance was determined by two-way ANOVA with Bonferroni’s post test. * indicates P < 0.01 and ** indicates P < 0.001. (b, c, e, f) Cells were loaded with Indo-1AM and stimulated with CHRG01 [1 µM (b), 10 µM (c)] or FK-13 [1 µM (e), 10 µM (f)] and intracellular Ca2+ mobilization was determined. Data shown are representative of three similar experiments.
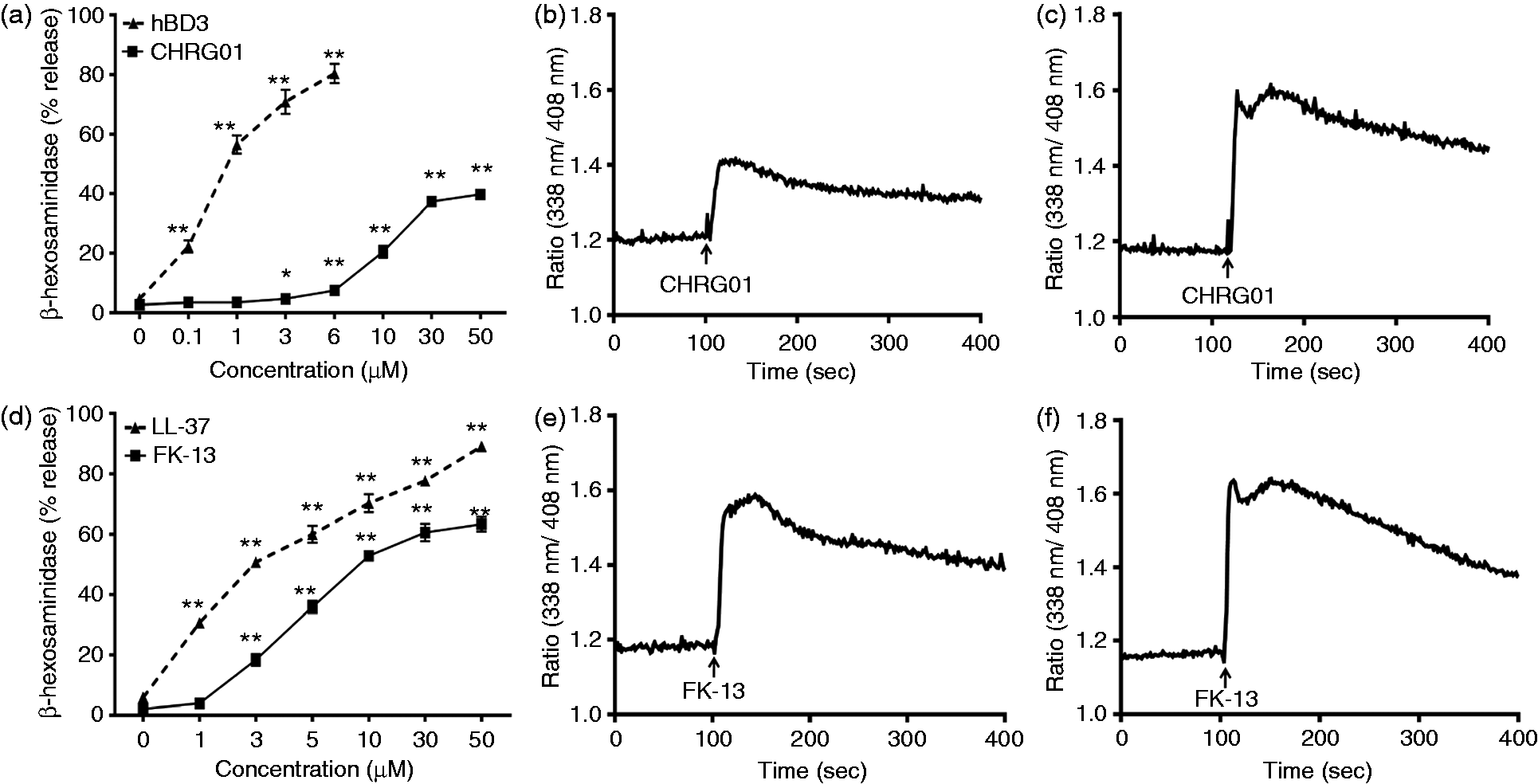
LL-37 has six net positive charges and activates human mast cells via MrgX2. 34 A 13-amino acid peptide corresponding to the residues 17-29 (FK-13; Figure 1) was identified as the central amphipathic helix that is critical for bacterial killing with an MIC (minimum inhibitory concentration for E. coli) value of 40 µM. 15 We found that FK-13 caused degranulation in LAD2 cells with an EC50 value of ∼5 µM and a maximum of 70% granule release (Figure 2d). However, unlike the situation with hBD3/CHRG01, the efficacy of FK-13 to induce degranulation in LAD2 cells was similar to that for LL-37. As for CHRG01, the ability of FK-13 to induce degranulation was associated with increased intracellular Ca2+ mobilization (Figure 2e, f).
A rodent mast cell line, RBL-2H3 cells, which do not express MrgX2, are unresponsive to human AMPs unless the cells are transfected with cDNA encoding the receptor.
38
To determine the role of MrgX2 on CHRG01 and FK-13-induced degranulation, we utilized RBL-2H3 cells stably expressing human MrgX2.33,34 In transfected RBL-2H3 cells, CHRG01 and FK-13 caused a dose-dependent increase in degranulation, with a similar profile as for LAD2 cells endogenously expressing MrgX2 (Figure 3a, d). Furthermore, as for LAD2 cells, the efficacy and potency of CHRG01 for degranulation in transfected RBL-2H3 cells were much lower than those for hBD3 (Figure 2a and Figure 3a). In addition, as for LAD2 cells, FK-13 and LL-37 induced degranulation in transfected RBL-2H3 cells with similar potency and efficacy (Figure 2d and Figure 3d). Given that CHRG01 was less potent than FK-13 for degranulation in both LAD2 cells and transfected RBL-2H3 cell, we expected a similar difference in the abilities of these peptides to induced Ca2+ mobilization in transfected RBL-2H3 cells. Indeed, we found that CHRG01 caused less robust Ca2+ mobilization than FK-13 when used at the same concentration (Figure 3b, e). We have previously shown that untransfected RBL-2H3 cells, which do not express MrgX2, are unresponsive to hBD3 and LL-37 for Ca2+ mobilization and degranulation.33,34 Not surprisingly, we found that CHRG01 and FK-13 did not induce intracellular Ca2+ mobilization in the absence of MrgX2 but the cells were fully responsive to antigen (Figure 3c, f). Taken together, these data demonstrate that CHRG01 and FK-13 activate mast cells via MrgX2.
hBD3, CHRG01, LL-37 and FK-13 cause degranulation and Ca2+ mobilization in RBL-2H3 cells ectopically expressing MrgX2. (a, d) Transfected RBL-2H3 cells were stimulated with different concentrations of peptides and percent degranulation (β-hexosaminidase release) was determined. Data are mean ± SEM of three experiments. Statistical significance was determined by two-way ANOVA with Bonferroni’s post test. *P < 0.01; **P < 0.001. (b, e) Transfected RBL-2H3 cells were loaded with Indo-1AM and stimulated with CHRG01 (3 µM) or FK-13 (3 µM) and intracellular Ca2+ mobilization was determined. (c, f) Un-transfected RBL-2H3 cells were exposed to IgE (16 h), washed and loaded with Indo-1AM. Cells were stimulated with CHRG01 (3 µM) or FK-13 (3 µM) followed by Ag (Ag, DNP-BSA, 30 ng/ml) and intracellular Ca2+ mobilization was determined. Data shown are representative of three similar experiments.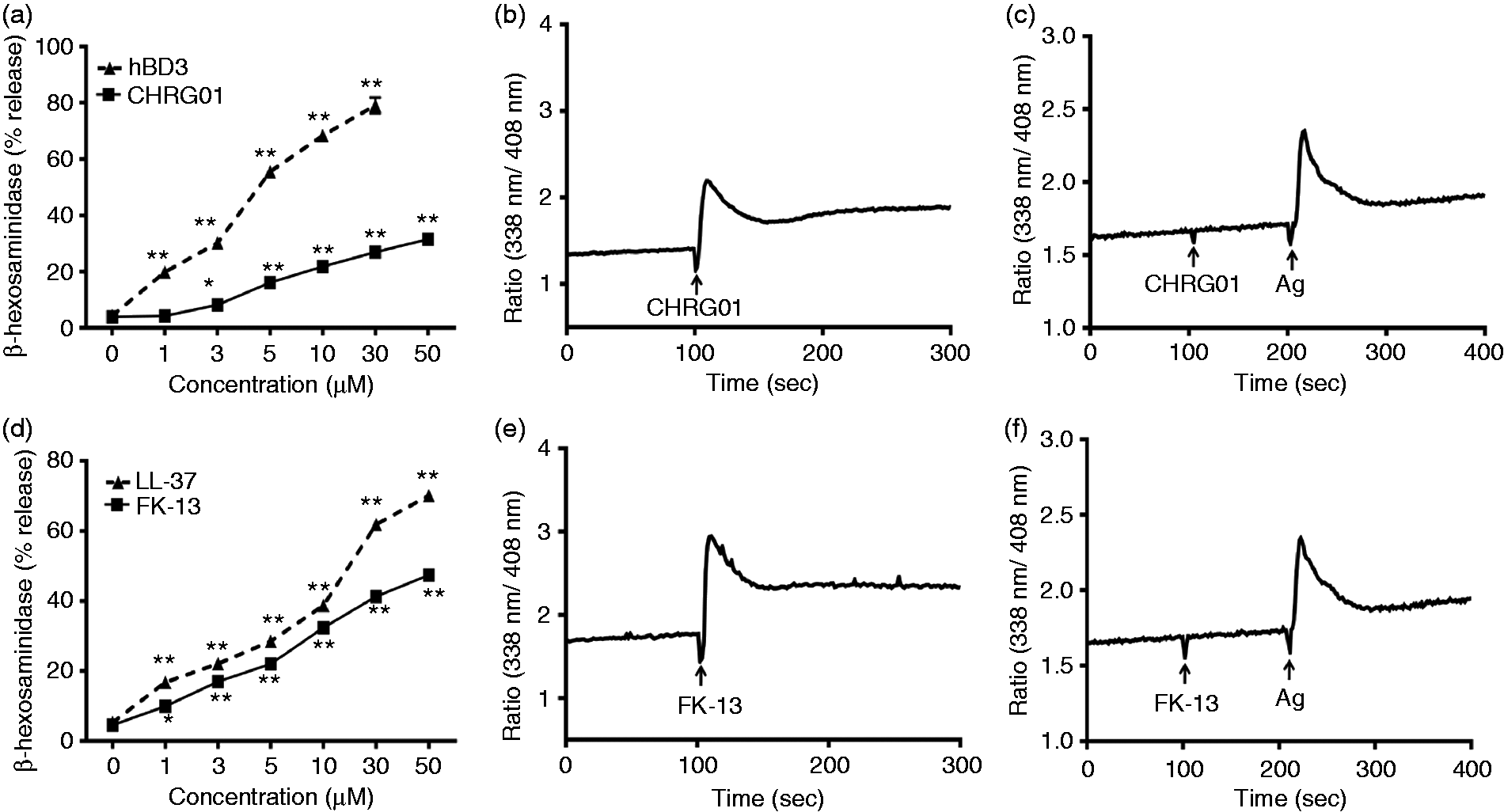
LPS inhibits hBD3/LL-37 but not CHRG01/FK-13-induced responses in mast cells
hBD3 has previously been shown to inhibit LPS-induced cytokine generation in macrophages.
39
Here, we sought to determine if LPS inhibits hBD3-induced Ca2+ mobilization and degranulation in mast cells. hBD3 (100 nM) caused a sustained intracellular Ca2+ response in LAD2 cells that did not return to basal level even after 5 min (Figure 4a). LPS (1 µg/ml) did not induce Ca2+ mobilization but pre-incubation of cells with LPS (1 µg/ml, 2 min) resulted in complete inhibition of hBD3-induced Ca2+ response (Figure 4b). While hBD3 activates human mast cells via MrgX2, the anaphylatoxin C3a does so via the activation of cell surface C3a receptor.33,45 To determine the specificity of LPS for hBD3 we tested its effect on C3a-induced Ca2+ mobilization. As shown in Figure 4a and 4b, unlike the situation for hBD3, LPS had no effect on the C3a-induced response. To test if pre-incubation of cells with LPS was required for inhibition of the hBD3 response we premixed LPS with hBD3 and determined Ca2+ mobilization. Under this condition, LPS inhibited the hBD3-induced Ca2+ response to the same extent as the 2 min pre-incubation protocol (Figure 4b, c). However, the ability of C3a to induce Ca2+ mobilization was not affected when the cells were exposed to the mixture of LPS and hBD3.
LPS inhibits hBD3-induced Ca2+ mobilization and degranulation in LAD2 cells. (a) Indo-1-loaded cells were sequentially exposed to hBD3 (100 nM) and C3a (10 nM) and intracellular Ca2+ mobilization was determined. (b) Cells were pre-treated with LPS (1 µg/ml) before exposure to hBD3 (100 nM) and C3a (10 nM) and intracellular Ca2+ mobilization was determined. (c) Cells were exposed to a mixture of LPS (1 µg/ml) and hBD3 (100 nM) followed by C3a (10 nM), as indicated, and Ca2+ mobilization was determined. Traces are representative of three independent experiments. (d) Cells were pre-treated with buffer (control) or LPS (1 and 10 µg/ml) and hBD3 (100 nM) or C3a (10 nM)-induced degranulation was determined. Data are mean ± SEM of three experiments. Statistical significance was determined by two-way ANOVA with Bonferroni's post test. **P < 0.001.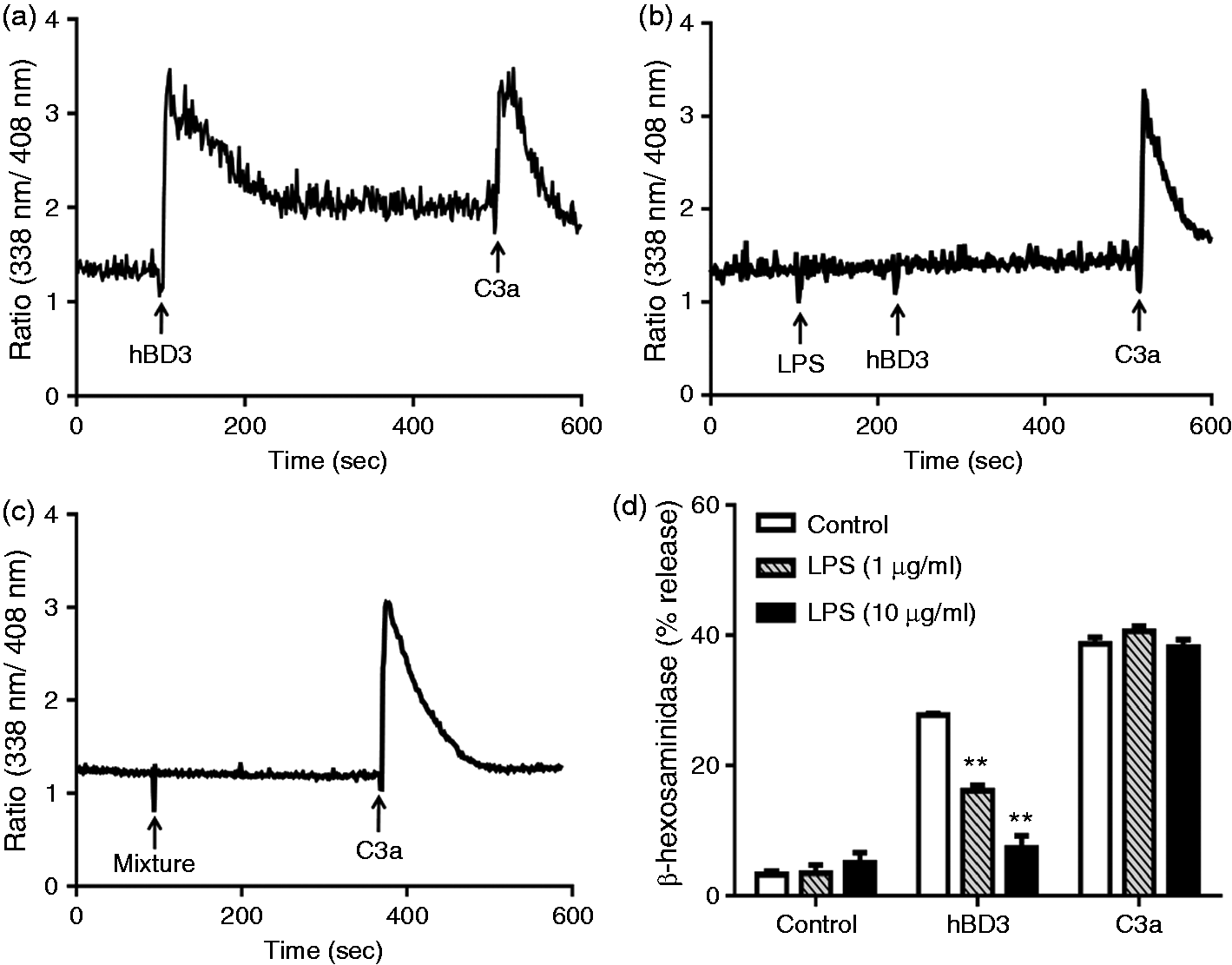
Intracellular Ca2+ mobilization is required for mast cell degranulation. 46 We therefore sought to determine the effects of LPS on hBD3-induced mast cell degranulation as measured by the release of β-hexosaminidase. Not surprisingly, LPS (1 µg/ml and 10 µg/ml) caused substantial inhibition of hBD3-induced degranulation (Figure 4d). Given that LPS had no effect on C3a-induced Ca2+ mobilization in LAD2 cells, we predicted that it would have no effect on C3a-induced degranulation. Indeed, as shown in Figure 4d, LPS did not inhibit the response to C3a.
As with hBD3, LL-37 has been shown to neutralize the ability of LPS to activate cytokine generation in macrophages.47–49 We therefore sought to determine if LPS inhibits LL-37-induced Ca2+ mobilization and degranulation in LAD2 cells. We found that LPS (2 min pre-incubation or added simultaneously) ablated LL-37-induced Ca2+ mobilization (Figure 5a–c) but had no effect on the response to C3a. Not surprisingly, LPS also blocked LL-37-induced degranulation in LAD2 cells (Figure 5d).
LPS inhibits LL-37-induced Ca2+ mobilization and degranulation in LAD2 cells. (a) Indo-1-loaded cells were sequentially exposed to LL-37 (1 µM) and C3a (10 nM) and intracellular Ca2+ mobilization was determined. (b) Cells were pre-treated with LPS (10 µg/ml) before exposure to LL-37 and C3a and intracellular Ca2+ mobilization was determined. (c) Cells were exposed to a mixture of LPS (10 µg/ml) and LL-37 (1 µM) followed by C3a, as indicated, and Ca2+ mobilization was determined. Traces are representative of three independent experiments. (d) Cells were pre-treated with buffer (control) or LPS (10 µg/ml) and LL-37 (1 µM)-induced degranulation (β-hexosaminidase release) was determined. Data are mean ± SEM of three experiments. Statistical significance was determined by two-way ANOVA with Bonferroni's post test. **P < 0.001.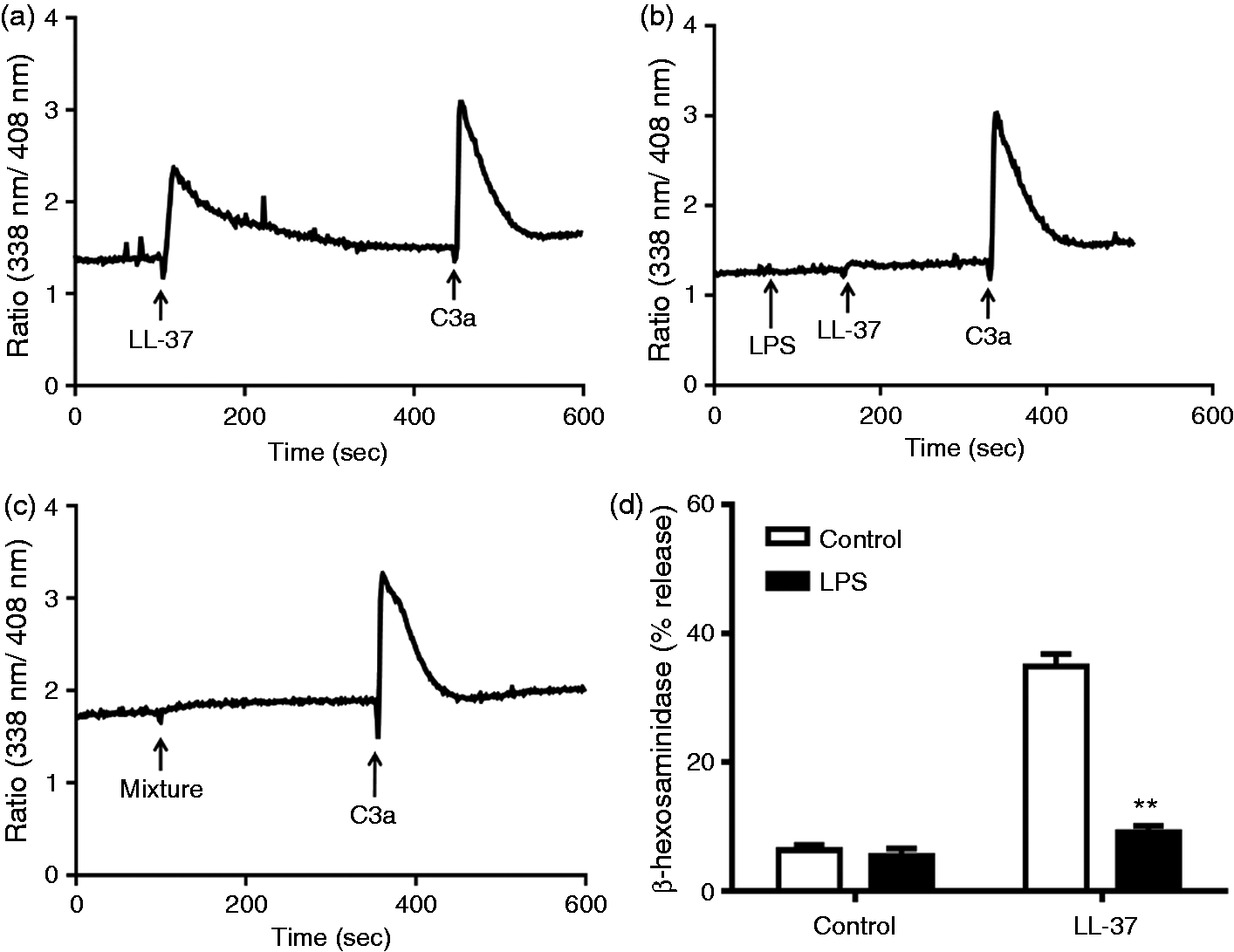
We next sought to determine if LPS inhibits CHRG01 and FK-13-induced mast cell degranulation. Unlike the situation with hBD3, LPS (10 µg/ml and 100 µg/ml) did not inhibit degranulation in response to CHRG01 (Figure 6a) or FK-13 in LAD2 cells endogenously expressing MrgX2 (Figure 6b). Finally, we tested the effects of LPS on LL-37, CHRG01 and FK-13-induced degranulation in RBL-2H3 cells ectopically expressing MrgX2. As shown in Figure 6c, LPS caused substantial inhibition of LL-37-induced degranulation but had no effect on CHRG01 or FK-13-induced response.
LPS has no effect on CHRG01 and FK-13-induced degranulation in LAD2 and RBL-2H3 cells. LAD2 cells were pre-treated with buffer (control) or LPS (10 and 100 µg/ml), and (a) CHRG01 and (b) FK-13-induced degranulation was determined. (c) RBL-2H3 cells stably expressing MrgX2 were pre-incubated with LPS (100 µg/ml) and the effects of LL-37 (10 µM), CHRG01 (10 µM) and FK-13 (10 µM)-induced mast cell degranulation was determined. Data are mean ± SEM of three experiments. Statistical significance was determined by two-way ANOVA with Bonferroni's post test. **P < 0.001.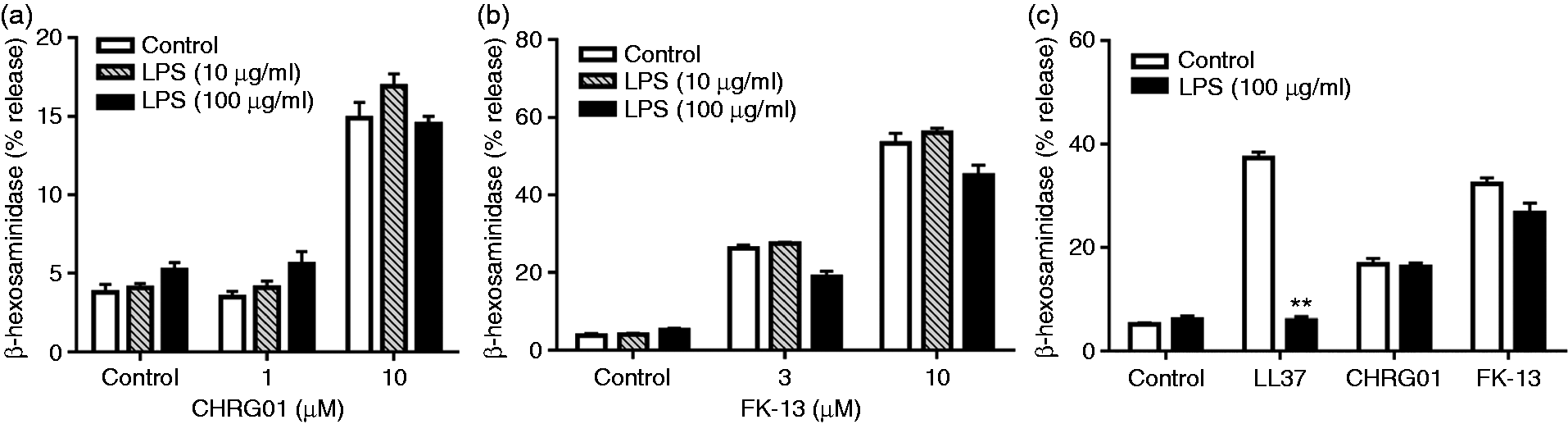
Effects of PSa-LPS on hBD3/LL-37 and CHRG01/FK-13-induced degranulation in mast cells
Most bacterial LPS contains three major components; the outermost polysaccharide, core oligosaccharide region and the innermost lipid A. The negatively charged component is the lipid A, which is the most active part of LPS and its structure differs greatly among Gram-negative bacterial species. Thus, the canonical E.Coli-LPS which possesses a hexa-acylated lipid A promotes a strong inflammatory response via the activation of TLR4.
50
In contrast, PSa-LPS is a heterogeneous mixture consisting of tetra- and penta-acylated lipid A and is a weak activator of TLR4.51,52 We therefore sought to determine if the acylation status of LPS affects its ability to modulate HDP-induced mast cell degranulation. We found that while E.Coli-LPS (LPS) caused almost complete inhibition of hBD3-induced (Figure 7a) and LL-37-induced (Figure 7b) degranulation in LAD2 cells, PSa-LPS inhibited this response by ∼50%. LPS or PSa-LPS had no effect on degranulation induced by CHRG01 (Figure 7a) or FK-13 (Figure 7b). These findings suggest that the ability of LPS to inhibit HDP-induced mast cell degranulation depends on the acylation status of the endotoxin and on the amino acid sequence of the peptide.
Effects of different LPS preparations on HDP-induced degranulation in LAD2 cells. (a) Cells were pre-incubated (5 min) with LPS 10 µg/ml, or PSa-LPS, 10 µg/ml, and stimulated with hBD3 (100 nM) and or with LPS 100 µg/ml, or PSa-LPS 100 µg/ml and stimulated with CHRG01 (10 µM), and percent degranulation (β-hexosaminidase release) was determined. (b) Cells were pre-incubated (5 min) with LPS 10 µg/ml, or PSa-LPS 10 µg/ml and stimulated with LL-37 (1 µM) or FK-13 (3 µM), and percent degranulation (β-hexosaminidase release) was determined. Data are mean ± SEM of three experiments. Statistical significance was determined by two-way ANOVA with Bonferroni's post test. *P < 0.01 and **P < 0.001.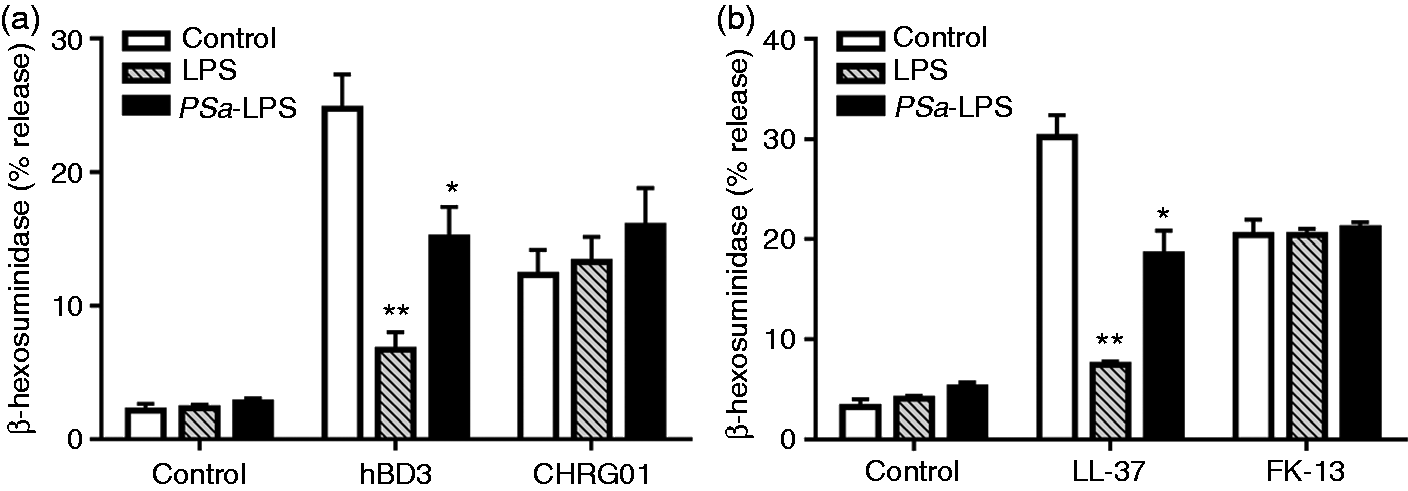
Discussion
hBD3 and LL-37 display antimicrobial activity against a wide spectrum of microorganisms and also have immunomodulatory properties. Interestingly, HDPs with little or no antimicrobial activity induce chemokines from epithelial cells, activate chemoattractant receptors on monocytes and protects mice from bacterial infection.53,54 These findings suggest that HDP’s immunomodulatory property plays a greater role in host defense against microbial infection than it antimicrobial activity. 12 Not surprisingly, attempts to identify the structural components of hBD3 and LL-37 that display antimicrobial and immunomodulatory properties have been the subject of many recent investigations.13–16,55 These studies showed that while short hBD3 and LL-37 peptides (CHRG01 and FK-13) display antimicrobial activity they do not activate chemokine generation in epithelial cells or promote monocyte chemotaxis. Surprisingly, data presented herein demonstrate the novel finding that CHRG01 and FK-13 activate mast cells via a novel G protein-coupled receptor, MrgX2, which is expressed only in human mast cells and no other immune or structural cells.36,56 Furthermore, we show that LPS inhibits HDP-induced mast cell degranulation and this inhibition depends on the acylation status of the endotoxin and on the amino acid sequence of the peptide.
CHRG01 and FK-13 were synthesized as the shortest peptides derived from hBD3 and LL-37 that have antimicrobial activity but do not display immunomodulatory properties.13–16 Given that HDPs activate a variety of receptors, including CCR2, CCR6, FPR2, P2X7 and EGFR, it was concluded that distinct regions of HDPs are important for their antimicrobial and immunomodulatory activities. We have recently shown that hBD3 and LL-37 induce mast cell degranulation via MrgX2.33,34 In the present study, we found that CHRG01 and FK-13 induced Ca2+ mobilization and degranulation in human mast cells endogenously expressing MrgX2, as well as RBL-2H3 cells ectopically expressing the receptor. These findings suggest that distinct structural components of HDPs are involved in monocyte/epithelial cell activation and mast cell degranulation via MrgX2. Given that CHRG01 and FK-13 also display antimicrobial activity, this suggests that shared structural domains of HDPs are involved in both microbial killing and mast cell activation via MrgX2.
The immunomodulatory properties of HDPs are thought to be mediated mostly via the activation of chemokine receptors on monocytes and lymphocytes. However, the significance of the finding that these peptides activate mast cells via MrgX2 is not clear. It is noteworthy that, unlike circulating leukocytes, mast cells are tissue-resident cells that are found beneath the epithelium and close to blood vessels. The epithelium expresses pattern recognition receptors (e.g. TLRs), which respond to infectious agents to generate HDPs.57,58 These cationic HDPs kill microbes by binding to negatively changed residues on the membrane and by forming pores. 59 Activation of mast cells by these peptides via MrgX2 cause increased vascular permeability and promote the recruitment of neutrophils, which contribute to host defense via their phagocytic activity.60,61 Furthermore, additional HDPs are released from neutrophils that likely contribute to host defense via the activation of mast cells through MrgX2.7,34 These findings suggest that MrgX2 expressed in mast cells provides an important link between the epithelium and neutrophils to promote host defense.
In addition to immunomodulation, HDPs display anti-inflammatory activity via the inhibition of LPS-induced cytokine generation in monocytes and macrophages.40,41,49 This effect is likely mediated via an electrostatic interaction between the cationic peptides and the negatively charged lipid A on endotoxin. This interaction prevents the binding of LPS to TLR4 on the surface of macrophages, thus inhibiting cytokine generation. 49 This contention is supported by the recent finding that citrullination of arginine residues within LL-37 results in a marked decreased in its affinity for LPS, leading to a substantially reduced ability of the peptide to inhibit cytokine generation in macrophages. 62 Another possible outcome of LPS/HDP interaction is that LPS inhibits HDP-induced mast cell degranulation via MrgX2, thus providing a mechanism for pathogens to escape mast cell-mediated host defense. 42 Indeed, we found that E.Coli-LPS caused almost complete inhibition of hBD3 and LL-37-induced Ca2+ mobilization and mast cell degranulation. However, PSa-LPS was less efficient in inhibiting hBD3 and LL-37-induced mast cell response. These findings suggest that Gram-negative bacteria that possess hexa-acylated lipid A species in their LPS (e.g. E. coli) are more likely to escape mast cell’s host defense strategy than those with tetra- or penta-acylated lipid A (e.g. PSa).
The most interesting finding of the present study was that while CHRG01 and FK-13 caused mast cell degranulation via MrgX2, this response was resistant to inhibition by LPS (E. coli or PSa). This suggests that CHRG01 and FK-13-based peptides could be developed for the treatment of antibiotic resistant bacterial infection because they would not only kill microbes but also harness mast cell’s host defense and wound healing properties without being inhibited by LPS. The mechanism via which LPS inhibits mast cell degranulation in response to hBD3/LL-37 without affecting the response to CHRG01/FK-13 is not known. However, this inhibitory effect is unlikely to be mediated at the level of the receptor because all four HDPs used in the present study activate mast cells via the same receptor, MrgX2. It is generally accepted that LPS binds to HDPs via an electrostatic interaction between the negative charges on LPS lipid A and positive charges on the peptide.49,62 Thus, it is possible that distinct regions of hBD3/LL-37 bind to LPS and MrgX2 and that CHRG01 and FK-13 possess the binding sites for Mrgx2 but not for LPS. CHRG01 is a 14-amino acid derivate of hBD3 corresponding to its C-terminal region. The finding that both hBD3 and CHRG01 induced mast cell degranulation via MrgX2 but LPS blocked the response to hBD3 but not to CHRG01 suggests that hBD3 utilizes the N-terminal region for interacting with LPS and its C-terminal region activating MrgX2. In contrast to CHRG01, FK-13 is a 13-amino acid peptide that corresponds to the central region of LL-37. We found that both LL-37 and FK-13 induced mast cell degranulation via MrgX2 but LPS inhibited mast cell response only to LL-37. This suggests that while the central region of LL-37 contributes to its mast cell activating property, the N- and C-terminal regions interacts with LPS. This contention is supported by the recent demonstration that neutralization of positive charges within the N and C terminal regions of LL-37 greatly reduces its affinity for interaction with LPS. 62
In conclusion, we have shown that hBD3 and LL-37-induced Ca2+ mobilization and mast cell degranulation via MrgX2 are potently inhibited by LPS. In contrast, the ability of CHRG01 or FK-13 to activate mast cells via the same receptor is resistant to inhibition by the endotoxin. Most studies evaluating clinical potentials of HDPs for antimicrobial activity involve their topical application at the site of infection. 63 At these sites, it is desirable for a therapeutic agent to display antimicrobial activity and to harness mast cells’ host defense and wound-healing properties. Given that LPS is likely to be present at the site of bacterial infection, it would also be important to design peptides that activate mast cells but are resistant to inhibition by the endotoxin. Thus, we propose that peptides based on CHRG01 and FK-13 could be developed as topical applications for the treatment of antibiotic-resistant cutaneous infections, without being inhibited by LPS. It is certainly possible that topical administration of these peptides could lead to excessive detrimental inflammation at the application site. However, given that mast cell-mediated inflammation is critical for host defense and wound healing, we believe that benefits of harnessing these mast cell-mediated responses in the context of cutaneous microbial infection outweighs the risks of potential detrimental outcome.
Footnotes
Acknowledgement
The authors thank Drs. Arnold Kirshenbaum and Dean Metcalfe (NIAID/NIH) for providing LAD2 mast cells.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by NIH grants AI108585 to HA and K99-HL121073 to HS.