
Abstract
Antimicrobial peptides or host defense peptides are fundamental components of human innate immunity. Recent and growing evidence suggests they have a role in a broad range of diseases, including cancer, allergies and susceptibility to infection, including HIV/AIDS. Antimicrobial peptide elicitors (APEs) are physical, biological or chemical agents that boost human antimicrobial peptide expression. The current knowledge of APEs and their potential use in the treatment of human infectious diseases are reviewed, and a classification system for APEs is proposed. The efficient use of APEs in clinical practice could mark the beginning of the urgently needed post-antibiotic era, but further trials assessing their efficacy and safety are required.
Keywords
Introduction
Antimicrobial peptides (APs; also called host defense peptides or innate defense peptides) are a group of fundamental components of innate immunity. Recent and growing evidence supports their role in a broad range of diseases, including cancer, allergies and susceptibility to infection, including HIV/AIDS. 1 The most studied APs are α- and β-defensins, both families are comprised of cationic peptides of 2–6 ku, with three pairs of disulfide bridges and the linear antimicrobial peptide LL-37, the only member of cathelicidin family in humans. 2 Other reported APs in human are histatins, dermcidin and psoriasin.2,3
APs can limit microorganism virulence directly or indirectly by enhancing the host immune system. 4 Direct administration of APs has been proposed; however, the high costs of APs and the acquired resistance observed in vitro makes antimicrobial peptide elicitors (APEs) a logical and potential alternative.1,5 Enhancing innate immunity as a way to combat infection in humans was first suggested in 2004. 6 For example, in a bioterrorist scenario where it would not be possible to identify the infectious agent(s), up-regulation of the hosts innate armament might provide effective treatment. 6 However, this systemic strategy could be energetically unfavorable for the host.
Proposed classification of APEs and their effects on selected human APs in different human tissues and cell lines. AP genes (italics) codify for the following peptides: CAMP, LL-37; DEFB1, hBD-1; DEFB4, hBD-2; DEFB103, hBD-3; DEFB104, hBD-4; HAMP, hepcidin; S100A7, psoriasin; HIS1, histatin 1.
Why do we need alternatives to current antibiotics?
Antibiotics have been the treatment of choice for infectious diseases for more than 65 years. However, the drawbacks of antibiotic-based treatment include virulent multidrug-resistant bacterial strains and adverse reactions.The initial cases of antimicrobial resistance occurred in late 1930s and 1940s, soon after the introduction of the first antibiotic classes, sulfonamides and penicillin. 7 As Rene Dubos wrote in 1942, ‘susceptible bacterial species often give rise by training to variants endowed with great resistance to these agents’, leading, in some instances, to ineffective compounds. Mathematical models and clinical experience suggest that it takes much longer for reductions in antibiotic resistance than for antibiotic overuse to induce resistance in the first place. 8 Adverse drug reactions include fevers induced by β-lactam antibiotics and sulfonamides, and ototoxicity induced by aminoglucosids, Stevens–Johnson Syndrome (toxic epidermal necrolysis), secondary toxicity to nitrofurantoine, trovafloxacin-induced secondary hepatic necrosis, multiple antibiotics allergy syndrome and fatal outcome of anaphylaxis.9,10 Thus, there is a lack of a confident treatment of many human infectious diseases, and the microbial resistance to drugs is increasing.7,11 This resistance dramatically reduces the possibility of treating infections effectively and increases the risk of complications and even fatal outcomes. 12 Data from several countries reveal that 2–7% of total hospital admissions were due to adverse drug reactions (ADRs), and 16–23.3% of these admissions were due to antibiotics. Also, ADRs cause longer stays and increased healthcare expenditures.13,14 An increased understanding of innate immune activation pathways and adaptive immune cross-talk could lead to more effective, faster and safer treatments to combat infections. 1
Why is direct administration of APs not the best approach?
Antimicrobial peptides are ribosome-synthesized innate immunity effectors that appear to be an ancestral defense mechanism against environmental microorganisms. 15 APs have remained effective against bacterial infections for at least 100 million years, despite the continual presence of these peptides in bacterial environments, and it has been suggested that resistance is very unlikely to evolve in the short term. 15 More than 1500 APs have been reported and their expression has been demonstrated in all tested organisms. 2 Most APs are synthesized as propeptides and after postranslational maturation contain from ∼29 to 42 amino acid residues. 16
In humans, α- and β-defensins, and cathelicidin antimicrobial peptide (LL-37) are the most studied. These peptides are expressed mainly in peripheral lymphocytes and epithelia. Human β-defensin 2 (hBD-2) and LL-37 are induced in inflammation and upon immune challenge. 5 hBD-1 is mainly constitutively expressed but is also up-regulated by microbial or inflammatory stimuli. 1 Although many AP-based therapeutic agents are now in clinical trials as antimicrobial and immune regulators, 17 the cost to produce and purify even short APs remains high. 1
In vitro resistance to increasing concentrations of pexiganan, a magainin analog, appears in less than 700 bacterial generations (100 serial transfers) in a model of both mutator and non-mutator strains of the Gram-negative Pseudomonas fluorescens and Escherichia coli, suggesting that naturally occurring mutation rates are sufficient to create selectable variation in the populations. 18
In addition to resistance to APs, administration of cationic APs can promote pathogenicity by a PhoP-PhoQ-mediated system.
19
Salmonella enterica sv. Typhimurium PhoP-PhoQ system regulates hundreds of genes encoding the majority of virulence properties, including intracellular survival, invasion, lipid A structure, phagosome alteration and resistance to APs. Acidic pH and APs activate PhoP/PhoQ regulon and these signals, present within phagosomes and intestinal tissues, may serve as specific signatures of the host environment for Gram-negative pathogens, leading to modification of bacterial phospholipids and LPS, and thus mitigating actions of APs.19,20 The PhoQ homologue in S. enterica sv. Typhimurium exhibits an additional capacity for recognizing cationic APs via structural alterations—novel helices and acidic residues—that still allow for divalent cation binding and may allow for increased activation by acidification. Finally, these differences in ligand recognition and binding mechanism appear to have important functional implications to Salmonella virulence, suggesting that PhoQ sensing in Salmonella facilitates a pathogenic lifestyle.
20
Also, protease synthesis likely degrades certain APs. In parallel, a distinct two-component regulatory system, PmrA/PmrB, ultimately leads to synthesis and addition of 4-amino-4-deoxy-
Also, it is very unlikely for pathogens to develop a response against a signaling molecule that does not interact or interfere directly with their pathogenic mechanism. Compared with APs, APEs have been revealed as an affordable and safe alternative to antibiotics in the treatment of pulmonary tuberculosis with vitamin D
23
and in the treatment of acute diarrhea in children with
Chemical, physical and biological APEs
In this section, APEs are classified as physical, chemical or biological according to Table 1.
Class I: Physical APEs
Subclass A: UV light, a physical elicitor of skin APs
Antimicrobial peptide selectivity against common human pathogens.

+), active in vitro and/or in vivo against the pathogen; (−), not active; NR, not reported.
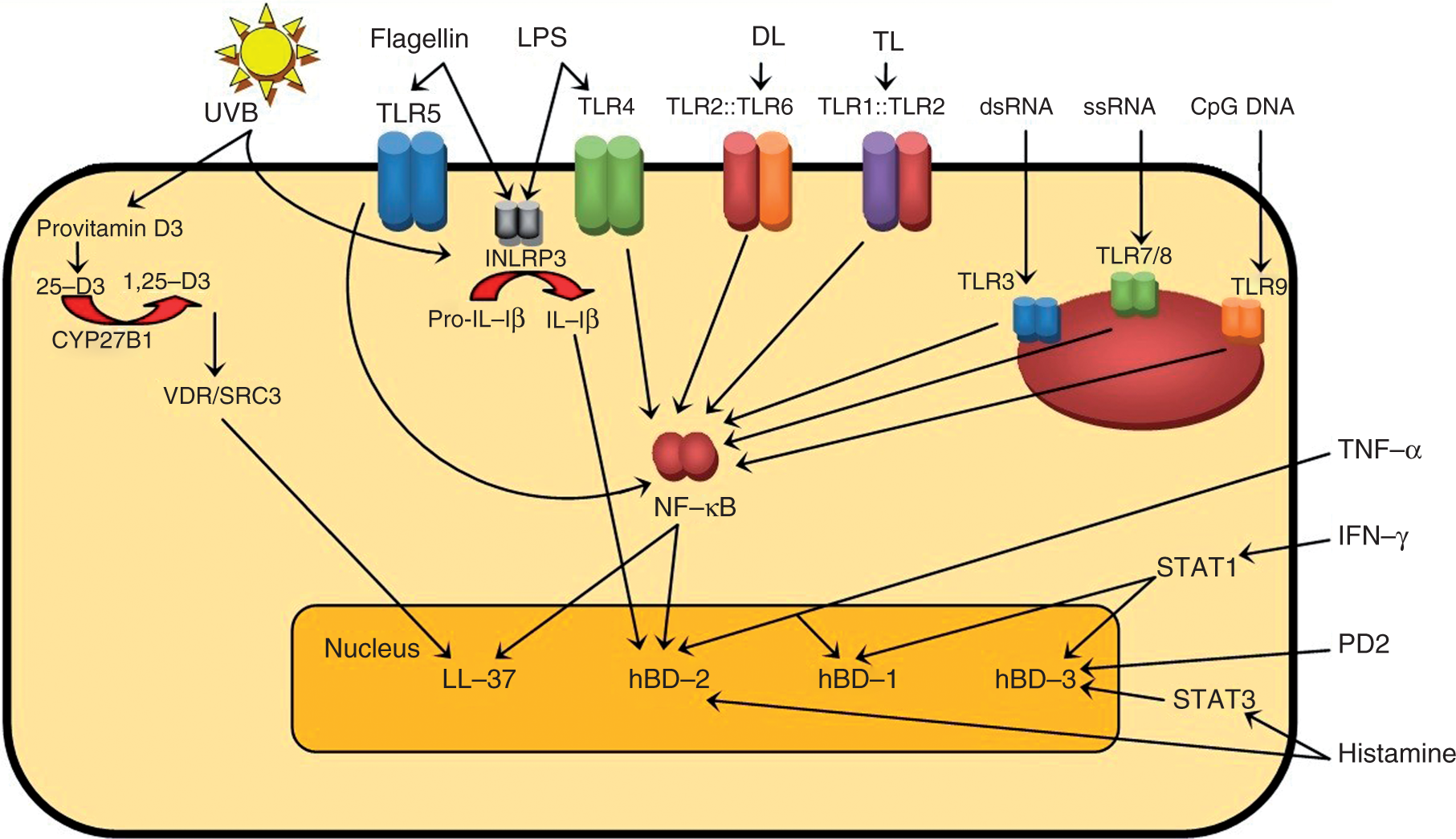
Pathways of APEs in human keratinocytes. From provitamin D3 (7-dehydrocholesterol), UVB produces the synthesis of calcidiol (25-hydroxy vitamin D3 or 25,D3) and then by 25-hydroxyvitamin D3 1-α-hydroxylase 1 (CYP27B1) action, produces 1,25 D3, the active form of vitamin D. 1,25 D3 binds and activates the vitamin D receptor (VDR) which recruits co-activator proteins. Activated VDR forms a complex with steroid receptor co-activator 3 (SRC3), leading to recruitment of histone acetyltransferases. Histone acetylation activates chromatin, presumably facilitating access to transcription machinery in CAMP gene producing LL-37 peptide; 132 however, histone acetylation in CAMP promoter is still controversial and an indirect activation through up-regulation of transcription factors that bind CAMP promoter has been suggested. 84 A second UVB-mediated AP expression pathway is through the inflammasome nucleotide oligomerization domain-like family, pyrin domain-containing 3 (INLRP3). The activated caspase 1 of INLRP3 processes pro-IL-1β into biologically active IL-1β, 133 which is an elicitor of hBD-2.69,134 Psoriasin, hBD-1, hBD-2 and hBD-3 are also up-regulated by UVB, but whether this expression is VDR-dependent is unknown.25–27 Flagellin through TLR5, LPS through TLR4, diacyllipopeptide (DL) through heterodimers composed of TLR2 and TLR6, triacyllipopeptide (TL) through heterodimers composed of TLR1 and TLR2 activates NF-κB. In endosomes, dsRNA through TLR3, ssRNA through TLR7/8 and unmethylated CpG DNA through TLR9 also activates NF-κB. 100 NF-κB activation leads to LL-37 and hBD-2 expression. Psoriasin is up-regulated through TLR5, 130 probably via the NF-κB pathway (not shown). LPS induces hBD-1 by an unknown pathway and hBD-2 through NF-κB. 135 In endosomes, TLR3, TLR7/8 and TLR9 also activate NF-κB. 100 TNF-α induces up-regulation of hBD-1 and hBD-2. 25 IFN-γ through JAK-signal transducer and activator of transcription (STAT1) induces hBD-1 and hBD-3. 69 Prostaglandin D2 (PD2) up-regulates hBD-3. 136 Histamine induces hBD-2 and hBD-3, the latter through STAT3.137,138 Detailed expression pathways for most APEs are unknown.
It has also been suggested that higher exposition to UVB reduces susceptibility to dental caries through production of vitamin D, followed by induction of cathelicidin and defensins. 34 In particular, hBD-3 is highly active against Streptococcus mutans, a pathogen involved in cariogenesis. 35
Class II: Chemical elicitors
Vitamin D (subclass A) and nucleic acids (subclass B) as elicitors
Vitamin D deficiency is associated with poor immune function and with an increase in susceptibility to infectious diseases. 36 Topically applied vitamin D active form (vitamin D3 or cholecalciferol) can decrease the prevalence of thymine dimers (a hallmark of UV-induced DNA damage in skin), but does not reverse UV-induced immunosuppression in humans 37 or mice.36,38,39 Epidermal Langerhans cells harboring UV-induced DNA damage induce regulatory T cells (Treg). IL-12 and IL-23 inhibit the suppressive activity of these Treg. 40 In models of local immunosuppresion, it is proposed that skin-derived dendritic cells (DCs) with damaged DNA move to draining lymph nodes, suboptimally present antigens, induce tolerance and produce antigen-specific Treg cells. UV-induced Treg cells then switch APCs from a stimulatory to a regulatory phenotype and thus the immune phenotype is maintained. 41 For systemic immunosuppression, the mechanisms that alter immune responses are not clear. 36 In psoriatic lesions, vitamin D and its analogs down-regulate hBD-2 and hBD-3. Contrarily, vitamin D up-regulates CAMP mRNA, but not the LL-37 processed peptide, probably explaining why vitamin D does not exacerbate psoriasis. 42 Vitamin D3 produces LL-37 expression in keratinocytes, monocytes, neutrophils,43,44 PBMC, 45 colon cancer cells, bone marrow, B cell lymphomas, prostatic, endometrial cancer cells 46 and also in numerous epithelial cells 47 and tissues. 48 In addition, calcipotriol, a vitamin D analog, induces LL-37 expression in human skin in vivo. 49
Vitamin D3 produces a LL-37-dependent anti-Mycobacterium tuberculosis response 31 and this expression could offer new treatments for this infectious disease, for example one single vitamin D dose improves in vitro activity against Mycobacterium bovis BCG. 50 Three doses of 2.5 mg of vitamin D3 hasten sputum culture conversion in patients with TaqI vitamin D receptor (VDR) tt genotype. 23 Furthermore, 1,25-D induces secretion of IL-10 and PGE2 in M. tuberculosis-infected PBMC, and both proteins are transcriptional regulators of matrix metalloproteinases (MMP) expression. In addition, 1,25-D3 controls M. tuberculosis infection by attenuating expression of MMP-7 and MMP-10, and suppresses secretion of MMP-7 by infected PBMC. 51
Irrespective of infection, 1,25-D3 inhibits MMP-9 gene expression, secretion and activity. 51 During M. tuberculosis infection, ESAT-6 induces MMP-9 in epithelial cells. MMP-9 enhances recruitment of macrophages presumably contributing to nascent granuloma and bacterial growth. 52 In this way ESAT-6 and vitamin D3 have antagonic roles in MMP-9 expression, and it has been suggested that this vitamin has a role in granuloma formation.53,54 Also, 1,25-D3 has been suggested to play a role both at the onset of infection and in the development of the granuloma in infected cattle. 55 Vitamin D and some analogs block IL-17 A-induced hBD-2 expression by increasing IκB-α protein and through inhibition of NF-κB signaling. 56 Furthermore, topically applied 1,25-D3 enhances the suppressive capacity of CD4+CD25+ cells from the draining lymph nodes in mice. 39
Vitamin D also impacts the phenotype and function of DCs by down-regulating expression of co-stimulatory molecules, such as CD40, CD80 and CD86, as well as IL-12 and IL-10. 57 IL-10 is capable of down-regulating hBD-2 and LL-37 expression. 58 LL-37 promotes DCs differentiation by up-regulating endocytic capacity, up-regulating co-stimulatory molecule expression, enhancing secretion of TH1-inducing cytokines and promoting TH1 responses in vitro. 59 Surprisingly, LL-37 also inhibits DC activation by TLR ligands (well known APEs, such as LPS, lipoteichoic acid and flagellin). In addition, LL-37 provokes naive T cells to produce less IL-2 and IFN-γ, and to decrease their proliferation. LL-37 also delays the response of memory T cells to a recall antigen. 60 Vitamin D and VDR deficiency exacerbate experimental autoimmune diseases, such as inflammatory bowel disease, owing to increased numbers of IL-17 and IFN-γ secreting T-cells with a consequent reduction in Treg. 61 IFN-γ enhances TLR2/1 L induction of the 1-α CYP27B1, potentiating the conversion of 25-D3 to 1,25-D3, leading to VDR activation and expression of CAMP (coding for LL-37) 62 (Figure 1).
Oligodeoxynucleotides (ODNs) that contain the nucleotide pair CpG (CpG ODNs) are strong immunomodulators that have shown promising results both in vitro and in vivo, including mice and primate models for treating infections. The unmethylated cytosines of CpG ODNs are recognized by TLR9 as a bacterial signal triggering several innate immune responses. These responses include NK cells, DCs and macrophage activation; adaptive responses, such as TH1 polarizing ability, release of chemokines and cytokines (IL-6, IL-12, IL-18) and activation of CD8+ T cells;6,63 and inhibition of IFN type I (α/β). 64 When CpG ODNs are conjugated with antigen as adjuvants, the immune response is ∼100-fold stronger. 65 In addition, liposome-encapsulated CpG-DNA that contains immunomodulatory motifs up-regulates hBD-2 expression, as well as that of MHC class II molecules (HLA-DRA) in human B cells, 66 but not in keratinocytes nor in colon epithelial cells. 67 In keratinocytes, CpG ODNs induce LL-37 with no effect in hBD-2 (Table 1).
Amino acids, proteins (subclass C) and carbohydrates (subclass D) as elicitors
Arginine, isoleucine and BSA induce hBD-1 transcription and secretion in human colon cells HCT-116. Up-regulation of hBD-1 is concomitant with c-myc overexpression. 68 Interleukins of proteinic nature, such as IFN-γ, up-regulate human DEFB1 (coding for hBD-1), 69 DEFB103 (coding for hBD-3) and DEFB104 (coding for hBD-4) (Table 1 and Figure 1 for its effect in human keratinocytes). IFN-γ is relevant in immune response modulation, antagonizing IL-10 effects and inhibiting some TLR-induced genes. 70 For additional interleukins see Table 1.
In obesity, a low grade inflammatory state, inflammation also occurs in the central nervous system and alters hypothalamus function, resulting in a leptin-resistant state that is mediated by IKKβ/NF-κB and endoplasmic reticulum stress. 71 The leptin-dependent hBD-2 up-regulation 72 could explain why β-defensins are down-regulated in diabetic and obese rats when compared with slim control rats. 73 In human HEK-293 and HCT-116 cells, glucose and insulin levels up-regulate DEFB1 expression. 74 Contrarily, hBD-2 and hBD-3 are down-regulated in keratinocytes in a high glucose medium.75,76 These two genes could explain, at least partially, why infections are very common in diabetic patients; however, further research is needed in this area.
Subclass E: Fatty acids and prebiotics as gastrointestinal elicitors
Sodium butyrate (NaB) is a short chain fatty acid salt normally produced by the colon and provides ∼70% of the energy required by healthy colon enterocytes. 77 NaB does not possess direct antimicrobial activity. Shigellosis, also called bacillary dysentery, is an infectious disease and which is the main cause of morbidity, mortality and stunted growth in children in developing countries. It has been proposed that shigellosis pathogenesis is explained by the transfer of the Shigella spp. plasmid to the host, which blocks LL-37 and hBD-1 expression. 78 In a shigellosis rabbit model, stimulation of the epithelial lining of colon with NaB led to the production of CAP-18 (homolog to LL-37 in humans), which is bactericidal for Shigella spp. 79 In treated rabbits, colon inflammation was reduced, CAP-18 was significantly over-expressed and Shigella spp. counts in feces diminished ∼102-fold at 48 h and ∼104-fold at 72 h when compared with control animals receiving only saline solution. 79 Furthermore, intravenous administration of NaB up-regulates CAP-18 in the epithelia of lung, rectum and colon of rabbits, which suggests a systemic effect. Both 4-phenylbutyrate (PB) and NaB counteract the Shigella-induced down-regulation of CAP-18 in lung.80,81 In humans, NaB inhibits bacteria translocation in metabolic stressed colonocytes. 82
Efficacy of PB in bovine mastitis has also been evaluated. PB induces expression of APs and inhibits Staphylococcus aureus internalization in bovine mammary glands. 83 In humans, PB and its analog, α-methylhydrocinnamate (ST7), induce CAMP expression in a MAP kinase-dependent pathway. 84 PBA also induces DEFB1 expression in human bronchial epithelial cells (VA10), but not in leukemia cells (U937). 84 The tissue-specific response to APEs67,84 deserves further consideration in clinical trials (Table 1).
Other commonly used prebiotics (alimentary supplements) promote beneficial bacterial growth or activity in the colon. The molecular mechanisms for these effects have not been defined, as is the case of formulas that act as ‘immune system enhancers’, sometimes causing defensin up-regulation. This results in recruitment of adaptive immune response cells, 68 such as Treg, which inhibit differentiation of effector T cells that otherwise would lead to inflammation.5,85
Retinoic acid is not an elicitor by itself in humans
Retinoic acid-inducible gene-I-like receptors (RLR) are other members of the pattern recognition receptors (PRRs), which are known to be crucial molecules in innate immune responses. 86 In humans, pre-incubation of keratinocytes with retinoic acid (RA) inhibits up to 90% of the up-regulation of hBD-2, -3 and -4 mediated by phorbol-myristate-acetate (PMA), pro-inflammatory cytokines and bacteria. 87 Otherwise, 9-cis RA and 13-cis RA, as well as Ca+ independently, induce serine proteases Kallikrein 5 (KLK5) and 7 (KLK7), which control enzymatic processing of cathelicidin precursors in the skin and regulate the eventual function of the final forms of these peptides. 88 KLK5 and KLK7 are enzymes that control the physical barrier of the stratum corneum. 89
In pigs (Sus scrofa domestica), all-trans retinoic acid markedly induces both cathelicidins protegrin and PR-39 in bone marrow. 90 However, RA does not restore LL-37 expression in NB4 or HL-60 cell lines, 91 but it is a molecule that induces LL-37 maturation. 88 Thus, RA can antagonize the function of APEs or induce maturation of APs, but is not an elicitor by itself in humans because it does not increase CAMP transcription.
Class III: Biological elicitors
Subclasses A–E: Biological elicitors are also potential vaccine adjuvants and immunomodulators
Biological or microbial elicitors are microorganisms that can be beneficial probiotics 5 or attenuated pathogens, 92 causing efficient up-regulation of APs. Probiotics are microorganisms that, when administered in sufficient quantities, confer a health benefit to the host. Pediococcus spp. 93 and Lactobacillus spp. 94 elicit hBD-2. In particular, E. coli 81 and E. coli Nissle 1917 95 elicit hBD-2 through flagellin. 96 Flagellin is a structural component of bacterial flagella, whose elicitory effect is, in part, TLR5-dependent on CD11+ DCs through TH17 differentiation in the CD11chigh/CD11bhigh subset. Flagellin fused with M. tuberculosis protein p27 (gene Rv2108) shows the strongest cellular response and highest IFN-γ secretion when compared with CpG DNA plus Freund’s adjuvant. Flagellin is a promising candidate in vaccine development and it is currently in phase II clinical trials as an adjuvant included in the influenza vaccine. 97 It has been suggested that E. coli Nissle 1917 or its flagellin in gut epithelia may improve response to inflammatory bowel disease. 5 Furthermore, administration of these probiotics or flagellin itself in gut epithelia may improve response to human infectious diseases, as shown in both mice models of Pseudomonas keratitis 98 and pneumonia. 99
The potential approach of biological elicitors could be similar to vaccination, but the main goal of this class of APEs is to elicit and modulate innate and adaptive immunity, and not simply activate adaptive responses with the consequent Ab production. APs are up-regulated through TLRs 100 and nucleotide oligomerization binding domain (NOD)-like receptors (NLRs) signaling101,102 by exposure to chemical, physical and biological elicitors (see Figure 1 for selected APE signaling in human keratinocytes).
Potential application of APEs and theoretical limitations of their clinical use
In the near future APEs could be useful in treating diseases caused or modified by APs deficiencies, such as shigellosis, Crohn’s disease (CD), HIV/AIDS, atopic dermatitis (AD), lepromatous leprosy and cancer among others. 1 CD is a chronic inflammatory bowel disease, caused, at least in part, by a defect in a microbe sensor system of the intestinal cells, i.e. NOD2, which recognizes bacterial muramyl dipeptide. This sensory system defect is associated with deficient production of hBD-2, hBD-3, α-defensin 5 (HD-5) and 6 (HD-6).5,103 Furthermore, CD is more common in patients with ≤3 copy number variants (CNVs) of DEFB4 (hBD-2) and allele DEFB1 668 C has been found to be associated with CD. 104 These data may be useful in a personalized treatment approach to CD taking into account the above mentioned genetic variants of CD patients. Efficacy of APEs may depend on host genome capacity to respond with certain functional allele(s).105,106
A diminished affinity of a putative binding site in the 5′UTR regulatory region of DEFB1 to transcription factor NF-κB1 (p50/p105) when the C allele is present in single nucleotide polymorphism (SNP) rs1800972 (G in non-coding strand) is suggested as one reason of poor innate immunity response to infectious microorganisms and lower pathogen clearance.1,105 The C allele in this locus induces up-regulation in constitutive expression (compared with the G allele). Otherwise, the G allele up-regulates DEFB1 expression in an IFN-γ-dependent expression in a co-dominant manner. 106 The hBD-1 peptide chemoattracts immature DC and memory T cells, 107 and it has been proposed that hBD-1 may be involved in DC maturation and/or .activation.4,108,109 In addition, plasmacytoid DCs and monocytes increase hBD-1 production when infected with PR8, HSV and Sendai virus. 110 It is probable that hBD-1 has a primary effect on the skin inflammation and/or skin responsiveness in many allergic reactions and immune responses in general, acting not only as AP, but also as an immunoregulator.1,4,107,109 DEFB1 gene expression levels, as well as several SNPs mostly in 5′UTR, have been associated with several infectious, inflammatory and allergic diseases.105,108,111–123
Owing to the tissue-specific usefulness of APEs (Table 1), the side effects of up-regulated APs in certain diseases must be considered. LL-37 can be induced in inflammatory biliary disease by endogenous bile salt chenodeoxycholic acid and therapeutic bile salt ursodeoxycholic acid (UDCA), and further up-regulated by 1,25-D3. 124
In AD, LL-37 expression is down-regulated leading to increased rates of infection in affected skin. The expression of LL-37 in skin has been suggested to be a double-edged sword. In AD, a 1,25-D signaling of LL-37 would not be beneficial because it would skew the T helper response towards a TH2 phenotype, due, in part, to the induction of thymic stromal lymphopoietin, which is a 1,25-D target gene in humans.
Otherwise, in rosacea, hCAP-18, the precursor of LL-37, is processed abnormally by stratum corneum tryptic enzyme (SCTE) 125 and CAMP up-regulation could worsen the disease. In psoriasis, LL-37 is up-regulated, and vitamin D analogs have proven therapeutically effective; however, there is a paradox as these analogs up-regulate LL-37 and it binds to DNA, thus activating TLR signaling in plasmacitoid DCs (pDCs) of the psoriatic skin, leading to an autoimmune response. The mechanisms of LL-37 regulation and psoriasis etiology are poorly understood, 126 and CAMP up-regulation in psoriasis should be also contraindicated.
In patients with acute watery diarrhea caused by Vibrio cholera O1 or enterotoxigenic E. coli (ETEC), hBD-2 and HD-5 are down-regulated in duodenal epithelium and Paneth cells, respectively. Interestingly, LL-37 levels are also decreased in acute phase, but CAMP mRNA remain unchanged, suggesting a V. cholerae-dependent post-translational regulation of LL-37. 127 APEs capable of up-regulating hBD-2, LL-37 (Table 1) and HD-5 could be useful as an alternative therapy to Vibrio cholera-caused diarrhea and ETEC.
A recent clinical trial of NaB administered as an enema in adults with shigellosis from Bangladesh shows diminished hemorrhaging, faster epithelial healing and better counteracting action against infection-dependent down-regulation of LL-37 in stools and surface epithelia. 128 According to previous preclinical trials, NaB has the potential to be administered orally and/or systemically.79,80
The modest improvements provoked by APEs in humans when compared with animal models could be because (i) in human populations variants in AP genes are presumably higher (www.ensembl.org), as opposed to animal laboratory strains, which are highly endogamic, thus diminishing the genetic diversity and increasing predictability to APEs responses; (ii) when antibiotics are used in humans, the death of both commensal and potentially pathogenic bacteria could diminish their function as class III APEs, at least in digestive tract epithelia; (iii) other variables, such as levels and diversity of APEs in normal diet, can be harder to control in humans compared with laboratory animals. However, these probable explanations deserve further investigation.
Concluding remarks
The purpose of the review is to convince the reader that APEs are indirect, cheap and, theoretically, safe and effective weapons, even with their apparently modest success in clinical practice. 24 The use of APEs could overcome Stuart Levy’s shortcoming ‘antibiotic paradox’—miracle drugs destroy the miracle 129 —because it is more difficult for a pathogen to develop a way to respond to an APE, which can be a simple molecule as part of a normal diet and/or metabolism. The response to elicitors might depend on the tissue/cells analyzed because the same elicitor causes up-regulation or down-regulation,67,84 or has no impact at all in different cells/tissues (Table 1). Tissue specificity of APEs probably means that one or more of the following explanations exists: (i) tissue-specific regulation of APE receptors; (ii) specific APE transporters not expressed in all cells; (iii) tissue-specific epigenetic signals of accessibility to transcription machinery; (iv) tissue-specific alternative regulatory elements in AP genes; (v) polymorphisms that affect response to elicitors 106 depending on a higher/lower affinity of promoters or enhancers to transcription factors 105 because cell lines and primary cultures are derived from genetically different people.
In summary, a more complete knowledge about signaling pathways and tissue specificity of AP expression/suppression could be useful to design tailored methods to combat infections and other human diseases with the regulation of antimicrobial peptides, natural weapons of host innate immunity. The use of APEs is a promising, but incipient, field because an ideal elicitor must also restore host–microbe balance and not just suppress inflammation and adaptive immunity. 5 The main challenge in developing APEs as treatment for specific diseases is to optimize tailored elicitors with the following characteristics when compared with common antibiotics: (i) lower costs of production; (ii) higher efficacy; (iii) induce fewer and milder side effects; (iv) no generation or lower counts of drug-resistant microorganisms.
In regard to common infection models, caution must be taken in data extrapolation to human diseases, especially those obtained from rodent models. Human and murine innate immune systems differs mainly in (i) intestinal α-defensins (called cryptidins in mice) are expressed and processed differently; 16 (ii) murine neutrophils lack α-defensins, but they comprise 20–40% of the total proteins of azurophilic granules in humans; 16 (iii) mice lack the gene for psoriasin, but S100A7c is present at high concentrations in human stratum corneum; 130 (iv) induction of murine Cramp (CAMP homolog) does not depend on vitamin D response elements (VDREs) and hence its expression is not regulated by vitamin D;57,131 (v) infection of human macrophages with M. tuberculosis and other cell types with different pathogens leads to the repression of the CAMP gene, whereas the murine Cramp gene is induced. 57 Hence, more relevant alternative models to human biology must be sought because expression deficiencies of APs are responsible for several infectious and inflammatory diseases.1,3
The aforementioned ideas open new avenues for innovative treatment methods for many infectious diseases that are still common public health problems worldwide, with the hope of minimizing the potential risk for adverse drug reactions and pathogen multiresistance, which are common drawbacks of the currently used—and, unfortunately, abused—antibiotics.
Footnotes
Acknowledgements
Thanks to the Instituto de Biotecnología (IBT-UNAM, Mexico) and Marcel Roche Libraries (IVIC, Venezuela) for information resources. The author receives a research stimulus from the Sistema Nacional de Investigadores (SNI 41290, CONACYT, Mexico). Thanks to Dr L. Figuera (CIBO-IMSS), E. Sugg, Dr K. Allen (Peace Corps/CIATEJ, AC) and Dr M. Zasloff for reviewing the manuscript.
Funding
This work was supported by the Fondos Sectoriales de Ciencia Básica SEP-CONACYT Mexico (grant number CB-2008-01-105813)
Conflicts of interest
The author declares that his group is working with some elicitors presented in ![]() , as well as with novel ones. We are pursuing a patent for the synthesis methods, uses and, when applicable, the novel molecules generated in the name of CIATEJ AC, with Prado Montes de Oca E, Mateos Díaz JC, Flores Miramontes MG and Chavez Hurtado P as inventors.
, as well as with novel ones. We are pursuing a patent for the synthesis methods, uses and, when applicable, the novel molecules generated in the name of CIATEJ AC, with Prado Montes de Oca E, Mateos Díaz JC, Flores Miramontes MG and Chavez Hurtado P as inventors.