
Abstract
Insects are capable of innate immune responses elicited after microbial infection. In this process, the receptor-mediated recognition of foreign bodies and the subsequent activation of immunocompetent cells lead to the synthesis ex novo of a peptide pool with antimicrobial activity. We investigated the inducible immune response of a coleopteran, Rhynchophorus ferrugineus, challenged with both Gram-negative and Gram-positive bacteria. After immunization, we evaluated the presence of antimicrobial peptides using either biochemical analyses or microbiological techniques. The antimicrobial properties of the newly synthesized protein pool, detectable in haemolymph fractions of low molecular mass, showed strong antibacterial activity against various bacterial strains (Escherichia coli, Pseudomonas sp. OX1, Bacillus subtilis and Micrococcus luteus). In addition to the preliminary study of the mechanism of action of the pool of antimicrobial peptides, we also investigated its effects on bacterial cell walls by means of fluorescence microscopy and scanning electron microscopy. The data suggest that the main effects seem to be directed at destabilizing and damaging the bacterial wall. This study provides data that help us to understand some aspects of the inducible innate immunity in a system model that lacks anticipatory responses. However, the weevil has finely tuned its defensive strategies to counteract effectively microbial infection.
Introduction
In recent years, the increase of bacterial infections exhibiting multiple drug resistance has been identified as a consequence of the widespread use of antibiotics. In many cases, treatments with conventional antibiotics have become less effective, resulting in dire implications for human and animal health. Thus, the need for alternatives to conventional antibiotics is greatly increasing, leading to increased attention on natural compounds with antimicrobial properties.1,2 Many authors have reported the discovery of new antimicrobial compounds from a broad range of organisms.3,4 Among these, insects, especially of the order Coleoptera, which comprises the largest order of eukaryotic organisms with highly skilled survival, seem to offer a good source for these types of molecules. 5
The remarkable fitness of insects is also owing to their powerful innate immune system, which shares factors structurally and functionally related to mammals.6,7 The immunity of insects is quite effective in protecting them from infections. They can recognize intruding foreign bodies and discriminate them from their own body structures by means of cellular and molecular mechanisms. In addition, their immune response seems to be variable and related to the specific pathogen invader.8–10
Insects recognize pathogen-associated molecular patterns (PAMPs) as a result of the interaction with endogenous pattern recognition receptors (PRRs). The interaction of PAMPs and PRRs elicits humoral and cellular defence, leading to the elimination of foreign pathogens.11,12 The presence of microorganisms and their PAMPs inside insects’ hemocoelic cavity triggers various short-term immune processes, such as prophenoloxidase-phenoloxidase (proPO) system activation (humoral encapsulation) or phagocytosis. However, these defence processes are only successful in the presence of a limited infection; when bacterial proliferation overcomes short-term defence, the onset of antibacterial responses, performed by antimicrobial peptides (AMPs) and lysozyme, is the most effective mechanism to prevent host septicaemia. 13 Evidence in the literature suggests that host AMPs could act in cooperation with other immunocompetent factors to modulate innate immune responses. 14 Insect AMPs are synthesized in fat body tissues (comparable with a mammalian liver) and released into the haemolymph during a systemic response against pathogens. The sensing of foreign bodies culminates in the synthesis ex novo of AMPs; thus, these are considered inducible factors. Their occurrence is consequent to the activation of immune genes mediated by evolutionary conserved Toll/Immune Deficiency (IMD) pathways present in both vertebrates and invertebrates.15–17 AMPs are strong cationic, heat stable and amphipathic molecules that have a variable Aa composition and length (30–60). Positively charged peptides interact directly with the anionic moiety of bacterial membranes (e.g. LPS, lipoteichoic and teichoic acids), inducing an increase of membrane permeability, leading to rapid cell death. 18
Rhynchophorus ferrugineus (Olivier) (Coleoptera, Curculionidae), also named Red Palm Weevil (RPW), is an emerging insect pest representing the most dangerous and deadly coleopteran affecting many palm trees; the knowledge of RPW immunity is of great interest as it lives in an environment of high bacterial proliferation. Furthermore, there have been several attempts to isolate pathogens from RPW, but none of them have been classified as potential biocontrol agents.19–21 The antimicrobial activity from the cuticular surface of adults and larvae, as well as from eggs and larvae haemolymph, was previously investigated; however, no antimicrobial peptides were identified. 19 Beyond any other reasons, the choice of this biological model is particularly favourable because of the late larval stage at which RPW reaches a considerable size, allowing researchers to work with a large haemolymph volume, and, if required, immunize and bleed larvae several times without sacrificing them. RPWs live and reproduce in warm habitats, and therefore are reared at temperatures (the inner temperature of infested palms is about 37℃) to which human pathogens are well adapted. Consequently, they represent a suitable host model to investigate pathogenesis and immune responses to potential harmful microorganisms.
We started a project to investigate the response of RPW to bacterial infections, with the aim of verifying and analysing the production of AMPs. We assayed the effects of RPW antimicrobial peptides against both Gram-negative and Gram-positive strains, derived from bacteria-challenged RPW larvae. Antimicrobial activity was evaluated in whole haemolymph and samples of low molecular mass against Escherichia coli, Pseudomonas sp. OX1, Bacillus subtilis and Micrococcus luteus. To detect newly synthesized molecules, we analysed partially purified host haemolymph patterns by electrophoretic methods. We also investigated, by means of fluorescence and scanning electron microscopy (SEM), the effects on bacterial cell walls induced by AMP treatments. Finally, we checked the effects of larvae haemolymph against human red blood cells (RBCs) to assess possible haemolytic activity.
Materials and methods
Reagents and Instruments
Instruments and reagents were purchased from BioRad Laboratories (Detroit, MI, USA), Sigma Chemicals (St. Louis, MO, USA), SERVA Electrophoresis (Heidelberg, GE), Millipore (Billerica, MA, USA), Merck (Darmstadt, Germany), Celbio SpA (Milan, Italy), General Electric Healthcare (Uppsala, Sweden) and Sartorius Stedim Biotech GmbH (Göttingen, Germany). Centrifugations were performed by an Eppendorf MiniSpin and Eppendorf 5804R (Eppendorf, Hamburg, Germany). Spectrophotometric measurements were carried out using a Jasco V-560 (Jasco, Easton, MD, USA). Bright field and fluorescence microscopy were performed with an Olympus BX81 inverted microscope equipped with UPlanFl 40 × and a UPLFLN 100 × objectives. Images were acquired by a B/W CCD camera (PCO Sensicam) and PCO Camware software (PCO Kelheim, Germany). Images were analysed by ImageJ software (NIH, Bethesda, MD, USA). All materials and buffers were autoclaved or filtered on 0.22 µm. Assays were performed at 4℃ under sterile conditions when required.
Biological models
Rhynchophorus ferrugineus (Olivier) (Coleoptera, Curculionidae) larvae were captured in June 2013 after a cut of infested palms (Phoenyx canariensis Hort. ex Chabaud) in the area of Ribera (AG, Sicily, Italy) and moved (authorization MIPAF protocol 0025254) to our laboratory for experimental tests. RPW at various stages were maintained in a climatic chamber at 28℃, in the dark, with a relative humidity of 75%. Insects were fed with a formula based on apple slices, and healthy larvae (about 6–7 g) were used in all assays. All the assays were repeated at least five times, using about 200 larvae.
Bacterial strains and culture conditions
Gram-negative (E. coli C1a, Pseudomonas sp. OX1) and Gram-positive (B. subtilis ATCC 6051, M. luteus ATCC 4698) bacterial cultures were used for larval immunizations and AMP activity assays. After inoculation in Luria-Bertani (LB) broth (1% tryptone, 0.5% yeast extract, 0.5% NaCl), bacterial cultures were grown overnight (16 h) under shaking at 37℃, in a dark room. Bacterial growth was verified by spectrophotometric measurement of biomass (λ = 600 nm). Briefly, for immunization assays, cultures were centrifuged at 1700 g for 10 min at 20℃, the bacterial pellet was recovered and then cells were killed by heating at 65℃ for 1 h. Before injections, bacteria were washed several times with sterile PBS (138 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4/KH2PO4, pH 7.4); finally, bacterial strains (a mixture 1:1 of E. coli and B. subtilis) were injected in RPW larvae at a final concentration of 105 CFU/ml, or stored at –20℃ with glycerol (1:1 v/v).
AMP synthesis in RPW larvae
In order to induce the synthesis of AMPs in R. ferrugineus, larvae were infected with killed or live microorganisms, or a mix of purified LPS and peptidoglycan (PGN). Fifty microlitres of bacterial suspension (105 CFU/ml dead bacteria or 103 CFU/ml live bacteria), or 20 µg of LPS/PGN mixture, were injected using Hamilton microsyringes (model gas-tight) equipped with thin needles (0.13 mm internal diameter, 0.26 mm outer diameter); after 24 h of infection, haemolymph was collected. Immunized and naïve larvae were anaesthetized on ice, sterilized with 70% ethanol and bled by puncturing the dorsal vessel by a sterile needle. Haemolymph was flushed out in a refrigerated sterile microfuge tube containing a few 1-phenyl-2-thiourea crystals to avoid activation of prophenoloxidase enzyme. Humoral fractions were collected by increasing centrifugation (200, 400 and 1500 g for 10 min at 4℃) to remove cells and tissue debris. Whole haemolymph was processed to obtain low molecular mass (LMM) fractions (cut-off 30 kDa and 10 kDa) by means of centrifugal filter devices after centrifugation at 1500 g for 4 h at 15℃. Total proteins were estimated by Bradford protein assay, calibrated on BSA. All samples were used immediately or stored at –20℃.
Antimicrobial activity in haemolymph of R. ferrugineus larvae
To evaluate antimicrobial activity in the humoral fraction of RPW haemolymph, bacterial cells were grown overnight (as described above) and then diluted with LB broth to 106 CFU/ml for microbroth dilution assays, carried out in 96-microwell plates. Aliquots (20 µl) of whole haemolymph or fractioned haemolymph (<30 kDa and 10 kDa), with different amounts of AMP, were added to 180 µl of bacterial suspension and then incubated for 3 h at 37℃ under shaking. After incubation, 100 µl of each sample was placed in a well of a 96-well microtiter plate, samples were serially diluted with phosphate buffer (61.4 mM K2HPO4, 38.4 mM H2PO4) and finally, they were plated on solid agar and bacteria colonies counted after incubation at 37℃ for 24 h. The antibacterial activity in haemolymph samples was intended as the percentage of bacterial survival compared with the control (bacterial suspension incubated without RPW haemolymph). The final concentration of proteins used for antimicrobial activity tests were as follows: versus E. coli, whole haemolymph 500 µg/ml, <30 kDa and <10 kDa fractions, 15, 75, 150 µg/ml; versus Pseudomonas, <30 kDa and <10 kDa fractions, 75 and 150 µg/ml; versus B. subtilis, <30 kDa fraction, 15, 75, 150 µg/ml and <10 kDa fraction, 75 and 150 µg/ml; versus M. luteus, <30 kDa fraction, 75 and 150 µg/ml, <10 kDa fraction, 75 and 150 µg/ml.
Effects of lysozyme and lysozyme inhibition on antimicrobial activity versus B. subtilis
Haemolymph fractions from RPW larvae were assayed against B. subtilis in the presence of lysozyme or lysozyme inhibitor [L-histidine methyl ester dihydrochloride (LHMED)]. Haemolymph samples, 150 µg/ml of either <30-kDa or <10-kDa fractions, were incubated with 106 CFU/ml of B. subtilis in the presence of 100 mM LHMED or lysozyme (2 µg/µl), respectively, for 3 h at 37℃, under shaking in a dark room. Finally, antimicrobial activity was evaluated by microbroth dilution assay, as described above.
SDS- and 2D-PAGE analysis of larvae haemolymph
Whole (cell-free) and fractioned haemolymph (<30 kDa and <10 kDa) from non-immunized and immunized larvae were analysed by monodimensional SDS-PAGE, 22 16% Tricine-PAGE 23 and 2D electrophoresis. 24 2D-page was performed using an Ettan IPGphor II Isoelectric focusing System unit (GE Healthcare), according to the standard manufacturer’s protocols. Samples were focused on an immobiline Drystrip (13 cm; pH 3–10), Isoelectric focusing (IEF) strips were loaded onto a Protean II Cell System (Bio-Rad, Hercules, CA, USA), proteins were separated by Tricine-PAGE (16%) and revealed by silver staining. Molecular mass determination was carried out following the method of Weber and Osborn. 25
RBC lysis in the presence of RPW haemolymph
The lysis of human RBCs was checked by recording haemoglobin release into the extracellular environment after the incubation of blood cells with whole and fractioned haemolymph. Approximately 10 ml whole blood was drawn into K3 EDTA vacuum tubes (BD Vacutainer, Becton Dickinson Inc., USA), from healthy volunteers. Blood testing solution was prepared by washing 3 ml fresh human blood with 7 ml of pyrogen-free PBS (137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, 2 mM KH2PO4, pH 7.4); the suspension was centrifuged at 400 g for 10 min at room temperature (20℃), and washes were repeated until the supernatant turned clear. RBC pellets were diluted to 20 ml with PBS, and 180 µl of diluted blood was added to 20 µl of whole haemolymph from naïve larvae, <30 kDa haemolymph fractions from naïve larvae, and either <30 or <10 kDa haemolymph fractions from immunized larvae. All the samples were incubated for 30 min at 37℃ under gentle shaking. Positive or negative controls were performed by adding to blood samples 20 µl of 0.2% Triton X-100 or PBS respectively. Samples were centrifuged at 400 g for 5 min and 100 µl of supernatants were diluted to 1 ml with PBS. Finally, cells lysis was assessed by measuring the OD (λ = 404 nm) with a JASCO V-530 UV/VIS spectrophotometer. The percentage of haemolysis was calculated as follows: [(Apeptide − APBS)/(ATriton-X100 − APBS)] × 100.
Fluorescence microscopy of propidium iodide uptake
Bacteria from overnight cultures were diluted 1:10 (v/v) with LB and cultured until mid-log phase OD 0.6 (λ = 600 nm). Then, AMP samples (haemolymph fractions <30 kDa and <10 kDa) were added to cell cultures to a final concentration of 150 µg/ml. All assays were performed with 2.5 × 107 CFU/ml of E. coli and B. subtilis, incubated at 37℃ in a thermomixer, shaken at 800 rpm.
Microscope observations were carried on bacteria after various incubation times (up to 3 h). Propidium iodide (PI; 5 µg/ml) was added at 20℃ and samples drops (10 µl) were placed in glass bottom (0.170-mm-thick round glass cover slip) observation chambers (Okolab, Ottaviano, Italy) and then covered with a small round glass cover slip in order to trap cells, and reduce their mobility and medium evaporation.
SEM
AMPs treated or not treated with aliquots of E. coli or B. subtilis suspensions (106 CFU/ml) were fixed with 2.5 % glutaraldehyde in 100 mM PBS (pH 7.4) overnight at 20℃, and then post-fixed in 1% osmium tetroxide, dehydrated with graded ethanol, dried by the critical point method and coated with gold. Observations were made under a SEM LEO-1430 (Carl Zeiss, GmbH) scanning electron microscope.
Data processing and statistical analysis
Statistical analyses were performed using the Fisher’s exact test. Means and SD were calculated in all assays; a P-Value <0.05 was considered statistically significant and evaluated by comparison with naïve samples. All experiments were replicated at least five times. Data were processed with GraphPad Prism 4 (GraphPad Software, La Jolla, CA, USA) and R version 3.0.2 (Free Software Foundation, Boston, MA, USA).
Results
Protein and peptide analysis in RPW haemolymph
Protein and peptide pools in the haemolymph of RPW larvae were studied by means of 1D and 2D analytical electrophoresis.
We assessed the effects of the infection by microorganisms by examining the protein pool before and after treatments (samples from non-immunized and immunized larvae). The expected occurrence of newly synthesized peptides was verified by electrophoretical methods.
In Figure 1, left panel (SDS-PAGE), patterns of samples from naïve larvae, larvae immunized with a LPS/PGN mixture of PAMPs and larvae immunized with Gram-positive/Gram-negative bacteria mix are depicted. Differences in patterns are evident even if the SDS-PAGE does not provide a clear resolution of LMM compounds. Variations in protein patterns are particularly evident in the LMM region (<30 kDa) as we analysed fractioned haemolymph samples by means of Tricine-PAGE (Figure 1, right panel). The right-hand panels in Figure 1 (A and B) show electrophoretic patterns of haemolymph samples <30 kDa and <10 kDa, respectively. New bands, absent in the control, are particularly evident in the region between 5 kDa and 20 kDa.
Monodimensional PAGE analysis of haemolymph samples from naïve and immunized RPW larvae. Whole and fractioned (<30 and <10 kDa) haemolymph from naïve, PAMP-infected and bacteria-infected larvae, were analysed by SDS-PAGE and Tricine-PAGE. Left: 10% SDS-PAGE of whole haemolymph. S: standard molecular mass; n: whole haemolymph from naïve larvae; iP: whole haemolymph from PAMP-injected larvae; iB+/–: whole haemolymph from mixed (Gram-positive plus Gram-negative) bacteria-immunized larvae. Right: 16% Tricine PAGE of fractioned haemolymph. (A) S: standard low molecular mass; n: <30 kDa haemolymph from naïve larvae; iB+: <30 kDa haemolymph from Gram-positive-immunized larvae; iB–: <30 kDa haemolymph from Gram-negative-immunized larvae; iB+/–: <30 kDa haemolymph from Gram-positive plus Gram-negative-immunized larvae. (B) n: <10 kDa haemolymph from naïve larvae; iB+/–: <10 kDa haemolymph from Gram-positive plus Gram-negative-immunized larvae.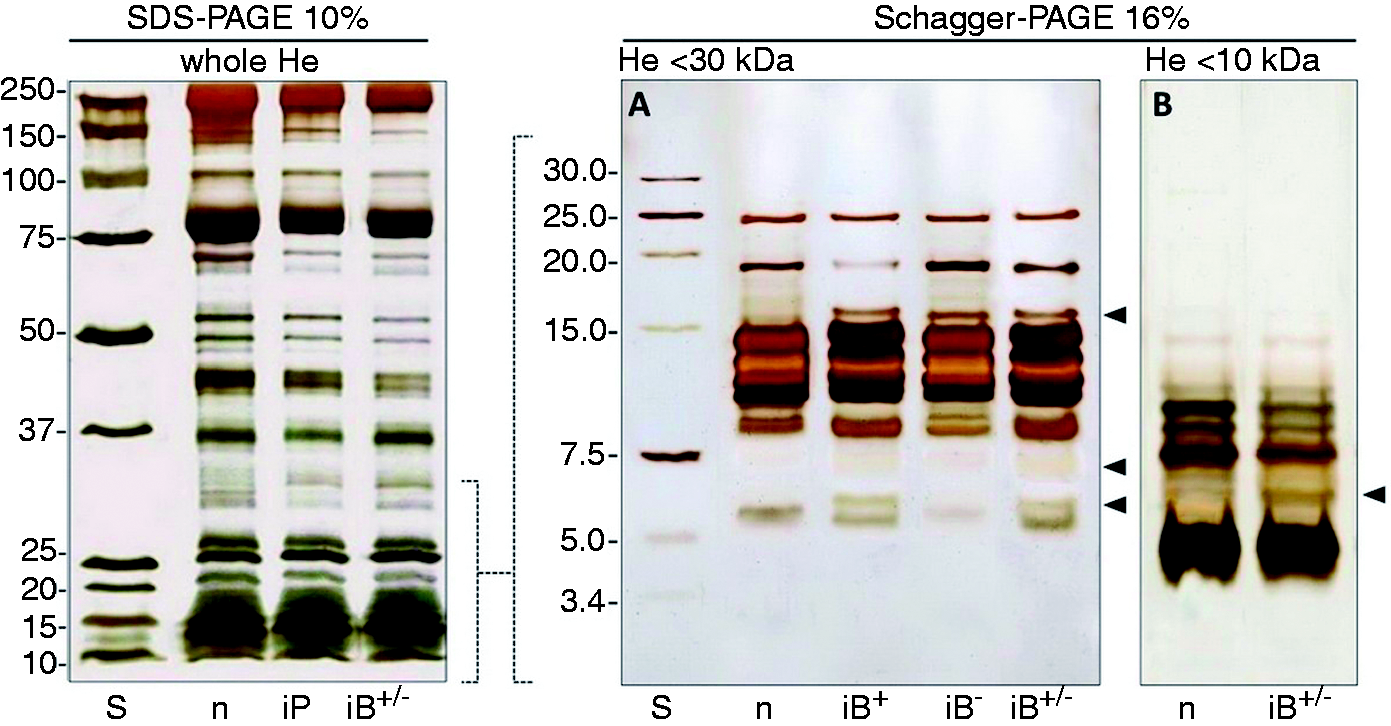
To further investigate the occurrence of new peptides (putative AMPs) we analysed the 2D-PAGE proteomes of <30 kDa haemolymph fractions (Figure 2). The first dimensions (isoelectric focusing gels) were carried out on linear 3–10 pH gradient gels, followed by a second dimension on Tricine-PAGE (16%). Two-dimensional analysis patterns of haemolymph samples are observable in Figure 2.
Bi-dimensional PAGE analysis. Two dimensional-PAGE analysis of proteins and peptides in fractions <30 kDa of haemolymph from (A) naïve and (B) bacteria-immunized larvae. IEF first dimension was run on a 3–10 linear pH gradient and the second dimension was carried out on 16% Tricine-PAGE. More than 70 spots are counted, and dotted boxes refer to areas in which new or up-regulated spots were observed (B). Boxed areas are enlarged below the panels (A1 and B1); circled spots (panel B1) indicate newly appeared or up-regulated spots.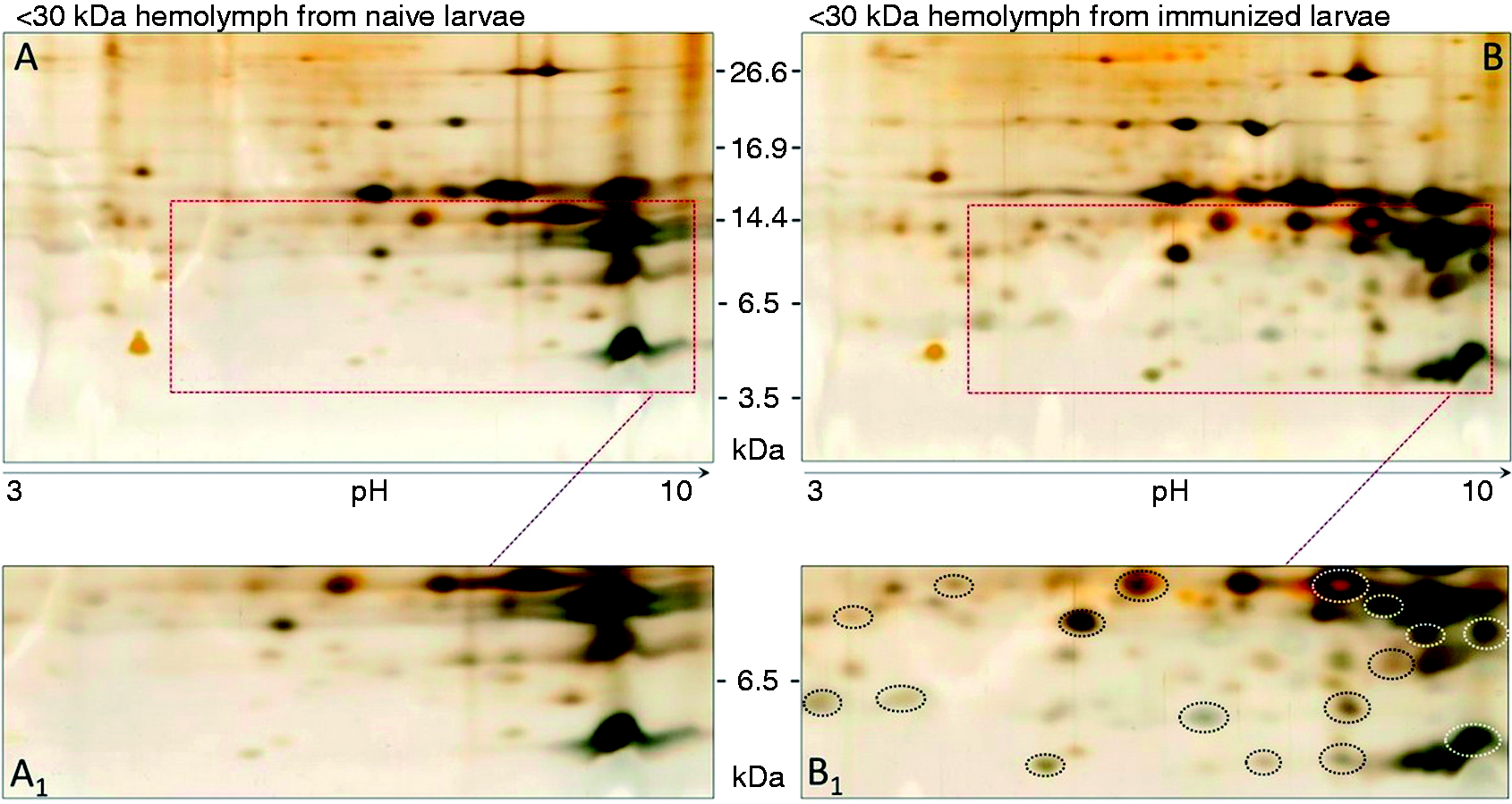
Antimicrobial activity assays versus Gram-negative bacteria
To verify the presence of antimicrobial activity (AMPs) in RPW haemolymph, we carried out bacteria growth tests on solid agar after serial dilution of bacteria incubated with AMPs. Samples used as source of AMPs were either the whole haemolymph (cell-free) or LMM fractions. Figure 3A shows the effects of whole haemolymph from naïve larvae (Hewn), live and dead bacteria-infected larvae (Hei-L and Hei-D) and PAMP-immunized (Hei-LPS/PGN) larvae against Gram-negative bacteria (E. coli). In all assays with immunized larvae (Hei-L and Hei-D and Hei-LPS/PGN), drastic antimicrobial effects are evident (bacteria mortality exceeds 99.9%) compared with the control growth (Ct3). Moreover, in naïve larvae haemolymph (Hewn), low activity is present. The high mortality rate was confirmed when antibacterial activity was tested in fractioned haemolymph samples. Figure 3 (B, C) shows data obtained with <30 kDa and <10 kDa haemolymph, respectively. In both cases, the activity of AMP increased with total protein concentration (Hei-15 < Hei-75 < Hei-150). The antimicrobial activity of the <30 and <10 kDa haemolymph fractions was also assayed on Pseudomonas sp., which seems to be highly susceptible to the action of RPW peptides.
Effects of AMPs versus Gram-negative bacteria. Haemolymph (He) from RPW larvae, not fractioned and fractioned were assayed against Gram-negative bacteria (E. coli). (A) Whole He (500 µg/ml proteins) was incubated with 106 CFU/ml E. coli; after 3 h, bacterial survival was evaluated by CFU count. Ct3: control (bacteria); Hewn: bacteria + He cell-free fraction (CFF) from naïve larvae; Hei-L: bacteria + He from larvae immunized with live bacteria; Hei-D: bacteria + He from larvae immunized with dead bacteria; Hei-LPS/PGN: bacteria + He from larvae immunized with bacterial PAMPs. (B, C) He fractions (<30 kDa and <10 kDa) from RPW larvae were assayed against Gram-negative bacteria (E. coli). (B) Fractioned CFF samples (<30 kDa) were incubated with 106 CFU/ml E. coli; after 3 h, bacterial survival was evaluated by CFU count. Ct3: control (bacteria); Hen: bacteria + He fraction (<30 kDa; 150 µg/ml) from naïve larvae; Hei-15, Hei-75, Hei-150: bacteria + He fraction (<30 kDa, 15, 75, 150 µg/ml, respectively) from immunized larvae. (C) Ct3: control (bacteria); Hen: bacteria + He fraction (<10 kDa; 150 µg/ml) from naïve larvae; Hei-15, Hei-75, Hei-150: bacteria + He fraction (<10 kDa, 15, 75, 150 µg/ml, respectively) from immunized larvae. Also, He from RPW larvae, not fractioned and fractioned were assayed against Gram-negative Pseudomonas sp. (D) Ct3: control (bacteria); Hen: bacteria + He fraction (<30 kDa; 150 µg/ml) from naïve larvae; Hei-75 and Hei-150: bacteria + He fraction (<30 kDa; 75 and 150 µg/ml, respectively) from immunized larvae. (E) Assays as in (D) carried out with <10-kDa fractions. Results are expressed as percentage of survival compared with the control Ct3 (100% survival). *P < 0.05; ***P < 0.001.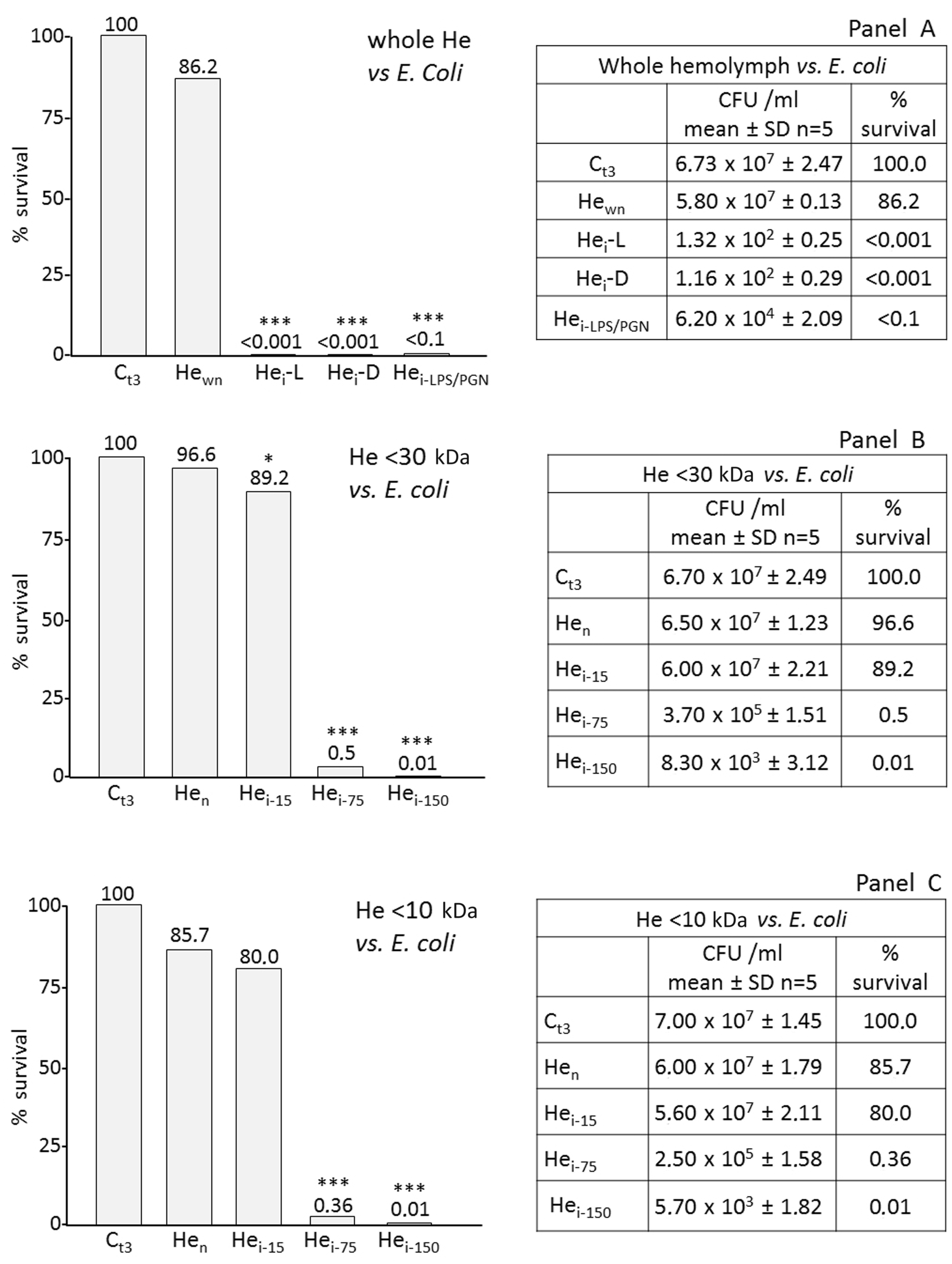
Antimicrobial activity assays versus Gram-positive bacteria
The effects against Gram-positive bacteria were assessed with 30 kDa and 10 kDa haemolymph fractions versus B. subtilis (Figure 4A, B) and versus M. luteus (Figure 4C, D). Fractions <30 kDa possess a dose-dependent antibacterial activity either versus B. subtilis (Figure 4A) or versus M. luteus (Figure 4C). In addition, a rate of bacteria mortality was also observed in the sample from naïve larvae, particularly evident against M. luteus (Figure 4C, Hen). Assays performed with <10 kDa fractions versus B. subtilis and M. luteus, revealed a reduced antibacterial activity either in samples from immunized larvae or in samples from naïve larvae (Figure 4B, D).
Effects of AMPs versus Gram-positive bacteria. Haemolymph (He) fractions [(A) <30 kDa and (B) <10 kDa] from RPW larvae were assayed against Gram-positive bacteria. Fractioned cell-free fraction (CFF) samples were incubated with 106 CFU/ml B. subtilis; after 3 h, bacterial survival was evaluated by CFU count. (A) Ct3: control (bacteria); Hen: bacteria + He fraction (<30 kDa; 150 µg/ml) from naïve larvae; Hei-15, Hei-75, Hei-150: bacteria + He fraction (<30 kDa, 15, 75, 150 µg/ml, respectively) from immunized larvae. (B) Ct3: control (bacteria); Hen: bacteria + He fraction (<10 kDa; 150 µg/ml) from naïve larvae; Hei-75, Hei-150: bacteria + He fraction (<10 kDa, 75 and 150 µg/ml, respectively) from immunized larvae. Also, He fractions from RPW larvae were assayed against M. luteus. (C) He fraction <30 kDa; Ct3: control (bacteria); Hen: bacteria + He fraction (150 µg/ml) from naïve larvae; Hei-75 and Hei-150: bacteria + He fraction from immunized larvae (75 and 150 µg/ml, respectively). (D) He fraction <10 kDa; Ct3, control (bacteria); Hen: bacteria + He fraction (150 µg/ml) from naïve larvae; Hei-75 and Hei-150: bacteria + He fraction from immunized larvae (75 and 150 µg/ml, respectively). Results are expressed as percentage of survival compared with the control Ct3 (100% survival). ***P < 0.001.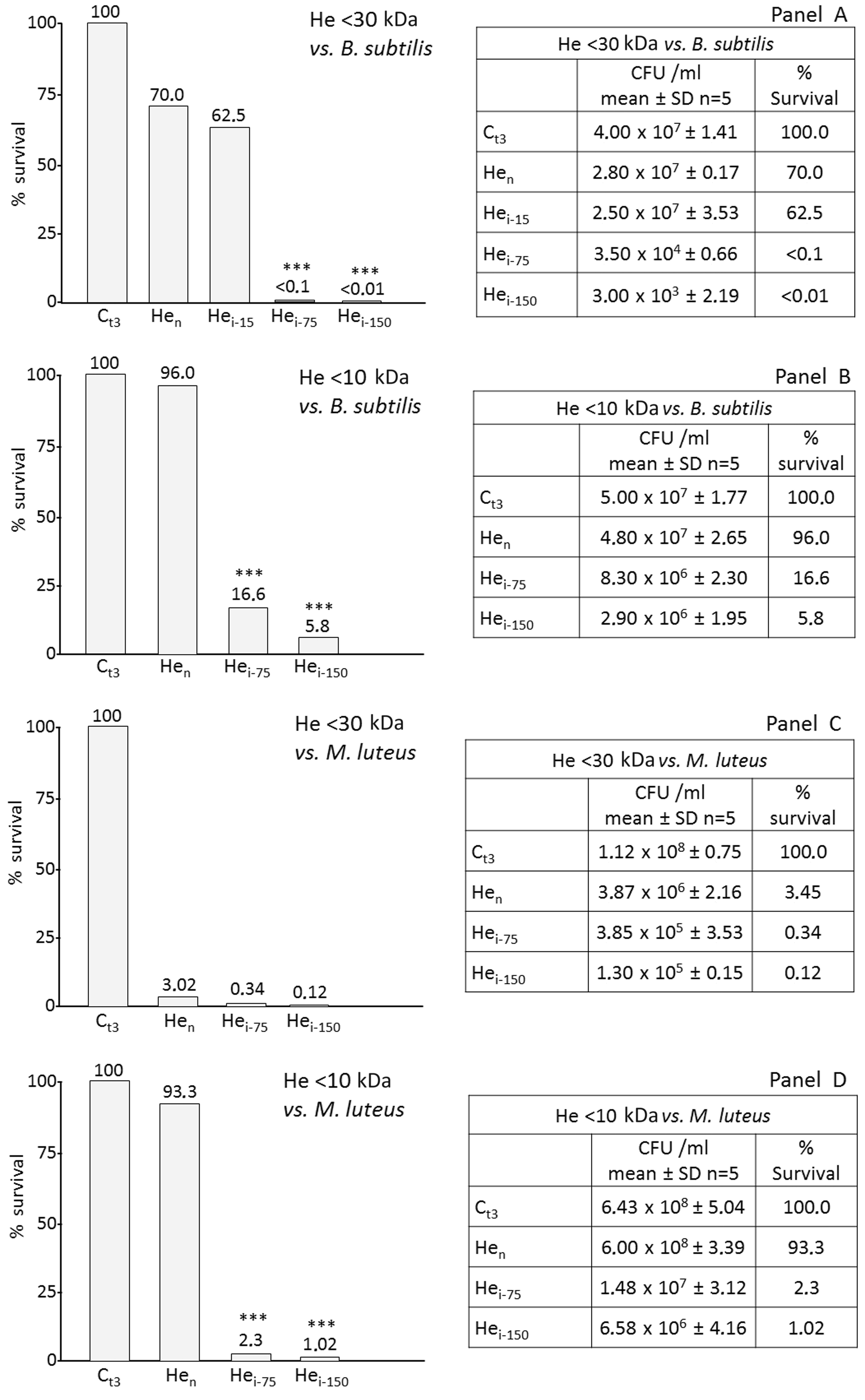
Role of lysozyme on Gram-positive bacteria
As the antimicrobial activity in the <10 kDa sample was not supported by endogenous lysozyme, we carried out assays adding exogenous lysozyme to the fractioned haemolymph.
Figure 5A shows the strong effect of the enzyme when added to <10 kDa naïve larvae haemolymph (Hen). Likewise, when in the presence of AMPs (Hei150 + Ly), a further increase in bacteria mortality is observable compared with the <10 kDa fraction from immunized larvae (Hei-150).
Effects of lysozyme and LHMED added to AMP fractions. LMM haemolymph (He) fractions from RPW larvae were assayed against Gram-positive bacteria (B. subtilis). (A) He samples (150 µg/ml of <10-kDa fraction) were incubated with 106 CFU/ml B. subtilis in the presence of lysozyme (Ly; 2 µg/µl); after 3 h, bacteria survival was evaluated by CFU count. (B) <30-kDa fractioned He samples (150 µg/ml) were incubated with 106 CFU/ml B. subtilis in the presence of 100 mM LHMED. After 3 h, bacteria survival was evaluated by CFU count. Ct3: control (bacteria); Hen: bacteria + He fraction from naïve larvae; Hen + Lin100: bacteria + He fraction from naïve larvae + 100 mM LHMED; Hei: bacteria + He fraction from immunized larvae; Hei + Lin100: bacteria + He fraction from immunized larvae + 100 mM LHMED. Bacterial growth was not affected by the presence of 100 mM LHMED (data not shown). Ct3: control (bacteria); Ct3 + Ly: control bacteria + lysozyme; Hen: bacteria + He fraction from naïve larvae; Hen + Ly: bacteria + He fraction from naïve larvae + lysozyme; Hei-150: bacteria + He fraction from immunized larvae; Hei-150 + Ly: bacteria + He fraction from immunized larvae + lysozyme. Results are expressed as percentage of survival compared with the control Ct3 (100% survival). **P < 0.01; ***P < 0.001.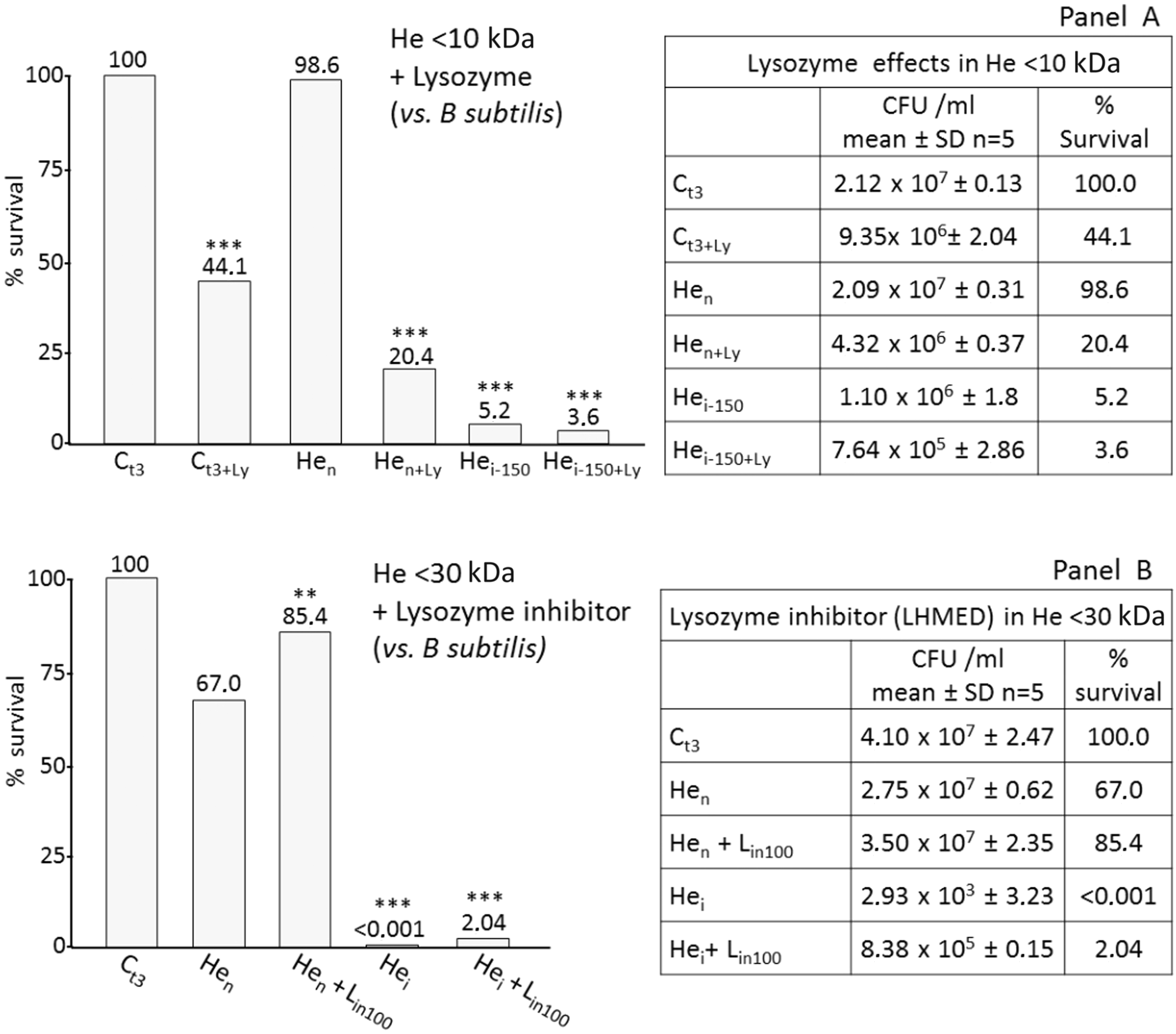
To ascertain if the mortality observed in samples from naïve larvae could be attributed to the presence of constitutive lysozyme, we repeated the assay adding the specific lysozyme inhibitor, LHMED (Figure 5B). Here, the presence of LHMED induced a substantial neutralization of the antibacterial effects, particularly in samples from naïve larvae (Hen + Lin100). Also, the effects were less obvious when the inhibitor was added to samples from immunized larvae (Hei + Lin100).
Haemolytic activity of RPW haemolymph
Human erythrocytes were used as target cells to test the haemolytic activity in RPW haemolymph samples. Haemolymph from naïve and immunized larvae were incubated with RBCs, and cell lysis was assessed as release of haemoglobin in the extracellular environment. Figure 6 shows the results of the lysis tests: whole and fractioned samples did not possess haemolytic activity (Hew, Hen30, Hei30, Hei10) compared with Triton x-100 treatment positive control.
RBC lysis assays. RBC lysis was checked in presence of whole or fractioned haemolymph (He) samples (150 µg/ml). Assays were performed by recording the absorbance at λ = 404 nm owing to the release of haemoglobin in supernantants. The graph shows the percentage of lysis referred to a 100% lysis positive control (C) obtained by RBC incubation with 0.02% Triton X-100. C: control; Hew: RBC + whole He from naïve larvae;Hen < 30, RBC + <30 kDa He fraction from naïve larvae; Hen < 10: RBC + <10 kDa haemolymph fraction from naïve larvae; Hei < 30: RBC + <30 kDa haemolymph fraction from immunized larvae; Hei < 10: RBC + <10 kDa haemolymph fraction from immunized larvae.
Bacterial cell wall damage induced by AMPs
We evaluated damage to bacterial walls by PI uptake: bacteria were treated with haemolymph extracts from naïve and immunized larvae, and fluorescence, due to the penetration of PI inside damaged cells, was visualized (Figure 7).
PI uptake after AMP treatments. Gram-negative (E. coli) and Gram-positive (B. subtilis) bacteria were treated with fractioned (<30 kDa) haemolymph from naïve and immunized larvae. t = 0: start of treatment; t = 3: 3 h after start of treatment; PH: phase contrast; Fluo: fluorescence microscopy. Fluorescent cells in micrographs at right (immunized, Fluo, t = 3) indicate propidium uptake as a result of the damage of cell surface.
Micrographs show the effects of AMPs against E. coli and B. subtilis at t = 0 and 3 h after treatment (t = 3). The effects of antimicrobial peptides resulted in a large number of fluorescent cells for both Gram-negative and Gram-positive bacteria (Figure 7; t = 3, Fluo).
We also observed the bacteria using SEM (Figure 8): cell damage is clearly observable in the right panels; blebs are evident on the cell surface of E. coli (Ect) and B. subtilis cells appear empty and flattened (Bst).
SEM of bacterial wall damage. Micrographs show the alteration of the bacterial wall after AMP treatments (<30 kDa fraction). Left panels are controls of untreated E. coli (Ecc) and B. subtilis (Bsc), right panels are results after 3 h of AMP treatment against E. coli (Ect) and B. subtilis (Bst). Escherichia coli cells show various blebs protruding from the surface (arrowheads); B. subtilis are clearly flattened.
Discussion
Animal and plant kingdoms are a favourable source of drugs, including antimicrobial peptides.26,27 Although antimicrobials developed for clinical use are mainly derived from bacteria, 28 the insect class represents a promising additional source for new biologically active molecules. 29 Some authors have suggested the use of insects as model organisms for pharmacokinetic studies, with the aim of avoiding conventional tests on vertebrates.30–34 However, beyond a possible use as overall experimental models, insects represent a proven source of peptides with antimicrobial activity. For therapeutic purposes, it is also important to consider that cationic AMPs do not trigger antibiotic resistance mechanisms, which often restrict the use of other conventional antibiotics. 35
Insect AMPs could be categorized into the following major groups based on their secondary structure, Aa sequence and antibacterial activity: linear amphipathic α helix-forming peptides (e.g. cecropins), β-sheets or cystine-rich and cyclic antimicrobial peptides (e.g. defensin), and proline-rich peptides and glycine rich peptides (e.g. drosocin and coleoptericin).36–38 A large number of natural AMPs have been identified, and many projects have been aimed to formulate pools of peptides with higher efficacy, rather than to characterize and to design new AMPs-derived molecular analogues. 35
In this work we focused our research on a coleopteran insect model with the purpose of investigating a physiological response that is part of innate immunity, but occurs exclusively following infections, and thus is considered an inducible process that leads to the synthesis ex novo of antimicrobial compounds useful against a broad range of microorganisms. RPW larvae, when immunized with bacterial strains, up-regulate the expression of genes encoding for small proteins and peptides detectable in the haemolymph. To date, few data on the antimicrobial clearance performed by AMPs in R. ferrugineus larvae are available.20,21
AMPs identified in many insects operate synergistically against a wide range of Gram-positive and Gram-negative bacteria. The onset of these molecules occurs after stimulation of the host with both viable and killed bacteria,39–41 but also if larvae are immunized with purified bacterial PAMPs (e.g. LPS or PGN).
Owing to its natural habitat being particularly propitious to bacterial growth, it is expected that the RPW possesses a powerful arsenal of AMPs. The literature describes several coleopteran AMPs. Among them, cecropins, defensin and attacins are not peculiar to this order, but coleoptericin, rhinocerosin and holotricin are described exclusively in Coleoptera. 5
To ascertain the presence of ex novo synthesis of proteins or peptides, and thus a process of post-infection inducibility, we carried out electrophoretical analyses of haemolymph samples from bacteria-challenged and unchallenged RPW larvae. The occurrence of several newly synthesized molecules was examined by means of 2D-PAGE. Putative AMP molecules seem to be present in the fractions<30 kDa. Among them, at least 14 spots were detectable in the range between 4 and 10 kDa, seven of which seemed to be more basic. We detected strong activity against E. coli in whole haemolymph from challenged larvae; however, data from 2D-PAGE led us to focus microbiological assays to fractioned haemolymph samples <30 kDa or <10 kDa, as newly synthesized peptides were observable in the <30 kDa zone.
Similarly, with the aim of defining the range of action of RPW AMPs, we performed tests on both Gram-negative (E. coli, Pseudomonas sp. OX1) and Gram-positive bacteria (B. subtilis and M. luteus). Data obtained with <30-kDa fractions revealed a marked dose-dependent efficacy against both Gram-negative and Gram-positive bacteria. The strong activity of the <30-kDa sample could also be due to the presence of lysozyme, as in many instances the muramidase activity of the enzyme acts synergistically with the AMPs.42,43 The <10-kDa fraction, which lacked lysozyme, showed a lower activity against B. subtilis, with respect to the effects observed either versus Gram-negative bacteria or versus M. luteus. Though E. coli and Pseudomonas sp. seemed to be less sensitive to lysozyme activity, the results with the <30-kDa and <10-kDa fractions were comparable.
According to the literature,44,45 insect lysozyme is also present constitutively in haemolymph samples from unchallenged larvae. This is particularly evident from our data, obtained against M. luteus, with samples of haemolymph from naïve larvae. The <30-kDa fraction, in which only constitutive lysozyme (about 14 kDa) was present, showed a strong activity against this bacteria. We also observed that naïve haemolymph fractions showed moderate antibacterial activity against Pseudomonas sp. and B. subtilis. The synergistic role of lysozyme to improve the efficiency of AMPs has been described exhaustively,44,46–48 and was confirmed by our data. The addition of purified lysozyme to RPW haemolymph lower mass fractions (<10 kDa) strongly increased the mortality of B. subtilis, and when lysozyme inhibitor was added to the <30-kDa fractions, bacterial survival increased.
Most AMPs exert their antibacterial or antifungal effects by interacting with and destabilizing both plasma membranes and bacterial walls, eventually leading to cell death; alternatively, some AMPs can enter the cell and interact with cytoplasmic factors.49–51 We confirmed the action of RPW peptides on bacterial cell walls by evaluating the uptake of intercalating agents. As observed by fluorescence microscopy, PI inside bacteria confirmed the presence of openings occurring in AMP-treated cells. Our SEM observations suggest that RPW antimicrobial pools contain peptides that led to the formation of holes responsible for the flattening of B. subtilis and of the appearance of blebs on E. coli. These data were in agreement with other publications that had explained the occurrence of blebs and pores on the bacterial surface as a consequence of a distortion of the outer membrane resulting from the interaction between AMPs and the anionic surface (LPS or teichoic acids) of bacterial cells.35,50
Many studies on insects have embraced different physiological aspects of this animal class. In this work, we have investigated an immunological process that is part of the internal defence of RPW, that is the inducible humoral response succeeding a significant bacterial infection. RPW spends a considerable part of its life cycle in an extremely warm environment with a high rate of fermentative processes that stimulate microbial growth; its physiology must therefore be able to effectively fight any infection status that may occur.
We believe that our data provide enough information to establish that RPW can counteract the intrusion of microorganisms, triggering the synthesis of antimicrobial peptide pools that are extremely efficient in preventing the onset of septicaemia (Figure 9).
Graphical overview of inducible response in RPW after immune challenge.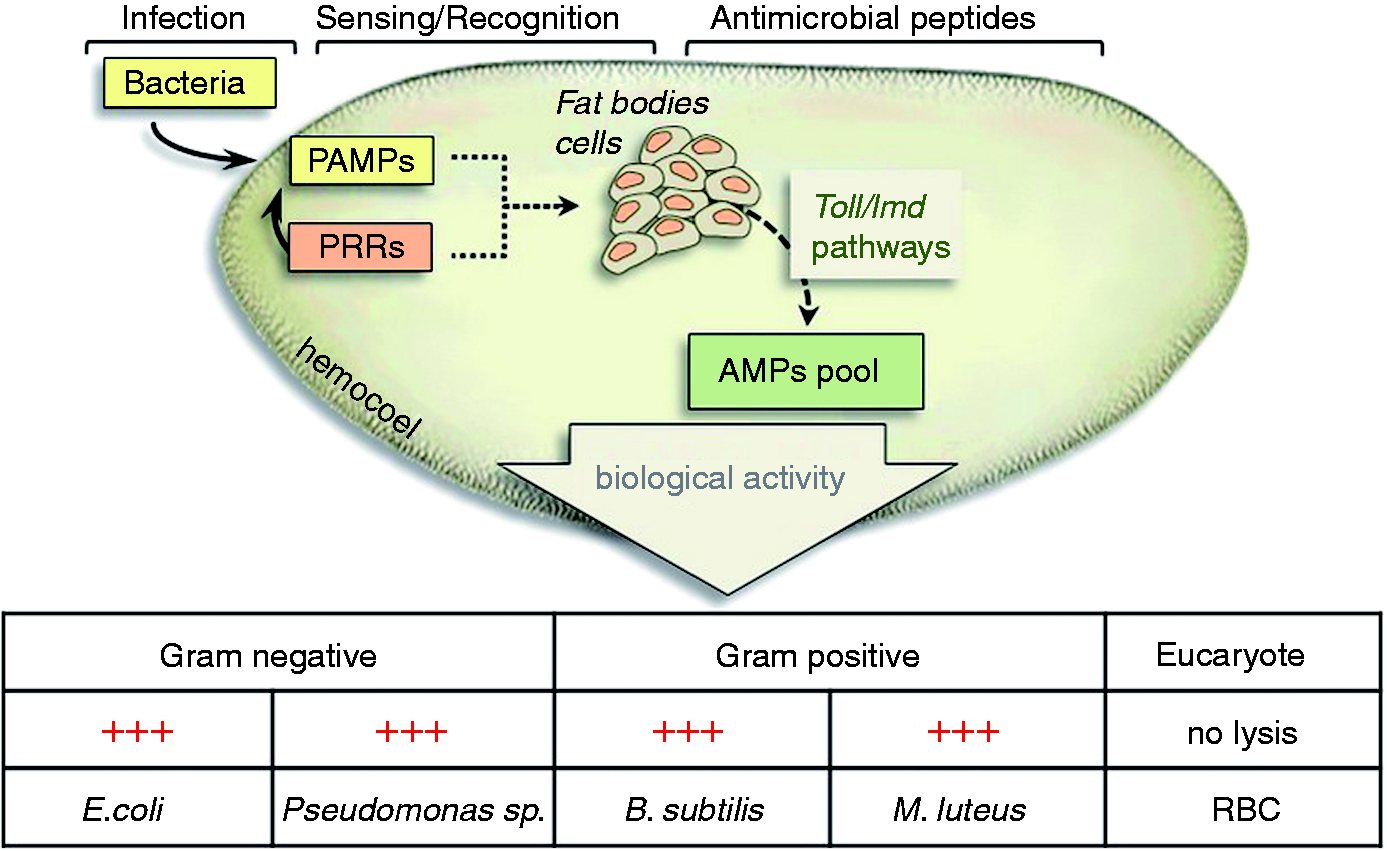
Experimentation with this animal model is favourable as it has several appealing features. The large dimension reached by larval stages makes RPW a good candidate as an alternative model to vertebrates for the study of important physiological processes and their changes in response to physical and pharmacological stresses. Another important feature of RPW, common to all insects, is that its innate immune defence displays many functional similarities to the vertebrate immune system (e.g. sensing mechanisms, the presence of AMPs, the proPO activating system versus human complement, phagocytosis, etc.). RPW larvae could represent a kind of living laboratory in which to identify new compounds with antimicrobial activity that might be useful for the development of innovative drugs of natural origin, able to counteract the antibiotic resistance that currently presents a serious threat to human and animal health.
Footnotes
Funding
This work was supported by funds from University of Insubria.
Acknowledgements
We are grateful to Dr Viviana Orlandi for the microbiological assay method, and to Drs Flavia Pennica and Celeste Re for their assistance during the assays. We express gratitude to the Major of Ribera, Mr Carmelo Pace (Sicily, Italy), for the logistic support.