
Abstract
Macrophages have been reported to initiate immunosuppression following trauma and hemorrhage, and recent experimental studies suggest a pivotal role of T-cells in maintaining immunosuppression. The aim of the present study was to investigate the interaction of APC and T-cells in humans following major surgery. First, APC and T-cells from 14 surgical patients were isolated, counted and characterized by their specific surface marker profile 2 and 24 h postoperatively. Then, these cells were co-incubated with cells of the other type, which had been isolated pre-operatively. Chemokine secretion from pre-operative cells as measured by enzyme immunoassay served as a bioassay for the function of the stimulating postoperative cells. CD3+ T-cells and surface marker CD28 were markedly suppressed postoperatively, while CD3+CD25+CD127−Tregs were not suppressed. CD14+APC counts were increased with the most significant increase observed in CD14+HLA-DR− myeloid-derived suppressor cells. In co-cultures, APC showed increased postoperative secretion of TNF-α and IL-6 independently of whether they had been co-incubated with pre- or postoperative T-cells. T-cells incubated with CD14+ cells 2 h postoperatively secreted diminished amounts of IFN-γ. The results of the study suggest that T-cells play a pivotal role in mediating immunosuppression after major abdominal surgery.
Introduction
Major surgical trauma and blood loss result in a depression of cell-mediated immunity.1,2 In particular, the interaction between APC and T-cells, which has been shown to play a pivotal role in the induction of an adequate cell-mediated immune response, is altered after trauma hemorrhage.3,4 Depression of T-cells and APC has been demonstrated 7 d after surgery and is associated with an increased susceptibility to infection. 5
Various mechanisms in both APC and T-cells are thought to be responsible for an insufficient interaction of the different cell types. In this respect, MHC II expression by APC is suppressed after trauma and blood loss, resulting in decreased Ag presentation. 6 In addition, LPS-induced IL-12 release capacities of APC are diminished after trauma and blood loss. 7 APC-derived IL-12 has been shown to activate T-cell responses, that is, the release of IFN-γ and IL-2. Conversely, the synthesis and release of IL-12 appears to be regulated by T-cell-derived IFN-γ. 8
On T-cells, distinct components of the T-cell receptor (TCR) complex (TCR-α and TCR-β chains,9,10 as well as the invariable CD3 proteins) and co-stimulatory molecules (CD4, CD8 and CD28) have been identified to be required for APC–T-cell interaction. 11 Trauma and surgery lead to a significant decrease in total lymphocyte counts and T-cell proliferation, which is associated with a shift from pro-inflammatory Th1 cytokine production (i.e. IL-2, IFN-γ) to an increased release of potentially immunosuppressive Th2 cytokines (i.e. IL-4, IL-10) mediated by regulatory T-cells (Tregs).2,7 Th2 cytokines increase the expression of arginase-1 in myeloid-derived suppressor cells (MDSCs), causing an arginine deficiency state, which further impairs lymphocyte function.2,12,13
Although numerous studies have been directed towards these immune cells and the distinct processes of their interaction after trauma and surgery, it is not known which immune cell type is predominately responsible for immunosuppression in humans under these conditions. Experimental studies in mice suggest that APC initiate immunosuppression after trauma and blood loss, 14 whereas T-cells play a key role in the perpetuation of diminished T-cell–APC interaction. 15 In particular, trauma-hemorrhaged T-cells depress cytokine responses and the expression of MHC II by APC irrespective of whether APC were subjected to trauma hemorrhage or sham operation.
The aim of the present study, therefore, was to investigate the T-cell–APC interaction in patients after major abdominal surgery in order to clarify the immunosuppressive effect of APC on T-cells in the clinical setting. The results from the present study may help to further elucidate the pathophysiological mechanisms of immunosuppression following major abdominal surgery and thus allow for the development of new targets for specific immunotherapy.
Materials and methods
Study design
This study was designed as a prospective observational trial with consecutive measurements and pre-operative values serving as internal controls. It was conducted in the Department of General, Visceral, Transplantation, Vascular and Thoracic Surgery at Großhadern Hospital, Ludwig-Maximilians University, in Munich, Germany. The study protocol was approved by the local institutional review board.
Study patients
Patient characteristics.

Study protocol
Three times were defined for measurements: 1 h pre-operatively as an internal control, and 2 and 24 h postoperatively (Figure 1). Blood (40 ml) was drawn at these three times and, in addition, 24 h pre-operatively, from all patients. Two types of experiments were then conducted with these samples: first, FACS analysis for surface marker expression at the predefined times was performed; second, co-cultures of T-cells and APC were established from these samples and from untreated 24 h pre-operative blood. Cytokine secretion from these co-cultures was then determined using enzyme immunoassays. The experimental procedures, in detail, were as follows.
Sample acquisition and cell utilization. For FACS analysis, cells were used without further treatment. Co-cultures were established using pre-operative ‘frozen’ together with pre- and postoperative ‘non-frozen’ cells. IL secretion from the pre-operative ‘frozen’ cells served as a bioassay for the function of the pre-operative controls or postoperative cells that they were incubated with.
PBMC isolation
PBMC were separated using a standard Ficoll–Hypaque density gradient (BioChrom AG, Berlin, Germany). Briefly, blood was diluted at a 1 : 1 ratio with Hank’s balanced salt solution (HBSS; Lonza, Walkersville, MD, USA) layered on top of the Ficoll Hypaque in 50-ml tubes and centrifuged for 25 min at 900 g. PBMC were collected using a medical syringe. Cells were then re-suspended in HBSS, counted and stored on ice at 4℃. Viability of PBMC was consistently > 95%, as determined by trypan blue exclusion.
APC and T-cell separation
APC and T-cells were purified in two sequential positive selection steps using magnetic-activated cell sorting kits (MACS; Miltenyi Biotec, Bergisch Gladbach, Germany) with magnetic beads labeling the target cells as described by the manufacturer. Briefly, after centrifugation (5 mins at 400 g), cells were re-suspended in 80 µl MACS buffer and 20 µl CD14 magnetic Mircrobeads (Miltenyi Biotec) per 107 cells and incubated at 4–8℃ for 15 mins. The suspension with labeled CD14+ cells was diluted in 1–2 ml buffer and centrifuged at 300 g for 5 mins. Cells were re-suspended in 500 µl MACS buffer and applied to the MACS column. CD14+ cells labeled with magnetic beads adhered to the column, while non-labeled cells were rinsed with 500 µl MACS buffer three times. Magnet-sorted cells were eluted from the column to a separate vessel using 1 ml buffer after removing the column from the magnetic field.
For CD3+ cell purification, the suspension passed through the column and was thus depleted of CD14+ cells, and was again centrifuged for 5 min at 400 g. Cells were then re-suspended in 80 µl MACS buffer and, this time, 20 µl CD3 magnetic microbeads per 107 cells and incubated at 4–8℃ for 15 mins. This suspension with labeled CD3+ cells was then processed and applied to the MACS column as described above.
To confirm purity, 2 × 105 cells were aliquoted from both the fractions held back and the fractions flown through the column and analyzed by flow cytometry. Sorted cells were > 99.5% pure, as determined by post-sort analysis for CD3+ or CD14+ cells. Purified cells were centrifuged at 400 g for 5 mins and then suspended in 1 ml medium (90% RPMI 1640 GlutaMAX-1 and 10% FCS), counted and stored on ice.
Utilization of separated APC and T-cells
Cells harvested 24 h before surgery were frozen at −80℃ using a buffer system with 70% RPMI 1640 GlutaMax-1, 20% FCS and 10% DMSO for APC and T-cells. Those cells were thawed on the day of surgery or the following day for the establishment of co-cultures. Henceforth, those cells will be named ‘frozen APC’ or ‘frozen T-cells’, respectively (Figure 1).
Separated T-cells and APC harvested pre-operatively on the day of surgery, and 2 and 24 h postoperatively were used for flow cytometry analysis and co-culturing without freezing. Henceforth, these cells are named ‘non-frozen APC’ or ‘non-frozen T-cells’, respectively.
Co-cultures of APC and T-cells
Separated non-frozen or frozen APC (CD14+ cells) and non-frozen or frozen T-cells (CD3+ cells) were co-cultured in 48-well culture plates. For each time point, the designed co-culture system resulted in four experimental groups:
group 1—frozen T-cells + frozen APC (control group); group 2—non-frozen T-cells + non-frozen APC (the ‘normal’ non-frozen situation); group 3—frozen T-cells + non-frozen APC (to study the influence of APC on T-cells); group 4—non-frozen T-cells + frozen APC (to study the influence of T-cells on APC).
Co-cultures were stimulated using LPS for APC stimulation and anti-CD3, as well as anti-CD28 for T-cell stimulation. Three separate plates (pre-operative, and 2 and 24 h postoperative) were prepared for each patient with pre-coated anti-CD3-Abs (250 µl; Miltenyi Biotec) and rinsed three times with 300 µl HBSS.
Co-cultures were incubated in 1 ml of DMEM (GIBCO, Invitrogen, Carlsbad, CA, USA) with 10% inactivated FCS (Cambrex, Verviers, Belgium) and 0.1% Gentamycin (Bio Whittaker, Verviers, Belgium) for 48 h at 37℃ and 5% CO2. In addition to anti-CD3 (pre-coated plates), cells were stimulated with 20 µl anti-CD28 Abs (Miltenyi Biotec) and 1 µg/ml LPS from Escherichia coli 055 : B5 (< 3% protein, TCA extracted; Sigma, St Louis, MO, USA).
After 48 h of incubation, the supernatants were harvested, aliquoted and stored at −70℃ for cytokine measurements.
Assessment of cytokines
IL-6, IL-12, GM-CSF, TNF-α (TNF) and IFN-γ were determined in the supernatants by multiplex assay (BioRad Laboratories, Hercules, CA, USA) as per the manufacturer’s instructions. In brief, for multiplex assay, 50 µl of each sample was allowed to react with the cytokine-specific Ab-conjugated beads in micro-plate wells. The flow-based Bio-Plex protein array system identifies and quantifies each specific reaction based on bead colours according to the manufacturer’s information. The magnitude of the reaction was measured using fluorescently-labeled reporter molecules, also specific to each target protein.
Flow cytometry
Abs used for FACS analysis.
Statistical analysis
Data are presented as mean ± SEM. Measurements from samples acquired 2 and 24 h postoperatively were compared with pre-operative measurements. ANOVA was used to adjust for these multiple comparisons. The Holm–Sidak post hoc test was used where appropriate. SigmaStat (v3.0; Systat, Chicago, IL, USA) software was used for statistical analysis. A P-value of ≤ 0.05 was deemed significant.
Results
Cell surface markers
CD3+ T-cells were significantly suppressed 2 h (−38%; P < 0.001) and 24 h (−23%; P < 0.001) postoperatively, while CD14+ APC were induced at 2 h (+64%; P < 0.001) and 24 h (+71%; P < 0.001).
Surface markers on T-cells
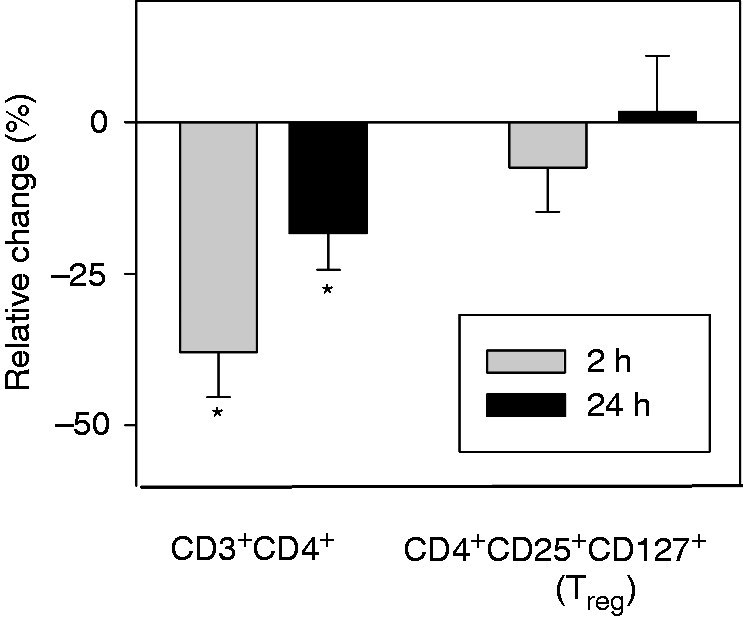
The subset of CD3+CD28+ cells was suppressed 2 and 24 h after surgery as compared with pre-operative measurements. In addition, the CD3+CD28+/CD3+ ratio was significantly reduced 2 and 24 h postoperatively, that is, the expression of surface marker CD28 was significantly reduced on CD3+ cells following surgery. Counts of CD3+CD25+ cells were similarly reduced 2 and 24 h postoperatively, with a significant drop in the CD3+CD25+/CD3+ ratio at 2 h, but not 24 h. Finally, fewer CD3+CD69+ cells were found postoperatively, although this decrease was not significant. The CD3+CD69+/CD3+ ratio did not change postoperatively.CD3+CD4+ T-helper cells were significantly suppressed 2 and 24 h after surgery (Figure 2) with no change in the CD3+CD4+/CD3+ ratio. Within the group of T-helper cells, the subgroup of CD4+CD25+CD127− Tregs remained unchanged compared with pre-operative cell counts (Figure 3). However, the CD4+CD127+CD25+/CD3+ ratio was significantly increased at 2 h.
Postoperative changes in CD3+ subpopulations. This graph shows relative changes in CD3+ subpopulations compared with pre-operative measurements. Most CD3+ subpopulations are suppressed postoperatively, at 2 h more than at 24 h. Of all subtypes analyzed, suppression of CD3+CD4+, CD3+CD28+ and CD3+CD25+ cells was the strongest, while suppression of CD3+CD69+ cells was statistically not significant. *P < 0.05 compared with pre-operative values. CD4+ subgroup analysis. As with other CD3+ cells, CD3+CD4+ cells are suppressed postoperatively. The subgroup of CD4+CD25+CD127– Tregs, however, are not suppressed. *P < 0.05 compared with pre-operative values.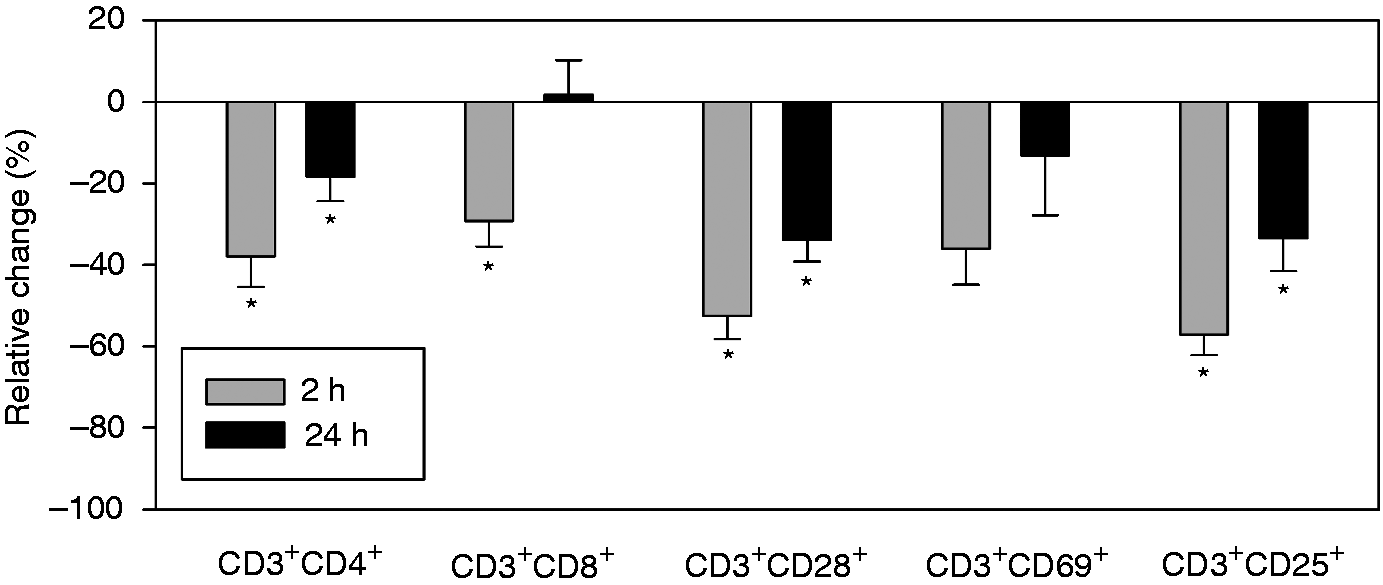
CD3+CD8+ cells were significantly suppressed only at 2 h, but not at 24 h, while no change in the CD3+CD8+/CD3+ ratio was found. The CD4+/CD8+ ratio was only slightly reduced after surgery (P > 0.05).
Surface markers on APC (Figure 4)
The CD80+ subset of CD14+ APC was significantly increased 2 and 24 h postoperatively, with no significant change in the CD14+CD80+/CD14+ ratio. Similarly, CD14+CD86+ cells were induced 2 and 24 h postoperatively. Here, a transient increase of the CD14+CD86+/CD14+ ratio was observed. Postoperative counts of CD14+HLA-DR+ cells were also significantly higher when compared with pre-operative values with no significant change in CD14+HLA-DR+/CD14+ ratios. The largest increase of all CD14+ cells analyzed was observed at both 2 and 24 h after surgery for CD14+HLA-DR–MDSC.
Postoperative changes in CD14+ sub-populations. CD14+ cells were induced postoperatively. The most important increase was observed for CD14+HLA-DR– MDSC. *P < 0.05 compared with pre-operative values.
CD14+CD16+ cells were significantly induced 24 h, but not 2 h, after surgery, while the CD14+CD16+/CD14+ ratio showed a transient, but significant, decrease 2 h postoperatively. CD14+CD40+ cells were increased 2 and 24 h after surgery, with no change in the CD14+CD40+/CD14+ ratio.
Cytokine secretion from co-cultures of T-cells and APC
APC-derived cytokines
Secretion of IL-6 (Figure 5) and TNF (Figure 6) by APC harvested postoperatively and co-cultured with postoperative T-cells (group 2) was significantly increased 2 and 24 h after surgery as compared with pre-operative measurements. When postoperative APC were co-cultured with frozen T-cells separated pre-operatively (group 3), secretion of IL-6 and TNF was induced comparably to the co-culture system using only postoperative cells. Secretion of IL-6 or TNF by pre-operatively frozen APC was not increased when co-cultured with non-frozen T-cells (group 4). The secretion of these cytokines was comparable to the control co-culture-system using only frozen APC and T-cells (group 1), and to pre-operative values.
IL-6 production was induced in co-cultures of frozen or non-frozen T-cells with non-frozen APC, but not with frozen APC. T: T-cells; M: APC; f: frozen. *P < 0.05 compared with pre-operative values. TNF production was induced in co-cultures of frozen or non-frozen T-cells with non-frozen APC, but not with frozen APC. T: T-cells; M: APC; f: frozen. *P < 0.05 compared with pre-operative values.

The secretion of IL-12 (Figure 7) did not change postoperatively (group 2) and was not affected by co-culture with frozen T-cells (group 3). It was reduced to almost undetectable levels in both co-culture systems containing frozen APC (groups 1 and 4). This reduction was also observed in pre-operative controls that had been frozen on the day of surgery.
IL-12 production was reduced to undetectable levels when frozen APC were used in co-cultures. T: T-cells; M: APC; f: frozen. *P < 0.05 compared with pre-operative values.
T-cell-derived cytokines
IFN-γ from postoperative T-cells co-cultured with postoperative APC (group 2) was significantly decreased at 2 h, but not at 24 h (Figure 8). When frozen T-cells were co-incubated with postoperative APC (group 3), IFN-γ secretion was transiently and not significantly suppressed at 2 h. Similarly, the transient suppression of IFN-γ secretion at 2 h from non-frozen T-cells cultured with frozen APC (group 4) was not significant.
IFN-γ production was suppressed 2 h postoperatively in co-cultures of T-cells and APC. Suppression was significant when non-frozen T-cells and APC were used but not when either T-cells or APC had been frozen. T: T-cells; M: APC; f: frozen. * P < 0.05 compared with pre-operative values.
At the same time, granulocyte macrophage colony-stimulating factor (GM-CSF) secretion was increased at 2 and 24 h when postoperative (group 2) or frozen (group 3) T-cells were co-cultured with postoperative APC (Figure 9). When frozen APC were used (groups 1 and 4), GM-CSF secretion did not increase.
GM-CSF production was induced in co-cultures of frozen or postoperative T-cells with postoperative APC but not with frozen APC. T: T-cells; M: APC; f: frozen. *P < 0.05 compared with pre-operative values.
No significant postoperative changes for any cytokine were found in controls (group 1), that is, co-cultures of frozen T-cells with frozen APC.
Discussion
Abdominal surgery with blood loss causes a depression of cell-mediated immune responses resulting in an increased susceptibility to infectious complications with increased mortality. 2 This immunosuppression affects, among other immune cells, both T-cells and APC. Th1-cells, for example, proliferate less and release smaller amounts of pro-inflammatory cytokines, such as IL-2 and IFN-γ.3,16 In addition to decreased cytokine release capacities, APC have been found to exhibit diminished MHC II expression and, thus, reduced Ag presentation. 6
The complex interaction between these two cell types plays an important role in the maintenance of regular immune function. 14 Experimental findings suggest that following trauma hemorrhage, defective T-cells suppress APC and vice versa. Kawasaki et al. 17 found dendritic cells isolated 2 h after trauma hemorrhage to cause diminished T-cell responses in vitro, while anergic T-cells have been shown to suppress dendritic cell function. 18 Similarly, recent studies by our group using a co-culture system following immune cell separation demonstrated that trauma-hemorrhaged T-cells are able to depress IL-12 release by APC. 3 However, the release of the T-cell cytokine IFN-γ, 3 as well as IL-4 and GM-CSF secretion, 15 remained unaffected by trauma-hemorrhaged APC. These results suggest that IFN-γ and other T-cell-derived cytokines contribute to immunosuppression following trauma hemorrhage causing diminished MHC II expression on APC in an experimental setting. To our knowledge, these interactions have not yet been investigated in humans undergoing major abdominal surgery.
Therefore, the aim of the present study was to investigate the changes in APC and T-cells, and how the two cell types influence each other following surgical trauma in abdominal surgery patients. To address this question, we measured surface markers on T-cells and APC, as well as cytokine responses in a co-culture system of the two cell types.
In this study, the number of patients was too small to correlate the amount of blood loss with immune responses. Moreover, no correlation between blood loss and minimum blood pressure (R = 0.22), vasopressor dose (R = –0.07) or lactate (R = 0.18) in our study patients was found. This suggests that that blood loss was probably too low and fluid resuscitation adequate to prevent hemorrhagic shock to occur.
Determining the effect of intra-operative blood loss on immune cell function was beyond the scope of the present study owing to the limited number of patients with excessive and small blood loss.
FACS analysis showed that CD3+ T-cells were suppressed postoperatively, with the lowest values observed early at 2 h. Various CD3+ subpopulations were likewise diminished. Interestingly, the surface molecules CD25 (at 2 h) and co-stimulatory CD28 (at 2 and 24 h) were reduced significantly not only when compared with pre-operative values, but also when compared with the overall CD3+ postoperative population. CD25 is the IL-2 receptor α-chain 19 and serves as an activation marker for T-cells. 20 CD28 is a co-stimulatory surface molecule on T-cells that may also activate dendritic cells via CD80 and CD86 in a reciprocal fashion. 11 The suppression of activation marker CD69 may not have been significant owing to the small sample size. These results indicate a suppression of T-cell activation occurring early in the post-traumatic period.
The expression of CD4+, CD25+ and CD127– defines a subset called Tregs. Tregs can suppress Th1 immune responses and promote a shift toward Th2 anti-inflammatory immunosuppressive responses.2,21 Interestingly, these CD4+CD 25+CD127− cells were not suppressed postoperatively in our patients. On the contrary, when compared with overall CD3+ cell counts, we observed a significant relative increase of Tregs 2 h postoperatively. Tregs, therefore, appear to be induced after abdominal surgery. An increase of Tregs has been described previously for trauma patients, 22 and was associated with Th2 anti-inflammatory activity. Treg activation may be part of the explanation for decreased IFN-γ release capacities in our patients.
The CD4+/CD8+ ratio was reduced postoperatively in the present study. This result was not statistically significant, but in line with previous results from our group, 15 and typical of trauma patients. 23
In contrast to CD3+ T-cells, CD14+ cell counts and various subpopulations were persistently increased after surgery, with the highest values observed at 24 h. At 2 h, the surface molecules CD16 and co-stimulatory CD86 were induced significantly when compared with the overall CD14+ population. CD16 is an Fc receptor that induces CD25 transcription and thus plays an important role in the interaction of T-cells and APC. 24 CD86 is a ligand for CD28 on T-cells and provides co-stimulatory signals necessary for their activation and survival.
MHC II expression was likewise induced, an observation contrary to a previous study. 6 This absence of MHC II suppression may be owing to the relatively mild hemorrhage in our patients who did not experience massive hemorrhagic shock. Alternatively, MHC II expression may have been caused by direct stimulation with LPS in the ensuing experiments.
The strongest induction of any cell type was observed for CD14+HLA-DR− cells. This subset belongs to the so-called MDSC, 25 which have been shown to be up-regulated after trauma.13,26 Increased production of arginase-1 by MDSC results in depleted arginine levels. 2 As arginine is required for lymphocyte proliferation and the formation of the T-cell receptor, 27 the induction of MDSC provides a mechanism for impaired T-cell immunoproliferative responses and cytokine production in postoperative patients. Arginase-1 may also be produced by alternatively-activated macrophages under the influence of Th2 cytokines or regulatory macrophages. 28 As macrophage differentiation and activation typically takes more than 48 h, it probably did not occur in the present study, but cannot be excluded without positive identification of the cell types involved.
In co-culture experiments, secretion of IL-6 and TNF from macrophages after surgical trauma was increased independently from co-incubation with pre- or postoperative T-cells. This implies a T-cell-independent mechanism for macrophage activation and cytokine secretion by these cells.
As for IFN-γ, secretion was suppressed 2 h after surgery when either postoperative T-cells or APC were co-incubated with the lowest values observed when both cell-types had been collected postoperatively. At the same time, GM-CSF secretion was significantly increased. This indicates that T-cell suppression may specifically concern IFN-γ, but not GM-CSF secretion, as observed in previous animal studies. 4
The interpretation of co-culture results is limited by unspecific effects that the freezing process might have had on APC. IL-12 and TNF secretion was significantly suppressed in all systems that contained frozen APC irrespective of the T-cells used. We were, therefore, not able to reproduce IL-12 measurements from our previous animal study. 3 Damage to APC during the freezing or thawing process must therefore be assumed, and conclusions about the effect of postoperative T-cells on IL-12 release from APC cannot be drawn from these data. The duration of freezing, nonetheless, had no influence on cytokine secretion. The fact that T-cells were suppressed in spite of insufficient IL-12 release implies an IL-12-independent mechanism for T-cell suppression. Apparently, freezing did not alter T-cell function, as observed by preserved IFN-γ and GM-CSF secretion.
Aiming for maximum stimulation of both APC and T-cells, all co-cultures in the present study were stimulated with both LPS and anti-CD3/anti-CD28. As LPS is able to stimulate T-cells via TLR4, 29 T-cell function in co-cultures may have been stimulated by LPS independent of their interaction with APC. Future studies should account for this by stimulation with either LPS or anti-CD3/anti-CD28, depending on the target cell that is being examined.
In conclusion, in the present study, T-cells were suppressed early after major abdominal surgery, while APC were persistently activated independently of T-cell function. T-cell suppression was associated with increased Treg activity and a marked induction of CD14+HLA-DR− MDSC. While cytokine secretion by APC in co-cultures was significantly increased postoperatively, only a transient decrease at 2 h not reflecting the numeric decrease could be demonstrated for T-cell-derived IFN-γ. Further studies are needed to define the role of specific surface markers and cytokines in postoperative immunosuppression.
Footnotes
Funding
This research received no specific grant from any funding agency in the public, commercial or not-for-profit sectors.