
Abstract
Lymphotoxin beta-receptor (LTβR) is involved in the formation and maintenance of secondary lymphoid structures, as well as in the regulation of inflammatory responses. Because LTβR lymphoid structure formation continues to develop in infants, we compared two different chimera models: one using adult mice and the other using a transplantation model of neonatal mice. To elucidate the function of LTβR on lymphoid and non-lymphoid cells, we generated bone marrow chimeras on the wild type C57Bl/6 and the LTβR-deficient (LTβR−/−) background, and reconstituted the mice with bone marrow cells reciprocally. These chimeric mice were analyzed in the experimental model of acute dextran sulfate sodium-induced colitis. Interestingly, both models revealed not only equal reconstitution levels but also similar immunological responses: LTβR expression on stromal cells is essential for lymph node formation, whereas LTBR on hematopoietic cells is crucial for a decrease in inflammation. In addition, mice lacking LTβR on hematopoietic cells revealed (a) an increase of immature granulocytic cells in the spleen and (b) a reduced proportion of myeloid cells in peripheral blood and spleen expressing CD11b+Ly6C+Ly6G− (myeloid-derived suppressor cells expression profile). In conclusion, LTβR expression on hematopoietic cells seems to be involved in the down-regulation of acute inflammatory reactions paralleled by the appearance of immature myeloid cells.
Introduction
Lymphotoxin beta-receptor (LTβR) signaling is involved in lymphoid organogenesis and the maintenance of secondary lymphoid structures,1–3 as well as the regulation of acute inflammatory reactions. 1 Recent studies described the importance of LTβR during dextran sulfate sodium (DSS)-induced colitis by inhibition of LTβR activation.4–6 Blocking of LTβR signaling was achieved either by treatment with LTβR–Ig, a functional inhibitor of LTβR signaling, or by congenital deletion of the LTβR or its functional ligand lymphotoxin (LT) α1β2.
LTβR also seems to be involved in the recruitment of different hematopoietic cells in lymphoid and non-lymphoid compartments, including the recruitment of B cells into the intestine 7 , denditric cells (DC) into the spleen in particular and in lymphoid tissues in general.8,9 LTβR signaling is also essential for the development and function of high endothelial venules, 10 the induction of lymphoid tissue chemokines and adhesion molecules 11 and the expression of mucosal addressin cell adhesion molecule-1 in DSS-induced intestinal inflammation. 12 These findings reveal the importance of LTβR and ligand interaction in the control of the trafficking of lymphoid cells.
In addition, LTβR signaling is also important in the maturation step, as well as the in situ survival of lymphoid cells. For example, the interaction between LTβR on bone marrow (bm) stromal cells and LT on NK cells is an important step in early NK cell development. Furthermore, survival signals via LTβR activation protect lymphocytes from apoptosis, for instance during virus infection. 13 Hence, LTβR signaling clearly plays a role in the control of local inflammatory reactions via lymphocyte attraction, maturation and survival.
Given that LTβR and its ligands are involved in lymphoid structure formation, which continues to develop in early life,14–16 we generated bm chimeras with neonatal or adult mice to compare the effect of LTβR signaling during maturation. Neonatal transplantation of human haematopoietic stem cells is commonly used in the generation of ‘humanized’ mice, and displayed high transplantation efficiency when transplanted into the liver of newborn mice.17,18
To elucidate the importance of LTβR signaling in the control of inflammatory processes, these mice chimeras were subjected to DSS-induced colitis, which represents a model of inflammatory bowel disease (IBD). IBD comprises different chronic or recurring immune response, and inflammation of the gastrointestinal tract. The main forms of IBD are Crohn’s disease and ulcerative colitis, which can trigger the development of colorectal cancer and limit the quality of life in general. The etiology of IBD is still unknown and, therefore, the only available treatments focus on unspecific immunosuppression. In colitis, the role of LT function has been shown to be as important as the TNF-α signaling for disease development and maintenance. Therefore, understanding the role of activation of the LTβR during this inflammatory disorder might lead to the development of more specific treatment schedules with reduced side effects.
Materials and methods
Animal husbandry
Mice used in all experiments were housed under specific pathogen-free conditions in the animal facility of the University of Regensburg, Regensburg, Germany, and regularly monitored for the absence of specific pathogens, as recommended by the Federation of Laboratory Animal Science Associations.
Bm chimeras
Bm cells were isolated from femur, tibia and humerus of donor mice [C57Bl/6, LTβR−/− and C57Bl/6 mice congenic for CD45.1 (Ly5.1)] flushed out with cold PBS, filtered through a 40-µm nylon cell strainer and centrifuged at 300 g at 4℃ for 10 min. Neonatal transplantations were performed by intrahepatic injection of 3 × 107 bm cells into 24–48-h-old mice 3 h after 5-gray (Gy) radiation. Eight wks post-transplantation reconstitution levels of the donor cells were analyzed in the peripheral blood by flow cytometry.
Adult transplantation was performed by injection of 6 × 106 bm cells intravenously in mice at 8–10 wks of age following two doses of 5-Gy radiation. Three and 6 wks post-transplantation peripheral blood was obtained and analyzed for reconstitution levels of donor cells as described for neonatal transplanted mice. A total of 35 neonatal and 29 adult transplanted chimeras were generated, and 30 (neonatal) and 29 (adult) mice used for colitis experiments.
DSS-induced colitis
Acute colitis was induced in all chimeras by addition of 3% DSS MP Biochemical (Solon, Ohio, USA) to the drinking water for a period of 7 d.
The animals’ mass and general conditions were monitored on a daily basis. On d 7, animals were sacrificed and all organs were harvested for myeloperoxidase (MPO), histology and single cell analyses in flow cytometry.
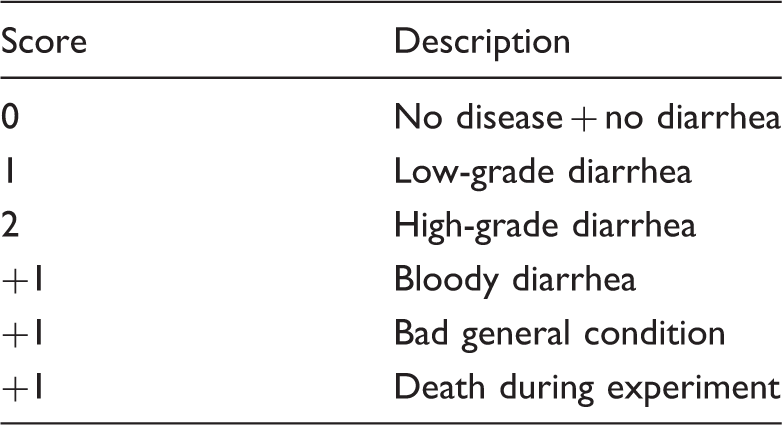
Disease score
Disease score.
MPO
To measure neutrophilic granulocyte infiltration tissue, samples from the middle part of the colon were harvested and prepared as described previously. 12 Briefly, tissue samples were placed in a solution of potassium phosphate buffer (50 mmol/l, pH 6.0) containing 0.5% (w/v) hexadecyltrimethylammonium bromide (1 ml/30 mg tissue). After homogenization (Ultra Turrax; IKA Labortechnik, Staufen, Germany), samples were frozen and thawed three times, centrifuged at 44,000 g at 4℃ for 20 min, and 10 µl of the supernatants were transferred into phosphate buffer (pH 6.0) containing 0.17 mg/ml 3,3′-dimethoxy-benzidine and 0.0005% hydrogen peroxide (H2O2). The H2O2-dependent oxidation of 3,3′-dimethoxybenzidine was measured for MPO activity.
Single cell isolation from different organs
Single cells were isolated from peripheral blood and from different mouse tissues, such as spleen, lymph nodes and bm, as described previously. 19 Briefly, spleen and lymph nodes were cut into small pieces and pressed through a 40-µm nylon filter using the plunger of a 2 ml syringe. Cells were washed with PBS supplemented with 5% FCS and centrifuged at 300 g at 4℃ for 10 min. For bm cells, femurs were collected, both ends were clipped off and cells flushed out with PBS using a 25-G needle, filtered through a 40-µm nylon cell strainer and centrifuged at 300 g at 4℃ for 10 min.
Flow cytometry
Reconstitution levels of donor cells in peripheral blood were determined using CD45.1 phycoerythrin (PE) (clone A20; Becton-Dickinson (BD) Biosciences (San Jose, CA, USA)) and CD45.2 FITC-conjugated Abs (clone 104; eBioscience) together with anti-CD3 APC or Percp-Cy5.5 (clone 145-2C11; eBioscience) and anti-B220 Percp-Cy5.5 (clone RA3-6B2; eBioscience (San Diego, CA, USA)). Samples were further analyzed using the following mAb from BD Bioscience (San Diego, CA, USA): anti-Ly6C FITC or biotin (clone Al-21), anti-Ly6G PE (clone 1A8), and anti-CD11b FITC (clone M1/70). The following Abs from eBioscience were used in addition: anti-Gr-1 APC (clone RB6-8C5); anti-CD11c Alexa700 (clone N418); anti-F480 Pacific Blue (clone BM8). For detection of LTβR-expressing cells, anti-LTβR biotin (clone 3C8) from eBioscience was used. All samples were analyzed using a FACSLSR II flow cytometer with the Diva software package Becton-Dickinson (BD) Biosciences (San Jose, CA, USA). To reduce non-specific binding of mAb, cells were incubated with anti-mouse Fc receptor Abs (10 µg/ml; Sigma-Aldrich (Sigma-Aldrich, Taufkirchen, Germany)) on ice for 20 min before staining with specific mAb or isotype control mAb.
Histology
Histological score.

Musc.: lamina muscularis mucosae.
Statistics
Statistical analysis was performed using GraphPad’s Prism software (GraphPad, La Jolla, CA, USA). A Student’s t-test was applied for MPO, colon length and flow cytometric data. Two-way ANOVA + Bonferroni post-test were used for weight loss analyses, and a Mann–Whitney rank sum test was employed for histological score, disease score and colon length. Significance was set at P > 0.05.
Results
Bm transplantation of adult, as well as of neonatal, mice led to high reconstitution levels in all organs, but did not influence lymphoid structure formation
To elucidate the effect of LTβR signaling during hematopoiesis in a growing organism, we compared two different bm cell transplantation models in this study: one transplantation method using adult mice (aged 8–10 wks) and a transplantation model using neonatal mice (aged 24–48 h). Thus, we were able to compare the influence of transplantation on immune cell differentiation at an early stage of hematopoiesis with the adult model. Importantly, both transplantation methods revealed equally high reconstitution levels in all tissues, including the peripheral blood, the bm, the spleen and lymph nodes, independent of the time point of transplantation (neonatal: Figure 1a; adult: Supplementary Figure 1a). In both setups, reconstitution was stable for more than 1 yr, but neonatal transplantation resulted in decelerated growth of the animals owing to the irradiation dose of 5 Gy. Nevertheless, lymphoid structure formation could not be rescued by the wild type (wt) bm transplantation in LTBR−/− neonatal mice as only one out of 13 transplanted neonatal LTBR−/− mice developed mesenteric lymph nodes. These findings have also been observed in adult transplantation in LTBR−/− mice, which did not result in lymph node formation (0/9). All other transplanted groups (including neonatal wt mice transplanted with LTBR−/− bm) developed mesenteric lymph nodes (even; data not shown).
LTβR−/− bm-transplanted chimeras revealed aggravated signs of inflammation and tissue damage after DSS colitis. (a) Flow cytometry of peripheral blood (pb) of bm reconstituted mice (neonatal transplantation; 14 wks post-transplant) analyzed by the expression of the congenic markers Ly5.1 (CD45.1) in C57Bl/6 and LTβR−/− mice, and Ly5.2 (CD45.2) in CD45.1 mice. Inflammation and tissue damage after DSS-induced colitis were measured as MPO activity (b) in U/mg of protein isolated from the distal part of the colon; histological score (d) representing mucosal damage and inflammatory infiltrates, as described in Table 2; and reduction of colon length (e) as a marker of injury measured in cm of colon. (f) Mass was monitored daily during DSS treatment, and disease score (g) was assessed on the day of sacrificing the animals (Table 1). (c) Representative H&E-stained colon section of C57Bl/6 mice transplanted with CD45.1 bm, LTβR−/− mice transplanted with CD45.1 bm and CD45.1 mice transplanted with LTβR−/− bm. Bars represent 100 µm. Data were analyzed using Student’s t-test (MPO, colon length), one-way ANOVA (A* = P < 0.05) + Tukey’s multiple comparison test (histological score, MPO) and two-way-ANOVA + Bonferroni post-test (mass loss). Error bars represent means ± SEM. *P < 0.05), **P < 0.01). ln: lymph nodes.
Chimeras transplanted with LTβR-deficient bm cells developed exacerbated inflammatory reactions in DSS-induced colitis
After 7 d of DSS treatment, mice transplanted with LTβR−/− bm cells demonstrated (a) MPO activity, indicating granulocyte influx into the tissue (Figure 1b, Supplementary Figure 1B) and (b) the most serious tissue damage, including loss of crypts, severe inflammatory infiltrates and loss of mucosal architecture (Figure 1c), resulting in an increased histological score (Figure 1d) in both transplantation models. In addition, the reduction of colon length as a result of on-going inflammation was much more pronounced in mice lacking LTβR on hematopoietic cells than in mice lacking LTβR expression on stromal cells (Figure 1e, Supplementary Figure 1C). Both transplantation strategies revealed a significant worsening of disease and mass loss when bm cells lacked LTβR expression (Figure 1f, g, Supplementary Figure 1D, E). Thus, in both models LTBR deficiency led to an exacerbation of the inflammatory response upon DSS and therefore supports the earlier finding that LTβR signaling on hematopoietic cells (and not on stromal cells) down-regulates exaggerated inflammation in a model of acute DSS-induced colitis. 6 This result was independent of whether the bm transplantation occurred in neonatal or adult animals.
Myeloid cell proportion increased in peripheral blood after DSS-induced colitis
Peripheral blood of neonatally-transplanted chimeras was analyzed by flow cytometry 6, 10 and 14 wks after bm transfer. The results show an increase of B cells (Figure 2a) in parallel with a slight decrease of T cells (Figure 2b) and myeloid cells (CD33+; Figure 2c), independently of whether mice were transplanted with LTβR−/− or LTβR+/+ bm cells. Such changes in the lymphocyte composition were not observed in adult transplanted animals (data not shown), which already had a solid B cell population 2 wks post-transplantation.
Cell composition in peripheral blood changes after DSS-induced colitis in all chimeras. Peripheral blood (pb) analyses of B cells (a), T cells (b) and myeloid cells (c) in neonatal-transplanted chimeras 6, 10 and 14 wks post-transplant (without colitis) and at d 7 after DSS-induced colitis (arrow). Chimeras [CD45.1 bm in LTβR−/− mice (n = 4), LTβR−/− bm in CD45.1 mice (n = 5)] were analyzed for CD11c+ cells (d) and their subsets CD11c+Gr1+Ly6c+ (E) and CD11c+Gr1−Ly6c+ (g). (f) Density plot of peripheral blood showing two cell populations (indicated by arrows) that were increased after DSS colitis. Error bars represent means ± SEM. Data were analyzed using Student’s t-test. *P < 0.05).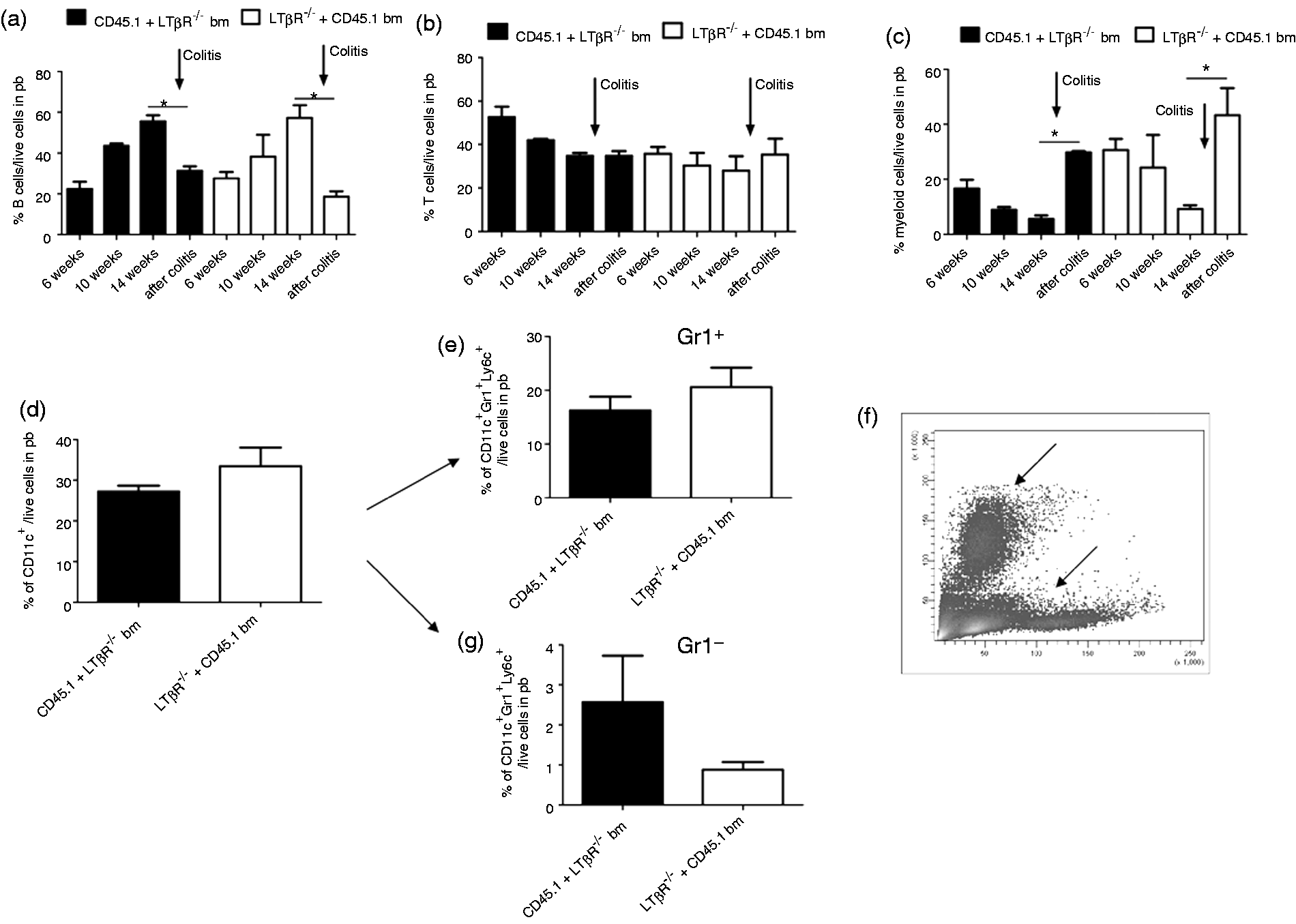
In both adult and neonatal chimera models an equal change in cell composition was found after 7 d of DSS-treatment characterized by a significant decrease of the B cell fraction (Figure 2a) paralleled by a significant increase of the myeloid cell fraction (Figure 2c). Further characterization of the myeloid cells revealed that these cells were positive for CD11c. The proportion of these CD11c+ cells (Figure 2d) was similar in both types of chimeras and independent of LTβR expression. Two different myeloid cell populations were found. The larger fraction was characterized by the markers CD11c+ Gr1+ Ly6C+ (Figure 2e) and located in the forward scatter (FSC)/side scatter (SSC) area of granulocytic cells (Figure 2f, top arrow). The smaller fraction with the profile of CD11c+ Gr1− Ly6C+ cells (Figure 2f, bottom arrow, Figure 2g) was slightly higher in animals transplanted with LTβR−/− bm.
Maturation of different splenic myeloid cell population in mice transplanted neonatally with LTβR−/− bm
Spleen cells serve as a reservoir for inflammatory monocytes during inflammation, and extra-medullary proliferation of myeloid cells contributes massively to the monocytic cell pool under inflammatory conditions. Therefore, we further characterized different subsets of myloid cells in this lymphoid organ by flow cytometry (Figure 3), including macrophages (F480), DCs (CD11c) and their subpopulations. Animals transplanted with LTβR−/− bm cells showed a significantly lower percentage of macrophages (Figure 3d, left), but a higher proportion of CD11c+ cells (Figure 3d, second graph). In all chimeras the predominant population of F480+ macrophages was F480+ CD11c− Gr1+ Ly6C− (data not shown). In contrast, the majority of CD11c+ cells were Ly6c++ GR1++ (Figure 3d, third graph), and were located in the FSC/SSC area of granulocytes.
Characterization of myeloid subpopulations in different chimera subsets after DSS-induced colitis. Density plots showing gating strategies for F480+ and CD11c+ cells (a) gated on live cells, Ly6c+ and Gr1+ of CD11c+ cells (b), and Ly6c++ and Gr1+ of CD11c−F480− cells (c). GraphPad Prism illustration of spleen cells separated from neonatal-transplanted chimeras after DSS-induced colitis (d) showing differences of myeloid cell fractions in mice transplanted with LTβR−/− bm in the population of F480+ cells (first graph), CD11c+ cells (second graph), CD11c+F480−Gr1++Ly6c++ (third graph) and CD11c−F480−Gr1−Ly6c++ cells (fourth graph). (e) Dot plots representing CD11b+ and CD11c+ populations of live cells (first plot), and Ly6g and Ly6c on CD11b+ cells (second plot) in peripheral blood (PB). Graphs of pooled mice samples (n = 3) from peripheral blood and spleen of CD45.1 mice transplanted with LTβR−/− bm and CD45.1 mice transplanted with C57Bl/6 bm. Graphs represent the percentage of CD11b+ cells (left graph), CD11b+Ly6c+Ly6g+ and CD11b+Ly6c++Ly6g− of live cells (right graph). Error bars represent means ± SEM. Data were analyzed using one-way ANOVA (A* = P < 0.05) and Tukey’s multiple comparison test. *P < 0.05), **P < 0.01.
Chimeras that had received LTβR−/− bm cells showed a reduced proportion of immature (F480+ Gr1+ CD11c− Ly6C−) monocytic cells in the spleen, independent of whether they had been transplanted as neonates or adults (data not shown).
Significant differences in myeloid subpopulation generation after DSS colitis were observed within a subset of Ly6c++ Gr1− cells. This population was significantly more reduced in adult and neonatal mice transplanted with LTβR-deficient hematopoietic cells (Figure 3d, right graph). These cells could neither be assigned to the population of mature DC (CD11c+) nor to macrophages (F480+). Therefore, we also stained for CD11b, which enables the separation of CD11b+ Ly6c+ Ly6G+ immature granulocytic and CD11b+ Ly6c+ Ly6G− immature monocytic populations (Figure 3e, first and second graph). Both of these subsets of immature myeloid cells were reduced in the blood and spleens of mice transplanted with LTβR−/− bm cells (Figure 3e).
Taken together, the lack of LTβR expression on hematopoietic cells induced a slight shift from mature macrophages toward DC and granulocytes. In addition, a reduced proportion of immature myeloid cells in mice transplanted with LTβR-deficient bm was observed in the spleen and in the peripheral blood (Figure 3e).
Of note, no significant differences in T and B cell proportion were detected in the spleen or the peripheral blood of any chimeras after DSS-induced colitis (data not shown).
Characterization of myeloid subpopulation expressing LTβR in peripheral blood, spleen and lymph node after DSS-induced colitis
Chimeras that had been transplanted with bm cells from wt mice were analyzed for their LTβR expression profile on hematopoietic cells. In peripheral blood, two different cell populations were positive for LTβR staining (Figure 4a, b). One population was characterized by F480+ CD11c− (Figure 4a, fourth graph) consisting of Ly6C-negative and Ly6C-positive cells (Figure 4a, fifth graph). The other population in peripheral blood (Figure 4b) and the LTβR+ populations in the spleen (Figure 4c) did not express CD11c, F480 or GR-1, but some cells displayed a high expression of Ly6C. Of note, this CD11c- and F480-negative myeloid cell subpopulation, which had the marker profile typical for immature monocytic inflammatory, or so-called ‘suppressor’ cells, was significantly reduced in mice that had been transplanted with LTβR−/− bm cells (Figure 3d, fourth graph). Only a few LTβR-expressing cells were detectable in the lymph nodes, mainly without expression of any of the tested markers (Figure 4d).
LTβR expression on different cell populations in peripheral blood, spleen and lymph node after DSS-induced colitis. (a–d first plot) Morphology plot of peripheral blood, spleen and lymph node of chimeras transplanted with wt bm. The second and third plots display isotype and LTβR staining of gated live cells from the population indicated in the morphology plot (arrows). Third (and fourth) plot outlines CD11c and F480 (and Ly6c and Gr, respectively) expression profile of LTβR+ cells.
Discussion
LTβR and its activation by the ligands LIGHT, LTα3 and LTα1β2 are involved in many immunologic processes, including immune cell induction, activation and recruitment, as well as lymphoid structure formation. The important role of LTβR signaling in the DSS colitis model was described previously and showed an enhanced inflammatory reaction when LTβR signaling is blocked.4–6
Because the LT system is involved in the development and maintenance of lymphoid structures in general,1–3 and at the mucosal site of gut-associated lymphoid tissue, in particular, 20 we used two transplantation models (neonatal and adult transplantation) to determine whether the expression of LTβR at different time points during hematopoiesis influences disease progression later in life.
Interestingly, both transplantation strategies resulted in very similar findings: wt bm transplantation in LTBR−/− mice was not able to rescue lymphoid structure formation, even when transplanted in newborn mice. Therefore, lymphoid structure formation is dependent on LTBR expression on stromal cells and not on hematopoietic cells. The lack of mesenteric lymph node formation in the LTBR−/− recipients does not seem to hinder acute inflammation processes in the gut. LTBR−/− and control mice (wt + wt bm) demonstrated comparable immune response, shown cumulatively by a similar inflammation profile, MPO activity, histological score and colon length. Furthermore, it has been previously reported that extra-lymphoid tissues play an important role in the induction of immune responses.21,22 In addition, LTBR−/− mice revealed similar allograft rejection as wt mice, but this graft-versus-host disease (GvHD) reaction was delayed in splenectomized LTβR−/− mice. 23 Therefore, immune activation, which usually occurs in the peripheral lymph nodes, can be compensated for by the spleen in LTBR−/− animals. Nevertheless, the influence of missing lymph nodes in LTBR−/− mice during DSS-induced colitis cannot be excluded completely.
During DSS-induced colitis, the lack of LTβR on hematopoietic, but not on stromal, cells led to an increase in inflammation and tissue damage, which was clearly demonstrated by the very high inflammatory cell influx and MPO activity. These results indicate that activation of the LTβR on hematopoietic and not on stromal cells is essential for restraining the inflammation. Thus, LTβR expression on hematopoietic cells seems to be important to dampen on-going inflammation, and to block the recruitment and proliferation of inflammatory cells in the tissue.
Characterization of different peripheral blood subpopulations after DSS-colitis showed an LTβR-independent significant reduction of B cells and increase of myeloid cells. The main subpopulation of the increased myeloid cell fraction in all chimeras were characterized as granulocytes (CD11c+Gr1+Ly6C+), which also entered the intestinal area, as indicated by the MPO activity.
Because LTβR−/− mice do not generate lymph nodes, we analyzed spleens as lymphoid organs to look for differences in the hematopoietic cell fraction. None of the chimeras showed significant variations in the splenic B and T cell fractions. However, differences in the myeloid cell fraction were found. Chimeras (adult and neonatal transplanted) that lacked LTβR on hematopoietic cells showed significantly fewer macrophages, and a significantly reduced myeloid cell population, which lacked F480, CD11c and Gr1 markers, but expressed Ly6C. Interestingly, this population of immature myeloid inflammatory cells was also partly positive for LTβR expression in blood, spleen and lymph nodes in chimeras transplanted with wt bm.
We further characterized this fraction as CD11b+ cells being either Ly6C- and Ly6G-positive or Ly6Chigh Ly6G-negative. Both cell populations were reduced in chimeras transplanted with LTβR−/− bm. These subsets of CD11b+ cells are also described as immature granulocytic or monocytic cells, respectively. They belong to the immature myeloid cell lineage with immunosuppressive functions [myeloid-derived suppressor cells (MDSC)] and were decreased in chimeras without LTβR on hematopoietic cells.
Such suppressive immature cells represent a heterogeneous population24–27 with multiple immunosuppressive functions27–33 and have been described in tumors,25,34–36 infection,37–39 and during GvHD 40 and auto-immune encephalitis. 41 Importantly, these cells were also involved in the regulation of IBD by inhibiting T cell-mediated intestinal injury. 42
The highest degree of intestinal inflammation in our chimeras reconstituted with LTβR−/− bm was seen in mice where such immature myeloid cells were reduced. Interestingly, this myeloid subpopulation expressed LTβR in wt chimeras. Therefore, LTβR expression on hematopoietic cells might be involved in induction or recruitment of immature myeloid cells in colitis, resulting in amelioration of inflammation.
Conclusions
In conclusion, our experiments indicate that activation of the LTβR could play a role in the induction or recruitment of myeloid suppressor cells in inflammatory processes. The results of the present study demonstrate that LTBR expression on stromal cells is essential for lymph node formation, whereas the regulation of inflammation is controlled by LTBR expression on hematopoietic cells. In addition, the maturation of myeloid subpopulation is also influenced by LTBR expression, and lack of this receptor results in an increased proportion of DCs, and reduced proportion of macrophages and myeloid progenitor cells (MDSCs).
Footnotes
Funding
This work was supported by the Deutsche Forschungsgemeinschaft (grant number: HE3116/8 to TH).
Conflict of interest
The authors do not have any potential conflicts of interest to declare.
Acknowledgements
We especially thank Dr Nancie Archin (University of North Carolina at Chapel Hill School of Medicine, USA) for proofreading the manuscript. We thank Prof. O. Kölbl (Department of Radiotherapy, University of Regensburg) for providing access to the linear accelerators for irradiation. We would like to thank M. Bock and K. Holz for excellent technical assistance, and Prof. G. Brockhoff (Clinic of Gynecology and Obstetrics, University of Regensburg) for the excellent graphic design of the figures.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.