
Abstract
Activation of the innate immune system and acute phase response (APR) results in several responses that include fever, metabolic adaptations and changes in behavior. The APR can be modulated by many factors, with stress being the most common. An elevation of stress hormones for a short duration of time can be beneficial. However, elevation of stress hormones repeatedly or for an extended duration of time can be detrimental to the overall health and well-being of animals. The stress and APR responses can also be modulated by naturally-occurring variations, such as breed, gender, and temperament. These three natural variations modulate both of these responses, and can therefore modulate the ability of an animal to recover from a stressor or infection. Understanding that cattle have different immunological responses, based on naturally occurring variations such as these, may be the foundation of new studies on how to effectively manage cattle so that health is optimized and production is benefited.
Introduction
The acute phase response (APR) is defined as the body's early defense mechanism in response to trauma, inflammation or infection, and is facilitated by a cascade of systemic physiological reactions.1–3 Initially, pro-inflammatory cytokines (i.e. TNF-α, IL-6 and IFN-γ) are produced by activated leukocytes to induce the APR by stimulating changes in target cells to combat infection, while the location, type and combination of these proteins determine the final inflammatory outcome.4–6 Typically, the response includes fever, changes in metabolism, and altered behavior accompanied by the hepatic production of various acute phase proteins (haptoglobin, serum amyloid A, C-reactive protein, fibrinogen and others) in response to trauma or infection. This non-specific component of immunity is defined as innate immunity, and serves as the first line of defense against bacterial, viral, protozoal or fungal pathogens. 7 Whether the APR is beneficial and appropriate to survival depends on the magnitude of the immunological challenge and the extent to which an animal's body can regulate the response to return to a homeostatic state. 8 Additionally, stress, which is defined as the state or perception of threatened homeostasis, 9 has been demonstrated to affect the APR through release of glucocorticoids that stimulate or inhibit, depending on the length of exposure, nearly all components of the innate immune response.10,11 Stress hormones (i.e. the catecholamines and glucocorticoids) can generate an APR similar to that which occurs when an animal reacts to an invading pathogen or tissue injury and trauma; 12 therefore, stress hormones, such as cortisol, are important components to consider when observing characteristics of the APR. While the APR is essential for animal survival, it appears there are naturally occurring variations not only between animal species, but within species that serve as modulators of innate immunity and the stress response. Differences in the APR attributed to gender, breed and temperament exist, and understanding these natural variations, in addition to potentially others, may allow for alternative management strategies to overcome barriers that these distinctions present.
Stress and the APR
The general concept that chronic stress has negative impacts on the health, well-being and productivity of animals is widely accepted among livestock producers and the scientific community alike. Stress in an animal can be indicated by transient increases in the hormone cortisol (or corticosterone in rodents), which is the primary glucocorticoid released from the adrenal glands during times of stress, and is easily measured in serum via standard laboratory tests. Cortisol can modulate a variety of biological effects in the body, namely, for the purposes of this review, overall immune function, and in cases of acute stress, can prime the immune system to prepare for the potential invasion of pathogens and subsequent infection. 7 Acting to suppress the APR during chronic stress, cortisol prevents excessive stimulation of the immune system by preventing the release of pro-inflammatory cytokines, decreasing B-cell Ab production and reducing inflammation, which could prove beneficial or detrimental to the animal, depending on a variety of factors. Release of glucocorticoids and catecholamines is considered a key component of the stress response, and the resulting increase in these stress hormones inhibits several pro-inflammatory cytokines (IL-6, IFN-γ and TNF-α).13,14 Glucocorticoids also cause a shift from a primarily Th-1 effector-driven response to favor a Th-2-mediated humoral immune response 15 by inhibiting the production of, and responsiveness to, certain pro-inflammatory cytokines.16,17
Acute versus chronic stress
Acute stress occurs when an animal experiences a stressor for a short period of time, and is associated with the ‘fight or flight’ response and priming of the immune system, which promotes adaptation to a short-term stressor. Chronic stress, however, manifests when an animal experiences a prolonged insult to its homeostatic state, shifting the stress response from one that is preparatory to one that is suppressive across the entire immune system. 7 In contrast to chronic stress, during a response to an acute stressor an animal initiates restraining forces to prohibit an over-reaction from the central and peripheral components of the stress system. The anti-growth, anti-reproduction, catabolic and immunosuppressive effects of the acute stress response are intended to be of limited duration—temporarily beneficial to the individual. Prolonged subjection to stress and, subsequently, prolonged secretion of glucocorticoids can lead to the development of pathological conditions. This shift from acute to chronic stress is dependent upon the animal and its perception of the stressor, the duration of the stressor and as its ability to overcome a stressful event based on previous exposure, genetics, gender, temperament and other contributing factors.9,18
Glucocorticoids and catecholamines have combined effects on monocyte/macrophage and dendritic cells to inhibit components of innate immunity and the APR, while promoting the production of Th-2 humoral-related cytokines, suggesting that stress-induced immune dysfunction primarily involves the innate immune system. 19 Recent research investigating the interaction between stress and immunity suggests that the APR produced by cattle in response to a pathogen is dependent upon natural variations associated with gender, breed and temperament.
Sexual dimorphism of the APR
Sexually dimorphic APRs have been demonstrated in human and animal models, suggesting that gender plays a role in an animal's ability to withstand infectious disease.20–22 These differences are attributable to steroid hormones, specifically estrogens, androgens and progestins, that have immunomodulatory effects.23,24 The specific role that different sex steroid hormones play in the APR is not completely clear; however, in vivo Ab production in human and animal models is enhanced by estrogen.25,26 Conversely, androgens tend to suppress certain aspects of the immune response. 27
Effects of estrogens on innate immunity
The female reproductive tract contains a full complement of immune cells involved in innate immunity. Yet, in order for reproductive functions such as spermatozoa or the developing fetus to thrive, local immunity within the uterus must be down-regulated to ensure survival of spermatozoa and the subsequent fetus. Therefore, increases in the female sex hormones estradiol and progesterone regulate the concentration and activity of leukocytes and other components of innate immunity in the reproductive tract throughout the reproductive cycle.28,29 Changes in the production of natural antibacterial components, recruitment of phagocytic cells, including neutrophils, macrophages and dendritic cells, as well as changes in NK cell function and concentration occur throughout the estrous cycle. Regarding inflammation, estradiol is considered anti-inflammatory because it has been reported to suppress pro-inflammatory cytokine production and inflammatory cell recruitment.30,31 Estradiol and progesterone also influence the APC functions of dendritic cells, and estradiol may influence dendritic cells to stimulate Th-2 responses, while simultaneously causing a decrease in production of the Th-1 cytokines, TNF-α and IFN-γ. 32 Further evidence of sex-related regulation exists in the mouse model; female mice mount a more vigorous T-cell response and produce more Abs than males when immunized.33,34 Kahl et al. 35 reported a greater APR after LPS challenge during estrus compared to diestrus in heifers, suggesting that the difference in the magnitude of pro-inflammatory response between phases of the estrous cycle may be due to the endocrine environment, particularly progesterone and estradiol concentrations. Further, existing evidence suggests that disease severity decreases during pregnancy, particularly during the third trimester when progesterone and estradiol concentrations are highest, while subsequent disease incidence is increased post-partum when these hormone concentrations are less.36,37
Effects of androgens on innate immunity
The endocrine system controls the development and expression of ornamentation, behavioral displays, and obligatory sperm production via testosterone and other adrenergic steroids in males.38,39 In terms of mating success, high concentrations of testosterone represent libido; however, components of innate immunity, such as pro-inflammatory cytokines, are known to adversely affect male ornamentation, behavioral displays and sperm quality.40,41 While testosterone has its reproductive benefits, it has also been shown to cause immune dysfunction.42–44 Androgens may have stimulatory or suppressive effects on various components of the APR, depending on animal species. Testosterone suppresses immune cell differentiation and macrophage activation in mice and rats, 23 whereas production of IL-2 and IFN-γ in peripheral T-cells increased following castration of male rats, improving their ability to overcome viral and bacterial infection. 45 However, in hamsters, testosterone had a direct stimulatory action on immune cells in vivo and in vitro, as evidenced by an increase in cell-mediated immunity. 46 Likewise, other studies suggest that immune function is positively correlated with increases in testosterone.47,48 Kahl and Elsasser 49 reported greater serum concentrations of TNF-α and serum amyloid A in steers administered testosterone via implant (a growth promotant commonly used in beef production) following LPS challenge, but no difference in clinical signs were observed between steers administered testosterone and control animals. Interestingly, in a subsequent LPS challenge on the same animals, the TNF-α response was not different between control and testosterone-treated steers. Yet, there was a prolonged acute phase protein response in steers administered testosterone for a second time. Nevertheless, the effects of androgens on the APR in cattle are not as clear as the regulation of the APR by female sex hormones.
Sexual dimorphism of the APR in cattle
Gender variations of the inflammatory response in cattle.
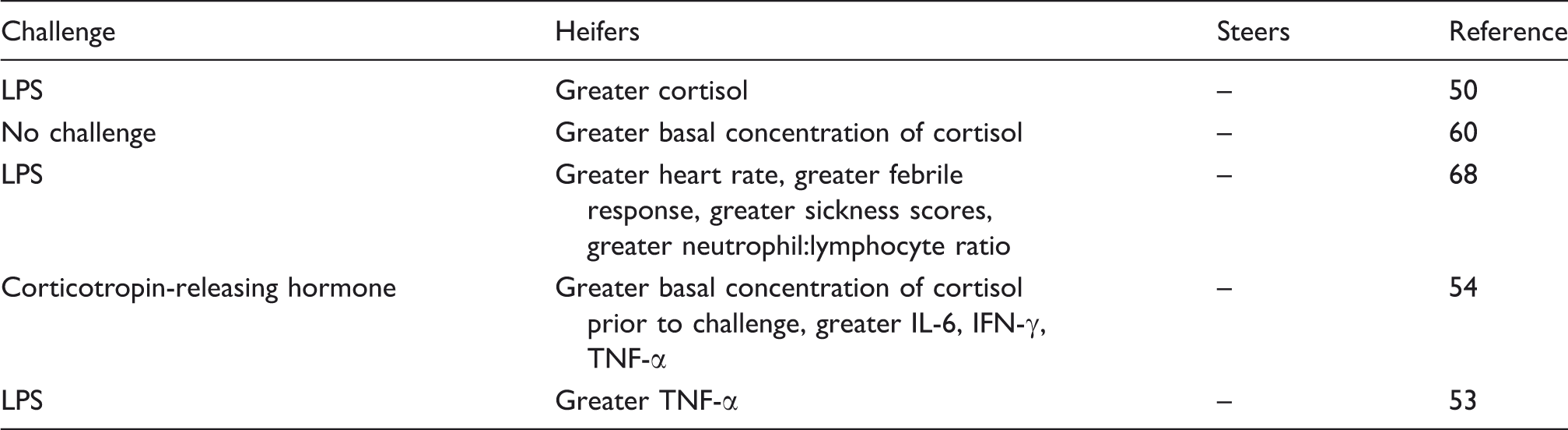
Genetics and the APR
Genetics play an important role in shaping the physical and physiological characteristics of an animal, and studies have reported differences among breeds in multiple species. In cattle, Bos indicus and certain Bos taurus breeds, such as Senepol, are more resistant to heat stress than breeds developed in Europe.59,60 Different breeds of mice have shown variations in production of heat shock proteins and sensitivity to heat-induced neural-tube defects. 61 Henken et al. 62 observed differences in energy and protein metabolism among varying breeds of pigs, as evidenced by differences in growth rate and feed conversion. Therefore, differences in various characteristics, such as heat tolerance, growth traits and carcass traits are recognized. Perhaps less understood are variations in stress and the innate immune responses as a result of breed type.
Stress response variations attributable to breed
Research has shown that genetic differences exist associated with stress responsiveness. Differences have been reported for indices of the APR between Angus and Brahman × Angus cattle when exposed to shipping stress, with Angus steers having greater total leukocytes and skin-test responses to phytohemagglutinin (PHA) than Brahman × Angus steers. 63 Similarly, Angus calves had greater total IgG and IgM titers against pig red blood cells, and greater lymphocyte proliferation in response to PHA compared with Simmental calves, 64 which is indicative of a greater adaptive immune response as a possible outcome of a greater APR and, therefore, stress response. Reed and McGlone 65 reported differences in neutrophil chemotaxis in two commercial lines of pigs when kept in an outdoor environment; however, no differences were observed between breeds when housed indoors. Greater post-stress adrenocorticotropic hormone (ACTH) concentrations after exposure to a novel environment were reported in Large White pigs when compared with Meishan pigs, 66 and while no immune parameters were evaluated, it is plausible that the immune responses of these breeds may have differed. In studies by Sutherland et al.,67,68 which reported numerous breed effects on immune components, the authors did not report any breed × stressor effects on immunity of pigs exposed to heat stress and crowding. Recent studies in cattle, however, have reported variations in the stress response between breeds. In these studies, physiological and blood serum differences were evaluated in heat-tolerant (Romosinuano) and heat-susceptible (Angus) Bos taurus cattle during a controlled heat challenge, with Angus cattle exhibiting greater respiration and sweat rates than Romosinuano cattle. 69 In a similar study observing the same two breeds (Romosinuano and Angus), Carroll et al. 70 reported greater cortisol responses in Angus cattle following LPS challenge, suggesting that Angus cattle more closely regulate the APR than Romosinuano cattle, given that cortisol serves as a potent anti-inflammatory hormone. These variations among breeds in response to stress and immune challenge may indicate a need to manage these breeds differently.
Breed effects on the APR
Genetics can also affect the APR in animals. Recent efforts have been made to select animals for greater disease resistance to improve health, performance and overall productivity, which requires characterization of immune parameters among varying breeds. 67 Genetic selection for Ab or adrenal responsiveness and its relationship to disease responsiveness has been implemented in mice, 71 poultry72,73 and pigs.74,75
In pigs, breed effects have been reported to have an effect on immune traits and cortisol in response to restraint stress, 76 exposure to a novel environment 66 and bacterial challenge. 77 This is consistent with earlier research conducted by Rothschild et al. 78 and Meeker et al. 74 who reported breed differences in immune responses to different antigens in swine. Two commercial lines of pigs demonstrated differences in NK cell cytotoxicity and the lymphocyte proliferative response, 65 while, more recently, Sutherland et al.67,68 reported differences in neutrophil phagocytosis and NK cell cytotoxicity for different breeds of pigs.
Breed variations of some immune and stress responses in cattle.

Temperament and the APR
Definition of cattle temperament
Cattle temperament is defined as the reactivity to humans and novel environments, 83 and is inherently linked to stress in that it involves an animal's response to physical and psychosocial stressors associated with livestock management procedures.84–86 Temperament can be influenced by breed, gender, age and previous handling.87–90 For example, Bos indicus and Bos indicus-crosses tend to be more high-strung or temperamental than Bos taurus cattle,83,91 and steers are typically calmer than heifers. 92 Additionally, heritability of exit velocity from a chute, and chute score, which are measurements of temperament, are affected by breed.93,94 However, extensive handling early in life, and repeated exposure to the same stimuli over extended periods of time, can improve the temperament of cattle. Under the same conditions of transportation, lairage and management of slaughter, improving cattle temperament could potentially reduce the negative effects of temperament on carcass quality.90,95,96 Social interactions among cattle can also affect behavior and temperament, especially if isolated rather than raised in a group. 97
Effects of temperament on the stress response
Temperament can be described as animal's physical response to various stressors, and the stress response interacts with the APR; therefore, it is logical to speculate that differences in an animal's temperament would have an impact on the APR in some cases. In cattle and mice, secretion of various stress hormones have been positively linked to temperament,98,99 and temperamental cattle tend to have greater basal concentrations of cortisol than calm cattle,90,100,101 which may indicate that cortisol acts to protect temperamental cattle facing immune challenge by mitigating the APR and allowing the animal to more quickly return to a homeostatic state. In mice, those showing high locomotion behaviors also had larger adrenals, greater concentrations of glucocorticoids and lesser concentrations of TNF-α following tail nicking, indicating greater hypothalamic–pituitary–adrenal (HPA) axis activation and, subsequently, a suppressed pro-inflammatory response. 102 Curley et al. 98 observed greater basal concentrations of cortisol in temperamental heifers; however, following ACTH and CRH challenge, temperamental cattle had a lesser response than calm cattle, demonstrating that characteristics of the stress response vary with temperament. When challenged with LPS, temperamental Brahman bulls had lower rectal temperature, less sickness behavior and failed to produce an epinephrine response, though it is not completely clear whether this is beneficial or detrimental to the health of temperamental animals. 99 The lesser sickness behaviors, lower rectal temperature responses and greater basal cortisol concentrations appear to contradict previous assertions that a greater or more robust APR is beneficial to the health of an animal. It still remains unclear how temperamental cattle appear to be less affected by an inflammatory challenge; however, it has been hypothesized that this is owing to differences in energy utilization. 103 Additionally, as temperamental cattle produce a blunted response to HPA axis activation it is possible that the greater circulating cortisol concentrations observed in temperamental cattle alter the cytokine response during inflammation. Further research is necessary in order to fully understand the influence of temperament on the APR following endotoxin exposure.
Effects of temperament on the immune response
Variations of stress and immune responses due to temperament in cattle.
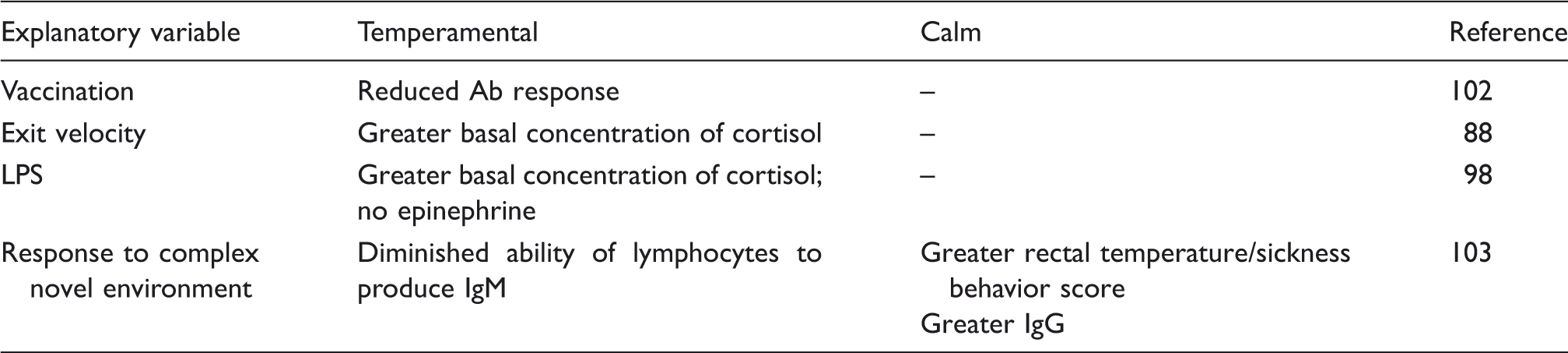
Conclusions
The APR involves the production of pro-inflammatory cytokines and acute phase proteins, increases in body temperature and behavioral changes, and is critical to an animal's maintenance of homeostasis. A homeostatic balance is necessary for optimal livestock production, where growth of the animal is a priority. Natural variations in the APR due to gender, breed and temperament are well documented. These natural variations also serve as modulators of the stress response, which underlies a different impact of stress on innate immune function. Understanding that cattle have different immunological responses, based on naturally occurring variations such as these, may be the foundation of new studies on how to effectively manage cattle so that health is optimized and production is benefited.
Footnotes
Acknowledgements
Mention of trade names or commercial products in this article is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture. The U.S. Department of Agriculture (USDA) prohibits discrimination in all its programs and activities on the basis of race, color, national origin, age, disability, and where applicable, sex, marital status, familial status, parental status, religion, sexual orientation, genetic information, political beliefs, reprisal, or because all or part of an individual's income is derived from any public assistance program. (Not all prohibited bases apply to all programs.) Persons with disabilities who require alternative means for communication of program information (Braille, large print, audiotape, etc.) should contact USDA's TARGET Center at (202) 720-2600 (voice and TDD). To file a complaint of discrimination, write to USDA, Director, Office of Civil Rights, 1400 Independence Avenue, S.W., Washington, D.C. 20250-9410, or call (800) 795-3272 (voice) or (202) 720-6382 (TDD). USDA is an equal opportunity provider and employer.
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.