
Abstract
TLRs, particularly TLR2 and TLR4, play primary roles in inflammatory responses triggered by Aspergillus fumigatus and lead to the activation of signaling pathways that initiate host defense responses. We previously demonstrated that LPS, a ligand of TLR4, can induce tolerance of A. fumigatus hyphae in telomerase-immortalized human stroma fibroblasts (THSFs). In the present study we investigated the role of TLR4, TLR2 and their downstream signaling pathways in this activity. The THSFs were pretreated with low-dose LPS and then exposed to A. fumigatus hyphae. It was demonstrated that enhanced expression of TLR4, but not of TLR2, was associated with LPS pretreatment. Inhibition of TLR4 with monoclonal Ab prevented reduction of pro-inflammatory cytokine secretion in LPS-pretreated THSFs. Pretreatment of THSFs with low-dose LPS caused an impaired response of the MyD88-dependent classical and MAPK signaling pathway upon subsequent A. fumigatus challenge, while expression of signaling molecules in the MyD88-independent Toll-IL-1 receptor domain-containing adaptor inducing IFN-β pathway was increased in THSFs pretreated with LPS. These results indicated that TLR4 mediates attenuated cytokine production induced by LPS pretreatment, and regulation of MyD88-dependent and MyD88-independent pathways may contribute to the development of A. fumigatus hyphae tolerance in LPS-pretreated THSFs.
Introduction
Fungal keratitis is among the most dangerous ocular infections. It can make fast and vicious progress, and often leads to total sight loss in days if not properly controlled.1,2 The best known predisposing factors include trauma (in underdeveloped countries) or contact lens wear (in developed countries). Hot and humid weather is a further risk factor in all countries.3,4 Fusarium spp. and Aspergillus spp. are the most common pathogens of fungal keratitis. 5 Under normal conditions, the cornea is highly resistant to infection, and fungal keratitis always occurs after the epithelial integrity of the cornea has been compromised, exposing underlying fibroblasts. 6 The corneal fibroblasts are the second line of the host innate immunity defense and participate in immediate recognition and control of fungal invasion.
TLRs are key players in innate immunity through their recognition of conserved pathogen-associated molecular patterns in microbes and are expressed at the cell surface or in intracellular space in corneal fibroblasts.7–9 Among these receptors, TLR4 is capable of sensing LPS, while TLR2 recognizes a diverse set of microbial lipopeptides and lipoproteins that are present on Gram-positive and Gram-negative bacteria. 10 Our previous study demonstrated that TLR2 and TLR4 represent two of the master switches initiating innate immunity in the cornea and have determinant roles in Aspergillus fumigatus keratitis.11–15 Their activation leads to the initiation of downstream signal cascades, which regulate secretion of inflammatory cytokines that recruit inflammatory cells to the corneal stroma and the production of antimicrobial molecules that kill the invading pathogens. 16 However, this response of TLRs must be stringently regulated, as excessive inflammation can cause tissue damage and have devastating effects on the host, which can result in vision loss or perforations.
TLRs signal through the adaptor proteins MyD88 and Toll-IL-1 receptor domain-containing adaptor inducing IFN-β (TRIF) to activate downstream transduction cascades. 17 The detrimental effect of TLR signaling is associated with the MyD88-dependent classical pathway and MAPK pathway, which lead to transcription factor NF-κB and activator protein-1 (AP-1) activation, which is essential for the induction of many cytokines and chemokines, such as IL-6, IL-8 and TNF-α.18,19 In contrast, the MyD88-independent TRIF signaling pathway that activates IFN regulatory factor-3 (IRF-3) can induce IFN-β and anti-inflammatory mediators. 20 Thus, in TLR signaling there is a fine balance between pathways leading to injury or protection, with even minor alterations of these fine-tuned endogenous pathways having profound effects on cellular responses to TLR engagement.
We previously reported that telomerase-immortalized human stroma fibroblasts (THSFs) pretreated with LPS develop a state of A. fumigatus hyphae tolerance. Pretreatment of THSFs with low-dose LPS resulted in diminished production of cytokines IL-8 and IL-6, elevated expression of antimicrobial peptides CC chemokine-ligand 20 and thymosin b, and suppression of polymorphonuclear leukocyte migration upon subsequent A. fumigatus challenge. Therefore, LPS pretreatment may induce protective mechanisms during fungal keratitis that prevent an excessive inflammatory response and provide an innate defense in the cornea. 21
The underlying molecular mechanism by which LPS induces tolerance of A. fumigatus hyphae through THSF innate immunity pathways remains to be elucidated. In the present study, the effect of LPS pretreatment on TLR4 and TLR2 mRNA induction was evaluated in order to explore the signaling mechanisms for the resolution of inflammation, restoration of homeostasis and prevention of corneal destruction. Then, these TLRs were inhibited using monoclonal Abs to investigate the role of TLR2 and TLR4 in LPS-induced tolerance using the THSF cell line. In addition, the downstream influence of LPS pretreatment was examined for the purpose of identifying the TLRs signaling pathways involved in THSFs, including the MyD88-dependent classical pathway, MyD88-dependent MAPK pathway and MyD88-independent TRIF signaling pathway.
Materials and methods
A. fumigatus strain and preparation of hyphae
The A. fumigatus strain CCTCC 93024, purchased from the China Centre for Type Culture Collection, was grown on Sabouraud Glc agar for 24 h at 37℃ on a shaking table with a rotation speed of 200 rpm. Conidia were harvested and planted into Sabouraud fluid media, at a final concentration of 108 microorganisms/ml. The tubes were shaken for 18 h at 26℃ at 500 rpm and then centrifuged at 1000 g for 10 min. The mycelia were washed twice, suspended in PBS and sterilized by heat treatment at 56℃ for 60 min. Then, the mycelia were disrupted into 20–40 mm pieces in a Potter-Elvehjem Tissue Grinder (Wheaton Industries Inc., Millville, NJ, USA), which yielded 5 × 106 pieces/ml as A. fumigatus hyphae Ag.
Cell cultures
THSFs (kindly provided by Dr Fu-Shin X. Yu and produced by Dr Ilene K. Gipson) were cultured in DMEM (Invitrogen Life Technologies, Carlsbad, CA, USA) supplemented with 10% FBS (Gibco, Carlsbad, CA, USA) at 37℃ in a humidified atmosphere with 5% CO2. For experiments, cells were seeded at 2 × 105 cells/well in six-well microculture plates and grown until 80% confluence (∼ 2 d). To prepare cells for stimulation, the THSFs were then starved in serum-free DMEM for 16 h.
Cell treatment and tolerance induction
The THSFs were preconditioned with a TLR4 ligand, LPS (protein content < 3%; Escherichia coli, serotype O55:B5; Sigma Chemical, St Louis, MO, USA) at a density of 10 ng/ml for 12 h. Then the cells were washed twice with serum-free medium and re-stimulated with A. fumigatus hyphae (106 pieces/ml) for various periods (30 min, 1 h, 3 h, 6 h, 12 h). The cell culture supernatants and cells were collected for measurement of mRNA and protein expression.
TLR blocking experiments were conducted by incubating THSFs with monoclonal Abs against TLR2 or TLR4. THSF cells were incubated with anti-human TLR2 (100 µg/ml) or anti-human TLR4 (100 µg/ml), or both anti-human TLR2 (100 µg/ml) and anti-human TLR4 (100 µg/ml) monoclonal Abs (sc-47728 L and sc-13593 L respectively; Santa Cruz Biotechnology, Santa Cruz, CA, USA) for 30 min. They were then challenged with TLR2 ligand zymosan (Saccharomyces cerevisiae; Sigma Chemical) at a density of 1 mg/ml or TLR4 ligand LPS (1 µg/ml) for 12 h at 37℃, or pretreated with LPS for 12 h and re-challenged with A. fumigatus hyphae for 4 h. The cell culture media and cell lysates were harvested and used for IL-6, IL-8 and TNF-α protein ELISA and TLR2 mRNA real-time RT-PCR.
Quantitative real-time RT-PCR
Total RNA was extracted from THSFs with TRIzol reagent (Invitrogen), and cDNA was synthesized with a First Strand cDNA Synthesis Kit (Toyobo, Osaka, Japan) according to the manufacturer’s protocol. Quantitative RT-PCR was performed using SYBR Green Real time PCR Master Mix (Toyobo) with 2 µl of respective cDNA and 0.4 µm of each oligonucleotide primer in a total volume 20 ml. The reaction was performed on the ABI 7000 Sequence Detection System (Applied Biosystem, Foster City, CA, USA). The real-time primers used were as follows: TLR4, forward 5′-GATTGCTCAGACCTGGCAGTT-3′ and reverse 5′-TGTCCTCCCACTCCAGGTAAGT-3′; TLR2, forward 5′-CTGCAAGCTGCGGAAGATAAT-3′ and reverse 5′-AGGACTTTATCGCAGCTCTCAGA-3′; MyD88, forward 5′-CTGCTCGAGCTGCTTACCA-3′ and reverse 5′-CTTTTGGCAATCCTCCTCAA-3′; IκB-α, forward 5′-AAGTGATCCGCCAGGTGAAG-3′ and reverse 5′-GCAATTTCTGGCTGGTTGGT-3′; MAPK3, forward 5'-TCAACACCACCTGCGACCTT-3' and reverse 5'-GCGTAGCCACATACTCCGTCA-3; AP-1, forward 5'-GGATAGCCTCTCTTACTACCAC-3' and reverse 5'-TCCTGTCATGGTCTTCACAACG-3'; TRIF, forward 5′-CCAGATGCAACCTCCACTGG-3′ and reverse 5′-CTGTTCCGATGATGATTCC-3′; IRF-3, forward 5′-GAACCCCAAAGCCACGGATC-3′ and reverse 5′-CCTCCCGGGAACATATGCAC-3′; IFN-β, forward 5′-GCTTGGATTCCTACAAAGAAGCAGC-3′ and reverse 5′-CAAAGTTCATCCTGTCCTTGAGGC-3′; GAPDH, forward 5′-TGAACGGGAAGCTCACTGG-3′ and reverse 5′-TCCACCACCCTGTTGCTGTA-3. The real-time RT-PCR protocol was as follows: one cycle at 94℃ for 1 min, followed by 40 cycles at 95℃ for 15 s, then at an annealing temperature of 56℃ (TLR4, MyD88, IκB-α, IRF-3, GAPDH) or 60℃ (TLR2, MAPK3, AP-1, TRIF, IFN-β) for 15 s and, eventually, at 72℃ for 1 min. The mRNA level was normalized to the gene expression of GAPDH in each sample.
ELISA detection of cytokines
Concentrations of IL-6, IL-8, TNF-α and IFN-β protein were measured in the supernatants by ELISA, following the manufacturer’s protocol (KYM, Beijing, China). In this study the sensitivity range of the IL-6, IL-8, TNF-α and IFN-β assay were 0–2000, 0–2000, 15–1000 and 0–100 pg/ml, respectively.
Western blot analysis
THSFs were collected at the indicated times and lysed with RIPA buffer (50 mM Tris–HCl, 1% NP-40, 0.25% Na-deoxycholate, 150 mM NaCl, 1 mM Na3VO4 and NaF). Supernatants containing total cell lysate proteins were obtained by centrifugation at 12,000 g for 15 min and the protein concentration was determined using BCA Protein Assay Kit from Pierce Biotechnology (Rockford, IL, USA). The supernatant was boiled for 5 min in sample buffer, and 50 mg protein samples were electrophoresed on 10% SDS-PAGE. The contents of the gel were then transferred to a polyvinylidene fluoride membrane and the membrane was blocked with 5% skim milk for 2 h at room temperature (20–25℃). The membrane was incubated overnight (15–18 h) with primary Abs (Abcam, Cambridge, MA, USA) diluted 1:1000 at 4℃. Protein blots were then probed with the corresponding HRP-conjugated secondary Abs (Amersham; Buckinghamshire, UK) at room temperature for 1 h. The immunoreactive band was visualized using an enhanced chemiluminescent substrate. To perform densitometry analysis, digital images of the positive bands were analyzed using the image analysis program Quantity One (Bio-Rad, Hercules, CA, USA).
Immunofluorescence staining
The slides seeded with THSFs were fixed and permeabilized with cold acetone for 10 min, washed with PBS three times for 5 min and incubated for 30 min in blocking buffer of 5% goat serum at room temperature. Slides were incubated with anti-NFκB-p65 monoclonal Ab (Abcam) diluted 1:50 overnight at 4℃. They were then incubated at room temperature for 1 h with the diluted secondary Abs (1:100) conjugated to Cy3 or FITC (KPL, Gaithersburg, MD, USA), and 4′,6-diamidino-2-phenylindole dihydrochloride (; Roche Diagnostics, Mannheim, Germany) was used for nuclear counterstaining. The negative control was similarly treated, but the primary Ab was omitted. The staining was viewed and photographed with fluorescence microscopy (Olympus fluorescence convert microscopeX81; Olympus Optical, Tokyo, Japan).
Statistical analysis
The statistical significance of data was evaluated by a two-tailed Student’s t-test for unpaired data or by one-way ANOVA for multiple comparisons. A P-value < 0.05 was considered significant. All data of at least three independent experiments were expressed as means ± SEM.
Results
Efficient neutralization of TLR2 or TLR4 by monoclonal Abs
To determine the involvement of TLR2 and TLR4 in the inflammatory cytokine induction in LPS-pretreated THSFs, we used monoclonal Abs specific for TLR2 or TLR4 to neutralize their function. As shown in Figure 1, TLR2 or TLR4 Ab had no apparent effect on the basal release of IL-8 and TNF-α in THSFs. While THSFs secreted significantly high levels of IL-8 and TNF-α in response to TLR2 ligand zymosan or TLR4 ligand LPS challenge, this elevated production was significantly inhibited in cells treated with TLR2 or TLR4 Ab, and there was no significant difference between blank control and the ligand-treated THSFs with previous TLRs Abs inhibition. Thus, our monoclonal Abs specific for TLR2 and TLR4 can efficiently neutralize the function of TLR2 and TLR4, respectively.
Neutralizing capacity of TLR2 and TLR4 Abs. THSFs were incubated with 100 µg/ml anti-human TLR2 (A) or 100 µg/ml anti-human TLR4 (B) mAbs for 30 min, and then challenged with or without 1 mg/ml TLR2 ligand zymosan (A) or 1 µg/ml TLR4 ligand LPS (B) for 12 h. Data shown are representative of triplicate experiments for each different treatments. All values were expressed as mean ± SEM. One-way ANOVA: **P < 0.01, #P > 0.05.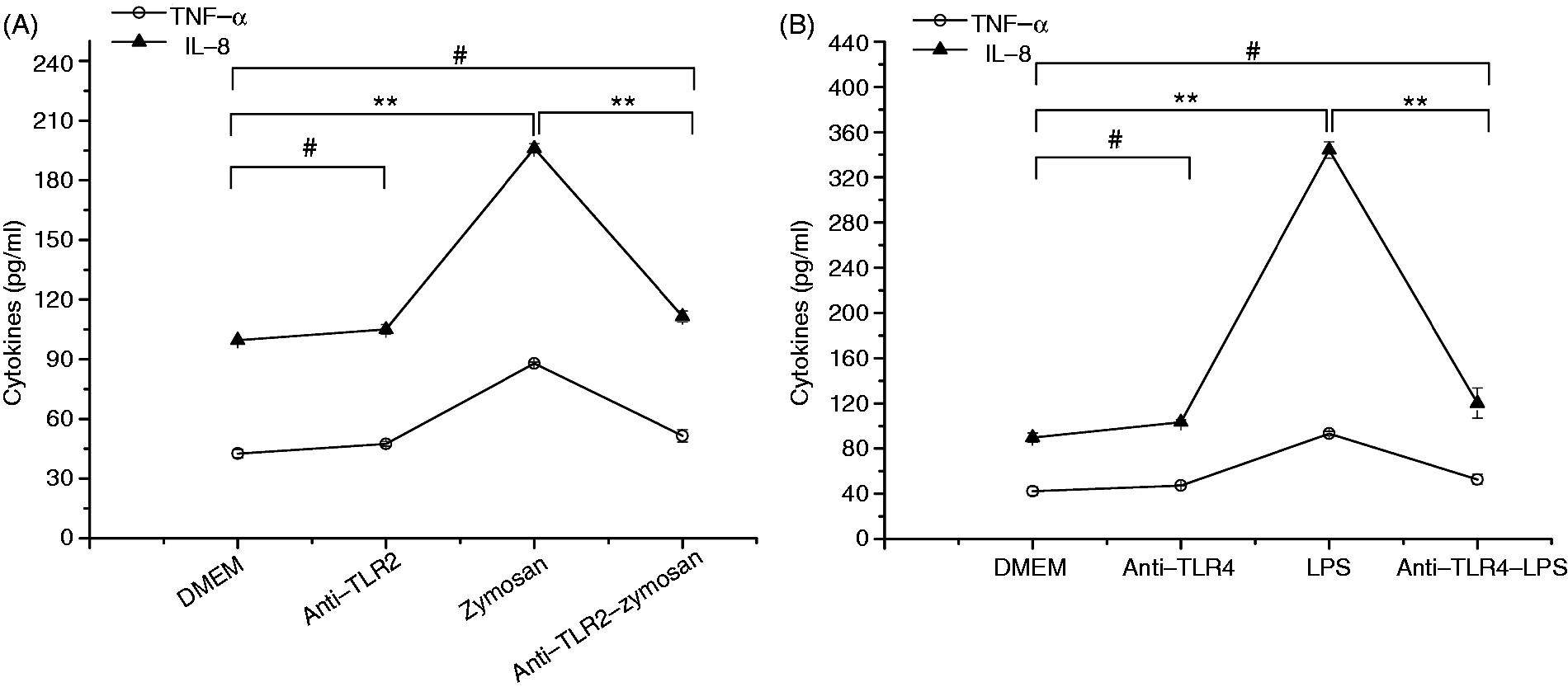
Effect of LPS pretreatment on TLR4 expression induced by A. fumigatus in THSFs
To examine whether TLR4 expression is affected by LPS pretreatment in THSFs, we pre-stimulated THSFs with 10 ng/ml LPS for 12 h and assessed the response of TLR4 expression to subsequent challenge with A. fumigatus hyphae. Real-time RT-PCR revealed that LPS pretreatment significantly down-regulated A. fumigatus hyphae-induced mRNA expression of TLR4 after 1 h and 3 h of re-stimulation (Figure 2A).
Role of TLR4 and TLR2 in A. fumigatus (AF) hyphae tolerance induced by LPS. (A) Aspergillus fumigatus-stimulated TLR4 mRNA expression in LPS-pretreated THSFs. After pretreatment with DMEM or 10 ng/ml LPS for 12 h, the THSFs were re-stimulated with A. fumigatus hyphae for various periods (30 min, 1 h, 3 h, 6 h). Real-time RT-PCR was performed to assess mRNA levels. Data represent mean ± SEM of three independent experiments. Student’s t-test: *P < 0.05 compared with A. fumigatus stimulation without LPS pretreatment. (B) Pro-inflammatory cytokines secretion in THSFs. The THSFs were pre-incubated with anti-human TLR4 (100 µg/ml) or both anti-human TLR2 (100 µg/ml) and anti-human TLR4 (100 µg/ml) mAbs for 30 min, and then cultured with or without 10 ng/ml LPS for 12 h. After being washed twice with serum-free medium, the cells were stimulated with A. fumigatus hyphae for 4 h. IL-6, IL-8 and TNF-α secretion into culture supernatants was assayed by ELISA. Data are representative of three repeated experiments. All values were expressed as mean ± SEM. One-way ANOVA: *P < 0.05, **P < 0.01, #P > 0.05. (C) Effect of LPS pretreatment on mRNA expression of TLR2. The THSFs were pre-incubated with anti-human TLR4 (100 µg/ml) mAbs for 30 min and then cultured with or without 10 ng/ml LPS for 12 h. After being washed twice with serum-free medium, the cells were stimulated with A. fumigatus hyphae for 4 h. Bars represented mean ± SEM of three independent experiments. Student’s t-test: *P < 0.05, **P < 0.01, #P > 0.05.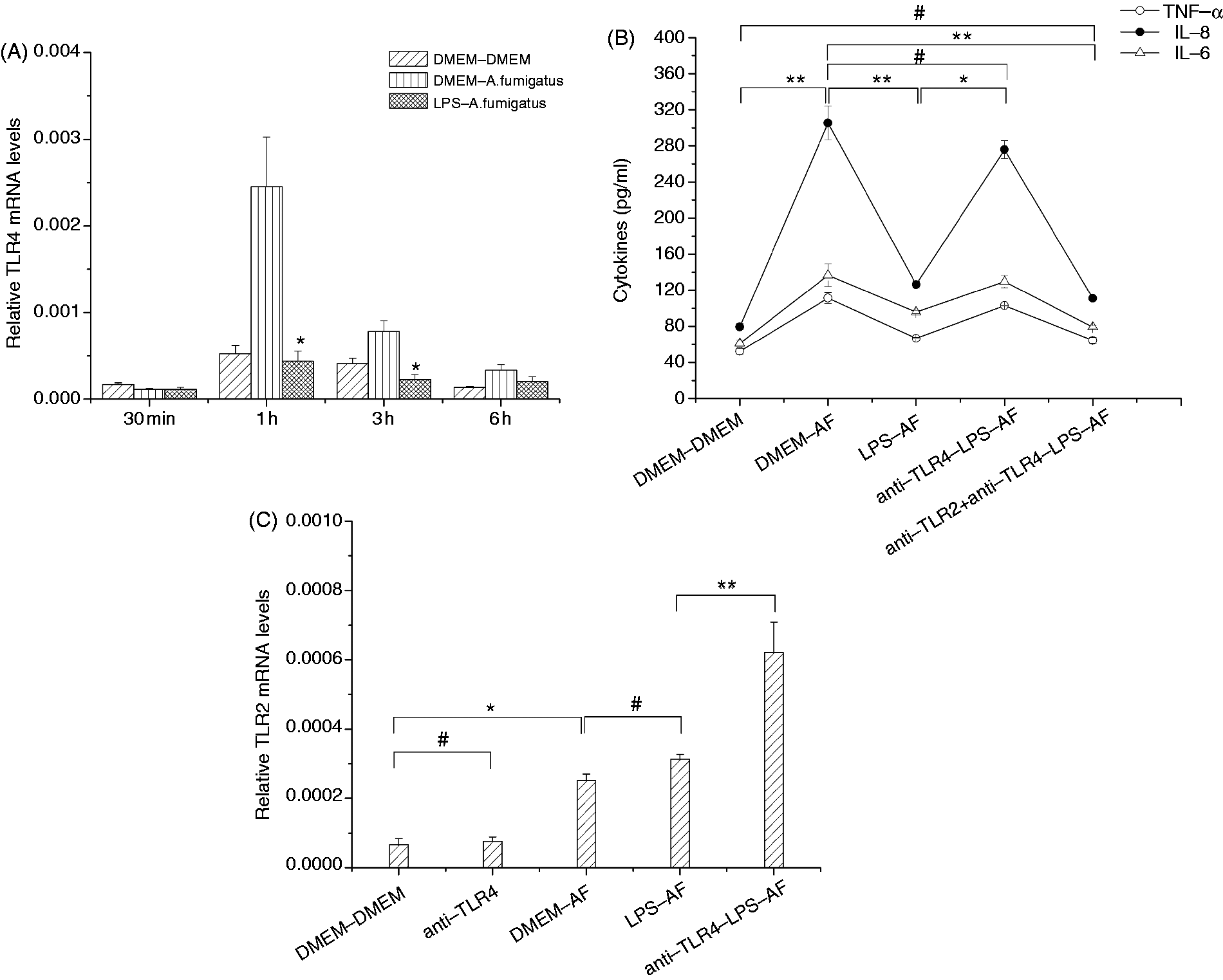
LPS pretreatment induced down-regulation of pro-inflammatory cytokines secretion in A. fumigatus Ag-challenged THSFs depends on TLR4
In our previous study, we demonstrated that pretreatment of THSFs with 10 ng/ml LPS resulted in impaired production of IL-6 and IL-8 in response to a secondary A. fumigatus hyphae challenge. We next sought to determine whether the reduction of pro-inflammatory cytokines secretion induced by LPS pretreatment in THSFs is dependent on TLR4. THSFs pre-incubated with TLR4 Abs before LPS pretreatment showed increased IL-6, IL-8 and TNF-α accumulation in culture medium compared with LPS-pretreated THSFs without the TLR4 Ab, and there was no significant difference of IL-6, IL-8 and TNF-α accumulation in culture medium compared to non-LPS-pretreated controls challenged with A. fumigatus without TLR4 Ab inhibition (Figure 2B).
Involvement of TLR2 in LPS-induced tolerance to secondary A. fumigatus hyphae stimulation
Real-time PCR was used to assess expression of TLR2. It showed that LPS pretreatment had no apparent effect on TLR2 expression in THSFs challenged with A. fumigatus hyphae. However, whereas the TLR4 Ab had no effect on basal TLR2 mRNA expression, THSFs treated with TLR4 neutralizing Ab before LPS pretreatment exhibited significantly increased levels of TLR2 after the secondary fungi stimulation compared with the pretreated controls. (Figure 2C). In addition, pre-incubation of THSFs with both TLR2 and TLR4 Abs before LPS pre-stimulation resulted in dramatic reduction of the pro-inflammatory cytokines, including IL-6, IL-8 and TNF-α expression compared with the non-pretreated THSFs (Figure 2B).
Effect of LPS pretreatment on the expression of downstream molecules in TLR signaling pathways in THSFs stimulated by A. fumigatus
To further explore the underlying mechanism by which pretreatment with LPS modulates innate immunity in corneal fibroblasts challenged with A. fumigatus, THSFs were pretreated with 10 ng/ml of LPS for 12 h and the cell responses of multiple downstream molecules in TLRs signaling pathways to subsequent stimulation with A. fumigatus hyphae were investigated in this study of gene and protein levels.
Down-regulated expression of downstream molecules in MyD88-dependent classical signaling pathway
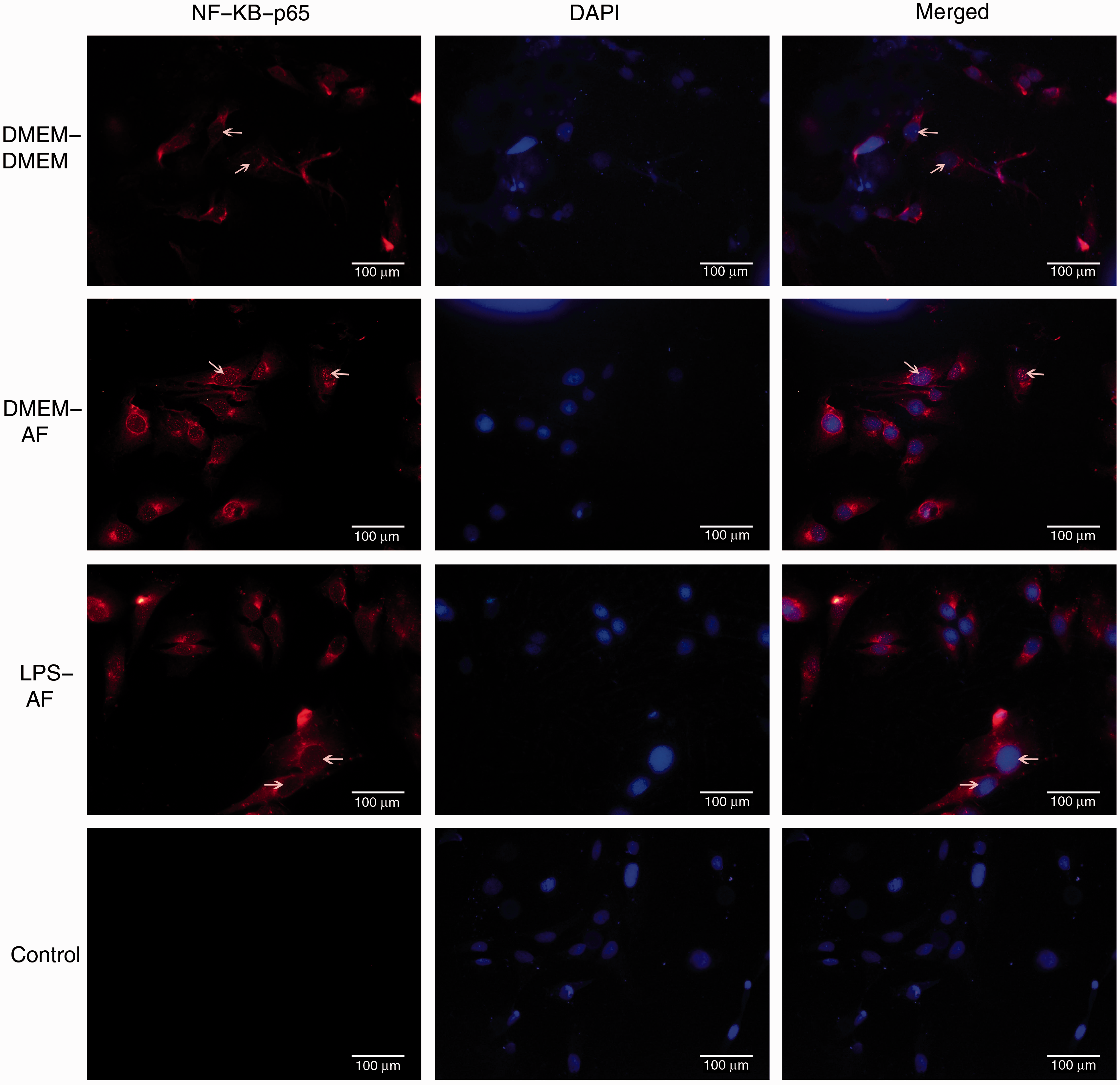
mRNA expression of MyD88 was proven to be inhibited after 1 h and 3 h of A. fumigatus re-stimulation according to the PCR analysis (Figure 3A), and an increase in gene (Figure 3B) and protein (Figure 3C) expression of inhibitory NF-κB (IκB)-α was observed 1 h and 3 h after fungus treatment. NF-κB-p65 protein expression was diminished after 1 h and 3 h of stimulation with A. fumigatus hyphae (Figure 3C), and the translocation of NF-κB-p65 into cell nuclei was significantly attenuated by LPS pretreatment (Figure 4).
Effect of LPS pretreatment on expression of downstream molecules in MyD88-dependent classical signaling pathway. The THSFs were pretreated with or without 10 ng/ml of LPS for 12 h before being challenged with A. fumigatus (AF) hyphae for various periods (30 min, 1 h, 3 h, 6 h). The cells were assayed by real-time RT-PCR to assess mRNA expression of MyD88 (A) and IκB-α (B). Cell lysates were prepared, and Western blotting was performed to determine levels of IκB-α and NF-κB-p65(C). Data from PCR are representative of three experiments and are presented as mean ± SEM. Student’s t-test: *P < 0.05; **P < 0.01 compared with A. fumigatus stimulation without LPS pretreatment. Immunofluorescence analysis of NF-κB-p65 expression and position. The THSFs were pretreated with DMEM or 10 ng/ml LPS for 12 h before 2 h re-stimulation with A. fumigatus (AF) hyphae (white arrows indicate positive staining of NF-κB-p65 in cell nuclei). Sections without primary Ab incubation were used as controls and showed no positive staining. DAPI counterstaining was used to visualize cell morphology. Original magnification: 400×.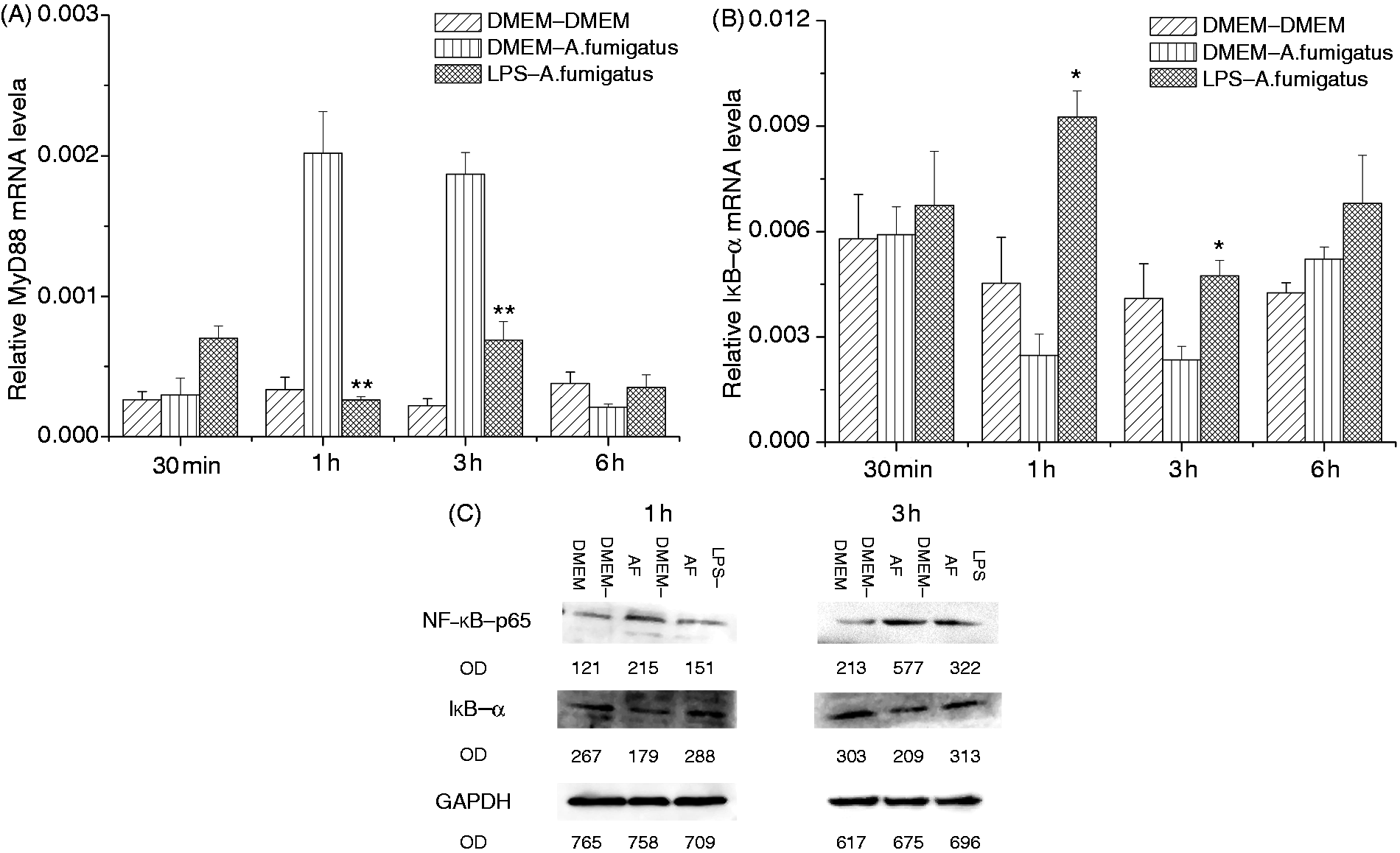
Impaired response of MyD88-dependent MAPK signaling pathway
Expression of MAPK3 (Figure 5A) and AP-1 (Figure 5B) mRNA was lowered by LPS pretreatment 1 h and 3 h after the secondary A. fumigatus stimulation, and Western blot analysis showed that protein level of phosphorylated extracellular signal-regulated kinase (ERK) 1/2 was decreased after 1 and 3 h of treatment with A. fumigatus hyphae (Figure 5C).
RT-PCR and Western blot assay of downstream molecules expression in MyD88-dependent MAPK signaling pathway. The THSFs were pretreated with DMEM or 10 ng/ml LPS for 12 h before re-stimulation with A. fumigatus (AF) hyphae for various periods (30 min, 1 h, 3 h, 6 h). (A, B) Real time RT-PCR for quantization of MAPK3 and AP-1 mRNA in THSFs.(C) Western blot analysis of p-Erk1/2 expression. Results are expressed as the mean ± SEM of three samples. Student’s t-test: *P < 0.05; **P < 0.01 compared with A. fumigatus stimulation without LPS pretreatment.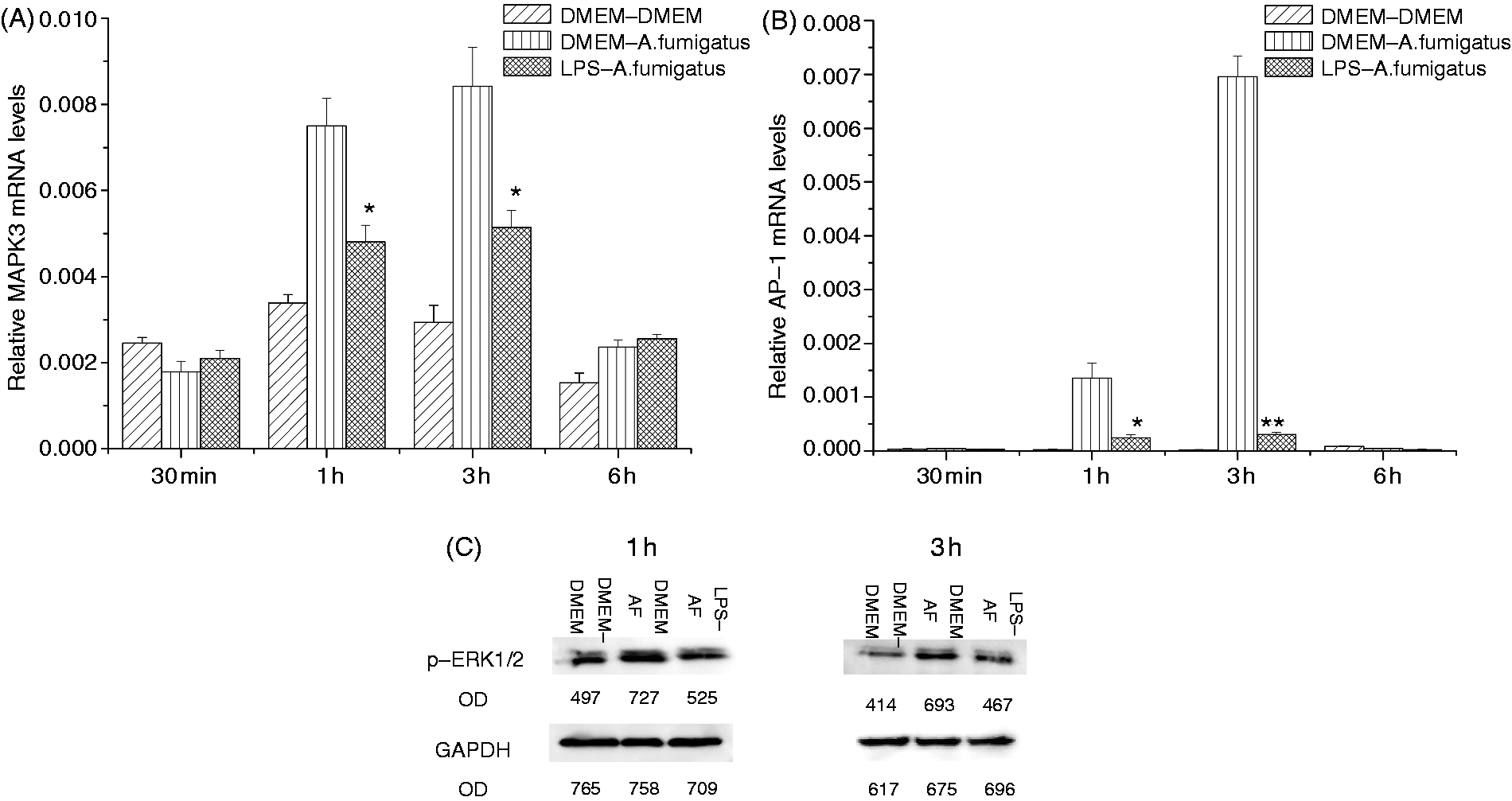
Up-regulation of downstream molecular expression and cytokine secretion of the MyD88-independent TRIF signaling pathway
THSFs pretreated with LPS were challenged with A. fumigatus hyphae, and TRIF, IRF3 and IFN-β expression were measured by real-time RT-PCR and ELISA. As shown in Figure 5, in non-pretreated controls, THSFs express relatively high levels of TRIF, IRF3 and IFN-β mRNA in response to A. fumigatus hyphae challenge for 3 and 6 h, and this elevated expression was significantly increased in cells pretreated with LPS. IFN-β secretion levels were consistent with trends observed for IFN-β mRNA (Figure 6D).
The MyD88-independent TRIF signaling pathway in LPS-induced tolerance to A. fumigatus hyphae in THSFs. The THSFs were pretreated with DMEM or 10 ng/ml LPS for 12 h and re-challenged with A. fumigatus hyphae for various periods (30 min, 1 h, 3 h, 6 h, 12 h). Total RNA was isolated from cells to assess mRNA expression of TRIF (A), IRF3 (B) and IFN-β (C). The culture supernatants were used for IFN-β ELISA (D). Results shown are means ± SEMs of three independent experiments. Student’s t-test: *P < 0.05 compared with untreated controls; #P < 0.05 compared with A. fumigatus stimulation without LPS pretreatment.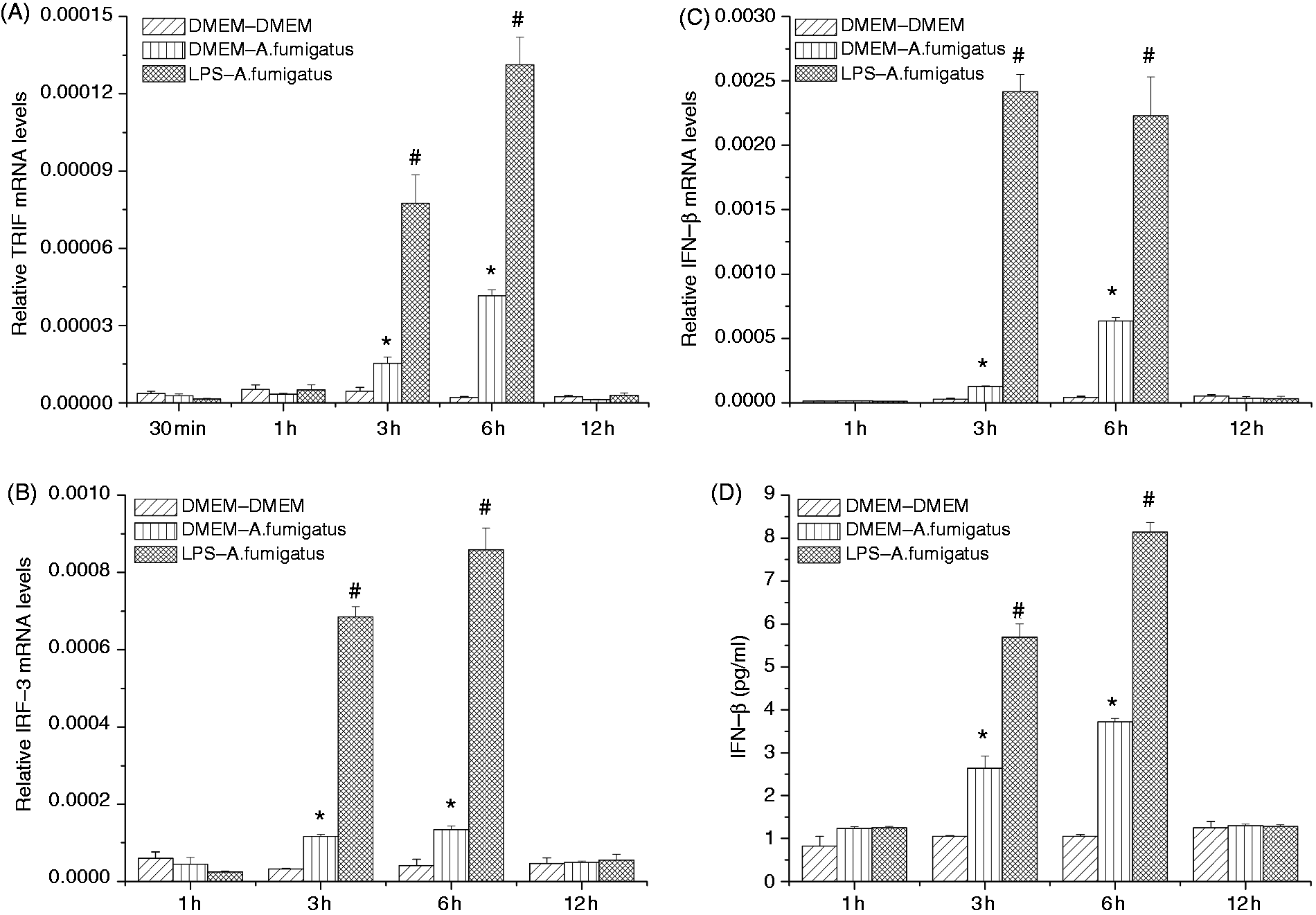
Discussion
In the present study, it was demonstrated that pre-exposure of THSFs to LPS led to decreased expression of TLR4 in response to re-stimulation with A. fumigatus hyphae, but failed to modulate TLR2 expression. TLR2 and TLR4 mAbs treatment is effective in the inhibition of TLR2 and TLR4 function, while the secretion of basal inflammatory cytokines was not affected by TLRs Abs. The repression of pro-inflammatory cytokines (TNF-α, IL-6 and IL-8) secretion caused by LPS pretreatment can be reversed when TLR4 is previously inhibited with a mAb in LPS-preconditioned THSFs. In addition, although the function of the MyD88-dependent classical pathway and MyD88-dependent MAPK pathway was dampened, expression of key factors and cytokine in the MyD88-independent TRIF signaling pathway was augmented.
Previous studies from our laboratory suggested that TLR4 is one of the major receptors in the cornea for recognition of hyphae and supernatant Abs from A. fumigatus. Release of IL-1β, IL-6 and IL-10 in corneal epithelium triggered by A. fumigatus Abs could be inhibited by treatment with the TLR4 Ab or TLR4-siRNA plasmid.9,11–14 Conflicting results have been published regarding the influence of LPS pretreatment on TLR4 expression level. It has been reported that LPS pretreatment did not modulate expression of TLR4 on the surface of human peripheral blood mononuclear cells, 22 macrophages, 23 liver sinusoidal endothelial cells, 24 intestinal microvascular endothelial cells, 25 mouse murine mast cells 26 and preterm fetal sheep lung and liver 27 in endotoxin tolerance, a phenomenon in which pre-exposure to low doses of LPS induces resistance to a lethal challenge. However, other studies showed that TLR4 expression was markedly reduced in LPS-pretreated dendritic cells, 28 monocytes, 29 human monocytic cell line THP-1, 30 mouse peritoneal macrophages 31 and spleen 32 upon secondary stimulation. Patenaude et al. 33 demonstrated that LPS pretreatment significantly decreased the percentage of CD11c+ Fms-like tyrosine kinase-3 ligand-dependent dendritic cells (FLDCs) that express TLR4. In the present study, it was shown that expression of TLR4 was decreased in LPS-pretreated THSFs compared with medium-pretreated cells after A. fumigatus hyphae challenge, and the dramatic decrease of IL-6, IL-8 and TNF-α secretion in LPS-pretreated THSFs was abrogated by blockade of TLR4 with a mAb. Thus, TLR4 appears to be necessary and sufficient for the development of A. fumigatus hyphae tolerance in LPS-pretreated THSFs and may play a gatekeeper’s role in regulating subsequent immune responses.
To explain the maintenance of the high level of pro-inflammatory cytokines (IL-6, IL-8 and TNF-α) after TLR4 inhibition in LPS-pretreated THSFs, we investigated the role of TLR2, another key receptor for A. fumigatus, in the A. fumigatus hyphae tolerance in LPS-pretreated THSFs. TLR2 recognizes lipoproteins from bacteria and mycobacteria, as well as zymosan from fungi. 10 It has been proven that TLR2 in corneal epithelium and fibroblasts had an important role in the pathogenesis of fungal keratitis, and TLR2 siRNA treatment attenuated A. fumigatus keratitis in rats by suppressing corneal inflammation and preventing fungal invasion.11–15 In contrast to the effects of LPS pretreatment on TLR4 expression, in the present study it was found that levels of TLR2 mRNA were not affected by LPS pretreatment. In agreement with these findings, Piao et al. 32 demonstrated that a bolus injection of 10 mg/kg LPS for mice pretreated with 0.5 mg/kg/d of LPS for 14 d caused no alteration of TLR2 protein expression in the spleen. Interestingly, we observed that although TLR4 Ab had no significant effect on basal TLR2 mRNA expression, TLR2 was significantly up-regulated in the LPS-preconditioned THSFs following inhibition of TLR4. A compensatory adjustment for TLR4 inhibition in LPS-pretreated THSFs challenged by A. fumigatus hyphae may be partially responsible for this curious phenomenon, and the elaborate and complicated feedback network that involves various regulators of TLR signaling may also play a role.34,35 Moreover, it was reported that the secretion of inflammatory cytokines by murine macrophage and human blood leukocytes previously exposed to LPS is markedly reduced when re-challenged with ligands for TLR2, indicating that TLR2 can be functionally affected after in vitro or in vivo exposure to LPS.36,37 Thus, a third possible mechanism is that after TLR4 inhibition, LPS pretreatment somehow exerts certain influence on TLR2 expression in THSFs. Although our data do not distinguish among these possibilities, it is clear that TLR4 and TLR2 may not work in isolation, but instead cooperate to fine-tune their specificity and to regulate the magnitude of cellular responses. The dual inhibition of TLR4 and TLR2 can bring secretion of IL-6, IL-8 and TNF-α to basal levels, which reinforce the idea that after TLR4 blockage, inflammatory cytokine secretion is mediated by TLR2 in LPS-pretreated THSFs.
On recognizing PAMPS, the TLRs signal through intracellular signal cascades and initiate gene expression of downstream molecules, which lead to transcription factor activation and the generation of cytokines and chemokines. 16 Each TLR family member, with the exception of TLR3, recruits the adapter molecule MyD88 by homotypic interaction with a Toll-IL 1 receptor domain in its terminus, which are MyD88-dependent pathways. In turn, this activity leads to the activation of transcription factor NF-κB in the MyD88-dependent classical signaling pathway. Members of the MAPK family, including MAPK3, c-Jun NH2-terminal kinase (JNK), p38 and ERK1/2, phosphorylate and activate components of the transcription factor AP-1 in the MyD88-dependent MAPK signaling pathway. Together, these transcription factors induce expression of many inflammatory cytokines such as IL-6, IL-8 and TNF-α.17–20 Decreased TLR4–MyD88 complex formation was observed in endotoxin-tolerant human monocytes. 38 LPS tolerance in experimental human endotoxemia and Gram-negative sepsis appeared to be associated with elevated levels of IL-1 R-associated kinase M, an intracellular negative regulator of MyD88-dependent TLR signaling. 36 Decreased phosphorylation and impaired signal transduction of ERK1/2, p38 and JNK was noted after LPS re-stimulation in FLDCs, 33 human peripheral blood mononuclear cells, 22 human monocytic cell line THP-123,30 and intestinal microvascular endothelial cells 25 preconditioned with LPS. Pretreatment of mouse macrophages with LPS inhibited activation of the transcription factors AP-1 in response to subsequent LPS stimulation. 39 The present study demonstrated that pretreatment of THSFs with LPS induced decreased expression of MyD88, MAPK3, phosphorylated-ERK1/2 (p-ERK1/2) and the transcription factor AP-1 upon subsequent challenge with A. fumigatus hyphae, suggesting that the impaired activity of the MyD88-dependent classical pathway and MyD88-dependent MAPK pathway may contribute to the suppressed secretion of pro-inflammatory cytokines.
Although the recognition of different ligands by specific TLRs leads to activation of an intracellular signaling cascade in an MyD88-dependent or MyD88-independent fashion, all TLRs share NF-κB signal transduction pathways for activation of the transcription factors. NF-κB is critical for maximal expression of many cytokines involved in the pathogenesis of inflammation. Activation and regulation of NF-κB are tightly controlled by a group of inhibitory proteins (IκB), which maintain NF-κB in the cytoplasm of effector cells. Phosphorylation and degradation of IκBs are necessary for the subsequent nuclear translocation of the transcription factor NF-κB and migration of NF-κB to the nucleus is a critical step in the coupling of extracellular stimuli to the transcriptional activation of specific target genes.16–20 Our previous study showed that the main pro-inflammatory cytokines, including IL-6, IL-8, IL-1β and IL-10 and TNF-α, were secreted in a TLR–NF-κB-dependent manner in human corneas challenged by A. fumigates.14,40 LPS tolerance was associated with an impaired activation of NF-κB, inhibition of NF-κB DNA binding, and depletion of both p65 and p50 forms in macrophages.31,37,39 Decreased activity of NF-κB-p65 and increased presence of NF-κB inhibitors, including Ship1, Tollip and p105, in LPS-preconditioned mice following stroke was observed by Vartanian et al. 41 Endotoxin tolerance in FLDCs was associated with a decrease in NF-κB translocation to the nucleus. 33 LPS preconditioning suppresses TLR4-inducible IκB-α degradation in endotoxin-tolerant human monocytes. 30 In our study, consistent with increased IκB-α level, we found that expression of NF-κB-p65 was decreased in LPS-pretreated THSFs upon subsequent A. fumigatus hyphae stimulation. Our results have also shown that LPS pretreatment induced a decrease in NF-κB-p65 translocation to the nucleus.
TLR4 is a unique member of the TLR family in that it signals through both MyD88-dependent and MyD88-independent pathways. The latter pathway leads to the activation of transcription factor IRF-3 and, ultimately, induces the production of IFN-β, one of the so-called type I IFNs. IFNs are natural cell-signaling proteins in response to viral, bacterial and parasitic infections.16–20 Despite the increasing incidence of fungal infections and their associated mortality, the presence and possible role of the MyD88-independent TRIF pathway and IFN-β in defense against these infections has received little attention. Evidence indicates that LPS preconditioning resulted in a marked up-regulation of anti-inflammatory/type I IFN-associated genes, and IRF3 activity was enhanced in LPS-preconditioned mice following stroke. 41 de Vos et al. 36 observed that intravenous injection of LPS in humans induces reduced levels of the recently identified TRIF inhibitor selective androgen receptor modulator in circulating leukocytes. Here we have demonstrated that IFN-β secretion is robustly up-regulated in the LPS-preconditioned THSFs in response to A. fumigatus and that several signaling factors are also up-regulated, including TRIF and IRF3, suggesting that increased signaling via the TLR4–IRF3 axis may mitigate the damaging effects of fungi. This is supported by the results of Gafa et al., 42 who proposed a possible effect of IFN-β on anti-Aspergillus immune responses in human monocyte-derived dendritic cells infected in vitro with A. fumigatus. In addition, the response of TRIF to the stimulation was found later than that of MyD88 in our study, indicating that LPS pretreatment may attenuate the inflammatory response of THSFs to A. fumigatus hyphae at the early stage, whereas TRIF-mediated immunological protection was augmented in relatively late-phase TLR signaling.
Finally, another interesting phenomenon is revealed in our study: LPS pretreatment exerted a relatively larger effect on the signaling factors, such as TLR4, MyD88 and AP-1, than on the secreted cytokines (IL-6, IL-8, TNF-α and IFN-β). First of all, TLR activation is strictly and finely regulated at multiple levels of the signal transduction pathways by various mediators and other innate receptors, and additional components and regulation of known components continue to be uncovered.34,35 Thus, the synthesis and secretion of cytokines in LPS-pretreated THSFs may also be specially regulated to dampen an excessive inflammatory response and, at the same time, control infection effectively. Also, most of the intermediate signaling factors, such as TLR4, MyD88 and AP-1, were evaluated at the genetic level, while the secreted protein levels of cytokines were employed in our study, hence the complex process of protein translation and post-translational modifications may also play a role. 43 Another possibility may lie in the fact that the levels of the secreted portion in the cell culture supernatant, which is also the effective part in the immune response, rather than the total production of cytokines, were estimated in this study.
In summary, we suggest that pretreatment with LPS reprograms the TLR4′s response to A. fumigatus in THSFs, resulting in alterations to TLR4 intracellular signaling by suppression of the MyD88-dependent pathways and up-regulation of the MyD88-independent TRIF signaling pathway. Reprogramming causes a finely controlled shift in the balance of TLR4-mediated pro-inflammatory and anti-inflammatory responses, and represents an endogenously orchestrated mechanism that may be sufficient to defend against fungal pathogens and, at the same time, avoid corneal bystander damage caused by excessive inflammation. These novel findings shed light on the understanding of innate corneal fibroblasts immunity involved in fungal inflammation, and might create new therapeutic targets and intervention strategies to control fungal keratitis.
Footnotes
Funding
This work was supported by the National Natural Science Foundation of China [grant number 81070707].
Acknowledgements
We are grateful to Dr Edward C. Mignot, Shandong University, for linguistic advice.