
Abstract
Epithelial to mesenchymal transition (EMT) has been linked to metastasis. Resveratrol exhibits potential antitumor activities; however, the inhibitory effects of resveratrol on the EMT of melanoma have not been demonstrated. Here, a new role for LPS in promoting EMT is described. LPS-induced EMT was identified by examining the markers of EMT. To assess the activation of NF-κB signal transduction pathway, we performed a reporter assay by using tumor cells transfected with the luciferase gene under the control of NF-κB response elements. The antitumor effects of resveratrol were evaluated in an experimental mouse metastasis tumor model. LPS increased N-cadherin and Snail expression and decreased zonula occludens-1 expression in a dose- and time-dependent manner. Meanwhile, LPS stimulated cell migration through activation of TLR4/NF-κB signal pathway. LPS-induced EMT is critical for inflammation-initiated metastasis. Nuclear translocation and transcriptional activity of p65 NF-κB, an important inducer of EMT, were inhibited by resveratrol. Resveratrol inhibited LPS-induced tumor migration and markers of EMT, significantly prolonged animal survival and reduced the tumor size. Thus, resveratrol plays an important role in the inhibition of LPS-induced EMT in mouse melanoma through the down-regulation of NF-κB activity. The data provide an insight into the mechanisms on the function of resveratrol during the processes of EMT.
Introduction
Melanoma is one of the leading causes of malignancy related death in the world. 1 Once melanoma has spread, there are few effective treatment options, thus there is an urgent need for understanding of melanoma progression. Inflammation is central in combating pathogens. Nevertheless, if it fails to proceed through an orderly and timely process, the resulting chronic inflammation may contribute to tumor formation. Chronic inflammation is considered one of the most important factors in tumor development and progression. 2 The inflammation in the tumor microenvironment plays a pivotal role, not only in tumor development, but also in metastasis.
Epithelial to mesenchymal transition (EMT) is a process that results in the loss of epithelial properties, such as cell-cell adhesion, and the gain of mesenchymal properties, such as increased ability to migration. 3 Phenotypic markers for EMT comprise an increased capacity for migration. EMT is a complex, stepwise phenomenon that occurs during tumor progression and is also associated with chronic inflammation. 3 Recent studies demonstrated that proinflammatory cytokines enhance invasion and the EMT of tumor cells.4,5
Resveratrol is found in grapes and red wine and exerts several biologic activities. 6 Resveratrol has been shown to reduce chemically-induced edema and LPS-induced airway inflammation.7,8 The anti-inflammatory mechanism of resveratrol is not completely understood. In this study, we examined the potential mechanism of LPS-induced EMT in mouse melanoma. We suggested that resveratrol could inhibit LPS-induced EMT and that this effect of resveratrol accompanied inhibition of NF-κB transcriptional activity. This study provides novel insights on the intra-relationship between inflammation and EMT induction in tumor and introduces novel agents, such as resveratrol, as potential blockers of the initiating steps of inflammation-related tumor metastasis.
Material and methods
Reagents, cell and mice
Resveratrol, DMSO, polymyxin B and LPS were purchased from Sigma Aldrich (St Louis, MO, USA). Murine K1735 melanoma was cultured in DMEM supplemented with 50 µg/ml gentamicin, 2 mM
Immunoblot analysis
The protein content in each sample was determined by bicinchoninic acid (BCA) protein assay (Pierce Biotechnology, Rockford, IL, USA). Cytoplasmic fraction and nuclear fractions were prepared according to the manufacturer’s instructions (Pierce Biotechnology). Proteins were fractionated on SDS-PAGE, transferred onto Hybond enhanced chemiluminescence nitrocellulose membranes (Amersham, Little Chalfont, UK) and probed with polyclonal Abs against zonula occludens (ZO)-1(Invitrogen, Camarillo, CA, USA), N-cadherin (13A9, Santa Cruz Biotechnology, Santa Cruz, CA, USA), Snail (H-130, Santa Cruz Biotechnology), p65 (C-20, Santa Cruz Biotechnology), proliferating cell nuclear antigen (PCNA) (GeneTex, Irvine, CA, USA) or monoclonal Abs against β-actin (AC-15, Sigma Aldrich). HRP-conjugated goat anti-mouse IgG or anti-rabbit IgG (Jackson, West Grove, PA, USA) was used as the secondary Ab and protein-Ab complexes were visualized by enhanced chemiluminescence system (Amersham). The signals were quantified with ImageJ software.
Migration assays
Cell migration was assessed using a modified Boyden Chamber (Corning Costar, Cambridge, MA, USA). The membrane was coated with gelatin (100 ng/ml) for the migration assay. Serum was added to the lower compartment of the chamber. K1735 cells (1 × 104 cells/well) treated with LPS were added to the upper compartment and incubated for various time points. The filter was then fixed with 100% methanol for 8 min and stained by Giemsa stain solution for 1 h. Cells were counted randomly—three images per well under a microscope. 10
Reporter gene assay
To assess the NF-κB activities in tumor cells treated with various concentration LPS, K1735 cells (2 × 104) were transfected with NF-κBLuc. 9 The luciferase activities of cells were determined 16 h after LPS treatment. K1735 cells (2 × 104) were cultured with or without LPS (5 µg/ml) or polymyxin B (20 ng/ml) for 16 h. The cells were then pre-incubated with 10 µg of neutralizing Ab against mouse TLR4 (MTS510; eBioscience, San Diego, CA, USA), or with control IgG (eBioscience) for 30 min at 25°C. After 16 h, the luciferase activities of the tumor cells were determined.
Histology and immunofluorescence
Mice were sacrificed at d 20 after tumor inoculation, and their lungs and tumors were formalin-fixed, paraffin-embedded and sectioned. Tumor tissues were then processed in 5 -µm sections and stained with hematoxylin/eosin (H& E). For immunofluorescence staining of p65 in the K1735, cells were fixed in 10% formalin, permeabilized with cold acetone, incubated with rabbit anti-mouse NF-κB p65 Ab at room temperature for 40 min, and subsequently incubated with fluorescein-conjugated goat anti-rabbit IgG (KPL). The stained cells were examined under a fluorescence microscope at a magnification of × 400. To detect expression of N-cadherin and ZO-1in K1735 cells, cells were fixed in 10% formalin, permeabilized with cold acetone and incubated with monoclonal Abs against N-cadherin and rabbit-anti-ZO-1, and subsequently incubated with rhodomine-conjugated rabbit anti-mouse IgG (KPL, Guildford, UK) and fluorescein-conjugated goat anti-rabbit IgG (KPL). Nuclei were counterstained with 4’,6-Diamidino-2phenylindole (DAPI).
Experimental lung metastatic models
In the experimental murine model for pulmonary metastasis, mice were injected with K1735 cells (105) prior to incubation with resveratrol (16µg/ml) plus LPS (5µg/ml) or LPS for 16 h via tail vein. In a parallel experiment, mice were injected with 105 cells of K1735 cells via the tail vein and survival of the mice was monitored daily. After 20 d, mice were sacrificed and the lungs were removed, weighed and histologically examined. Growth index (mean area of metastasis/total area) of tumor nodules in the lung was quantified at the microscopic level as previously described.9,11,12
Statistical analysis
The statistical differences in various parameters between different groups were analyzed by one-way ANOVA. The survival analyses were performed using the Kaplan-Meier survival curve and the log-rank test. Any P value less than 0.05 is considered statistically significant.
Results
LPS induces EMT
LPS is well documented to elicit a variety of cellular activations, including cell mobilization.
13
EMT is characterized by the disruption of intercellular junctions, the replacement of apical-basolateral polarity, and the acquisition of migratory and invasive phenotypes. First, LPS-induced EMT in K1735 cells was identified by examining the markers of EMT. LPS inhibited the expression levels of ZO-1. However, LPS increased the expression levels of N-cadherin and Snail levels (Figure 1A). The phenotype was dose- and time-dependent in manner (Figure 1B). Next, the migratory potential of K1735 cells treated with LPS was determined. As shown in Figure 1C, the migration of K1735 cells significantly increased after LPS treatment. Similarly, the phenomenon was dose- and time-dependent in manner (Figure 1D).These results suggest that LPS may induce EMT.
LPS induced EMT. (A) LPS dose-dependently induced the marker of EMT. After exposure to various LPS (0–5μ g/ml) for 16 h, the expression of ZO-1, N-cadherin, and Snail levels in K1735 cells were determined by immunoblot analysis. (B) LPS time-dependently induced the marker of EMT. After exposure to LPS (5 µg/ml) for the indicated time, the expression of ZO-1, N-cadherin, and Snail levels in K1735 cells were determined by immunoblot analysis. Inserted values indicate relative proteins expression in comparison with β-actin. (C) LPS dose-dependently induced cell migration. After exposure to various LPS (0–5 µg/ml) for 16 h, the cell migration was determined by Boyden chamber assays. (D) LPS time-dependently induced cell migration. After exposure to various LPS (5 µg/ml) for the indicated time, cell migration was determined by Boyden chamber assays. Each value represents mean ± SD (n = 4). *P < 0.05; **P < 0.01; ***P < 0.001. Data are representative of three independent experiments.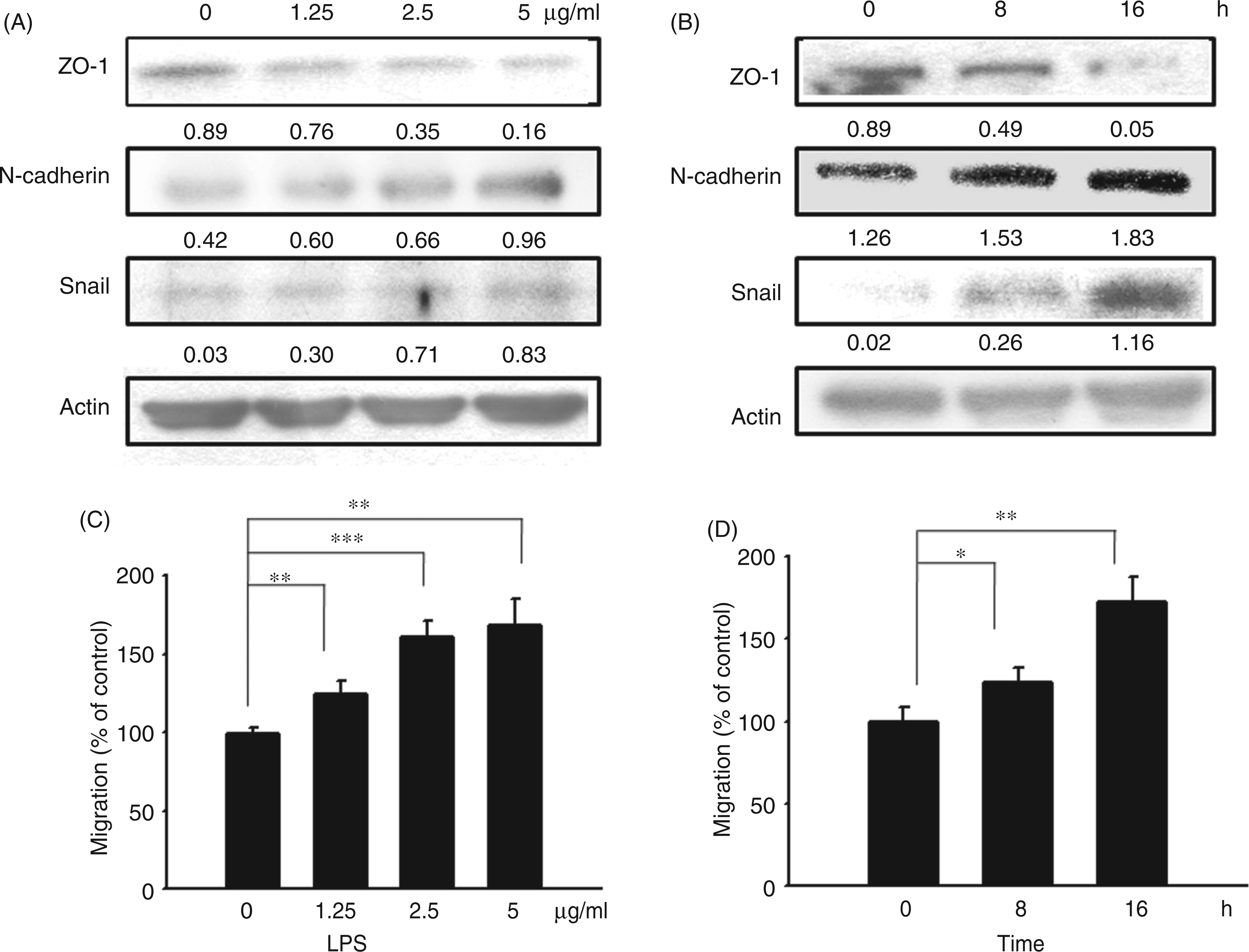
Activation of NF-κB in tumor cells is dependent on TLR4 signaling
NF-κB signaling might play an important role in inflammation and tumor progression.
14
Several NF-κB-regulated genes that encode adhesion molecules, anti-apoptotic proteins and cytokines are essential for tumor growth and migration.
2
We previously found that LPS stimulated tumor growth by activating macrophages via TLR4 signaling.
9
Herein, we investigated whether LPS influenced the NF-κB signaling in tumor cells through TLR4. We performed a reporter assay using K1735 cells transfected with the luciferase gene under the control of NF-κB response elements to assess the activation of the NF-κB signal transduction pathway. The reporter assay revealed that LPS significantly stimulated the NF-κB response in tumor cells. Subsequently, we examined whether LPS mediates the activation of NF-κB signaling via TLR4 on tumor cells by using an Ab-blocking experiment to identify the upstream receptor. By treating with LPS antagonist polymyxin B, it dramatically attenuated the ability of the LPS to induce NF-κB activation in tumor cells. Similar results were found when neutralizing Ab against TLR4 was used to treat the cells (Figure 2B). Some studies implied that resveratrol is unique in its ability to inhibit NF-κB activation.
15
We next examined the effects of resveratrol on the activation of NF-κB in K1735 cells. As shown in Figure 2C, resveratrol inhibited LPS-triggered NF-κB transcriptional activity. Taken together, using reporter assays in vitro, we demonstrated that the LPS stimulated NF-κB transcriptional activities of tumor cells through TLR4 and that resveratrol had the potential to inhibit LPS-induced NF-κB transcriptional activity.
LPS mediates NF-κB activation in tumors. (A) LPS dose-dependently induced NF-κB in tumor cells. The K1735 cells transfected with luciferase gene under the control of NF-κB response elements were treated with LPS (0–5 µg/ml) for 16 h. (B) Activation of NF-κB in tumor cells depends on the functional TLR4 of K1735. The K1735 cells transfected with luciferase gene under the control of NF-κB response elements were treated with LPS (5 µg/ml) or LPS plus polymyxin B (20 ng/ml) for 16 h or pre-incubated with anti-TLR4 or with control IgG. The transcriptional activity of NF-κB was determined by the luciferase reporter assay and is expressed as the percentage of the relative luciferase activity relative to that in the control tumor cells. (C) Resveratrol dose-dependently inhibited the NF-κB activation. The K1735 cells transfected with luciferase gene under the control of NF-κB response elements were treated with LPS (5 µg/ml) plus resveratrol (0-16 µg/ml) for 16 h. The transcriptional activity of NF-κB was determined by the luciferase reporter assay and is expressed as the percentage of the relative luciferase activity relative to that in the control tumor cells. Each value represents mean ± SD (n = 4). *P < 0.05; **P < 0.01; ***P < 0.001. Data are representative of three independent experiments.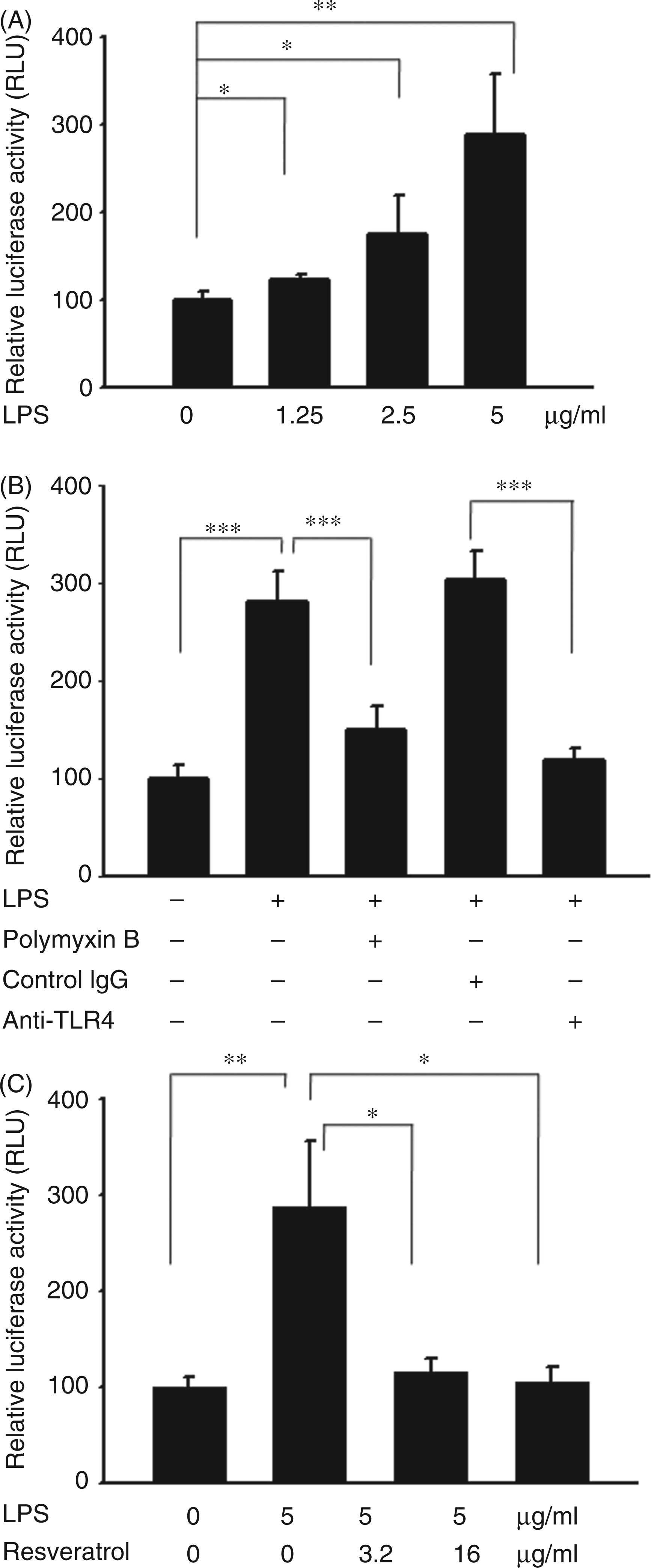
Resveratrol inhibited LPS-induced EMT
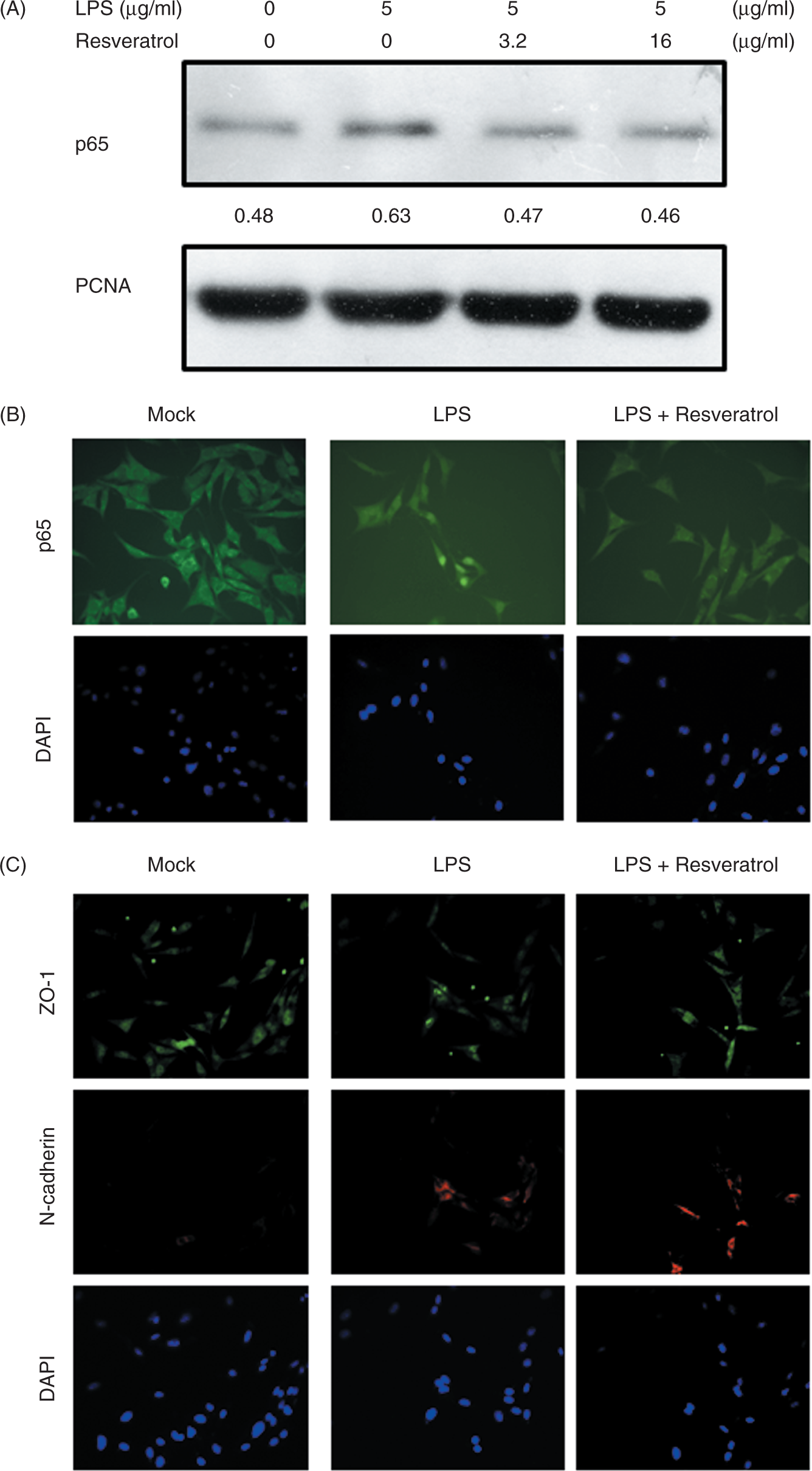
Induction of EMT in melanoma by LPS was performed according to the method as previously described. Resveratrol suppressed LPS-induced EMT, as determined by the expression levels of marker proteins of EMT (Figure 3A). Furthermore, the LPS-induced cell migration can also be inhibited by resveratrol (Figure 3B). Because LPS caused the activation of NF-κB in tumor cells via TLR4 signaling (Figure 2B), we further examined the subcellular localization of p65 subunit of NF-κB in tumor cells after LPS treatment by immunoblot assay and immunofluorescence staining. Resveratrol inhibited LPS-induced nuclear translocation of the p65 subunit of NF-κB (Figure 4A). Meanwhile, p65 was expressed only modestly at the nuclear periphery and in the cytoplasm of the control cells, whereas it was detected exclusively in the nucleus of the cells treated with LPS by immunofluorescence staining. The phenotype was reversed in cells by treating with resveratrol (Figure 4A and B). Analysis by immunofluorescence showed that treatment of K1735 cells with LPS caused a complete EMT phenotype (loss of ZO-1 expression and massive up-regulation of N-cadherin). Interestingly, resveratrol reverted to LPS-induced EMT, causing re-induction of ZO-1 and inhibition of N-cadherin expression (Figure 4C). The results suggest that resveratrol inhibits NF-κB transcriptional activity and suppresses EMT.
Resveratrol inhibited LPS-induced EMT. (A) Resveratrol dose-dependently inhibited the marker of EMT. The K1735 cells were stimulated with LPS (5 µg/ml) plus resveratrol (0-16 µg/ml) for 16 h. The expression of ZO-1, N-cadherin and Snail levels in K1735 cells were determined by immunoblot analysis. Inserted values indicated relative proteins expression in comparison with β-actin. (B) Resveratrol dose-dependently inhibited tumor cell migration. After exposure to LPS (5 µg/ml) plus resveratrol (0-16 µg/ml) for 16 h, cell migration was determined by Boyden chamber assays. Each value represents mean ± SD (n = 4). **P < 0.01. Data are representative of three independent experiments. Resveratrol inhibited LPS-induced NF-κB activation and EMT. (A) Resveratrol inhibited LPS-induced nuclear translocation of the p65 subunit of NF-κB. The K1735 cells were stimulated with LPS (5 µg/ml) plus resveratrol (0–16 µg/ml) for 16 h. The expression of p65 levels in nuclei were determined by immunoblot analysis. PCNA expression served as loading control for nuclear proteins. Inserted values indicated relative proteins expression in comparison with PCNA. (B) Resveratrol inhibited LPS-induced NF-κB activation. The K1735 cells were treated with or without LPS (5 µg/ml) plus resveratrol (16 µg/ml) for 16 h. The location of p65 in K1735 cells were determined by immunofluorescence staining. (C) Resveratrol inhibited LPS-induced EMT. The K1735 cells were treated with or without LPS (5 µg/ml) plus resveratrol (16 µg/ml) for 16 h. The expression of ZO-1 and N-cadherin in K1735 cells were determined by immunofluorescence staining. Nuclei were counterstained with DAPI. Data are representative of three independent experiments.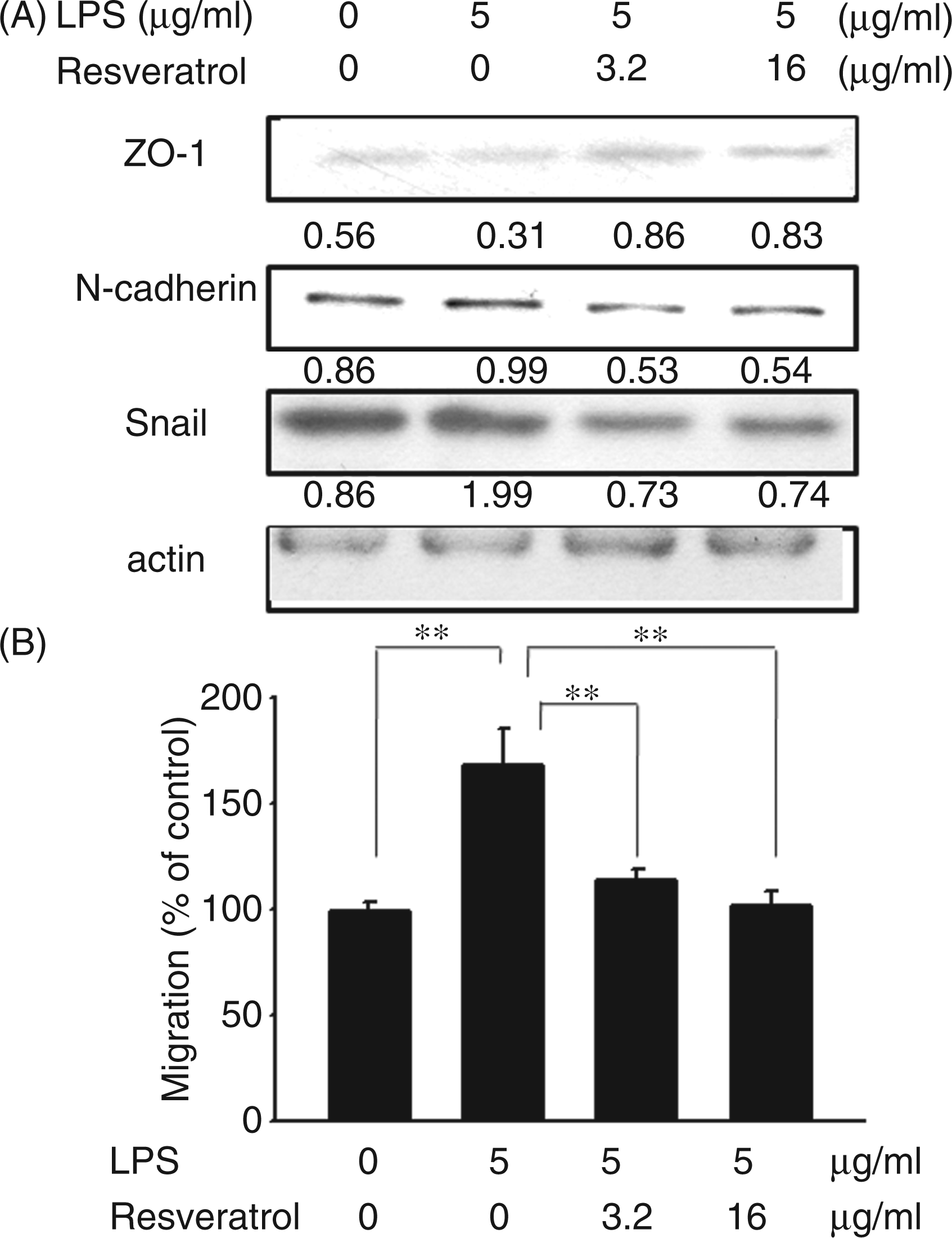
Resveratrol inhibited LPS-induced metastasis
Previously, we found that mice injected with tumor cells admixed with LPS had a significantly shorter survival time compared with those injected with only tumor cells.
9
We next determined whether resveratrol could inhibit LPS-induced metastasis. The tumor cells pre-incubated with LPS or LPS plus resveratrol were injected into mice via tail vein. Histologic examination confirmed the macroscopic findings (Figure 5A). Consistent with a previous report,2,5,9 our results suggest that LPS influenced the tumor mass in lung (Figure 5B) and the survival of tumor-bearing mice (Figure 5C). The lower tumor burden correlated with longer survival time in mice injected with cells treated with LPS and resveratrol compared with mice injected with cells incubated with LPS. Quantitative analysis at the microscopic level also shows that the growth index of tumor nodules was significantly lower in mice injected with cells treated with LPS and resveratrol than in the mice injected with cells incubated with LPS (mean ± SD 0.6783 ± 0.31 vs 1.31 ± 0.41, n = 8; P < 0.05). The growth index of tumor nodules was not different between mice injected with cells and mice cells treated with LPS and resveratrol (mean ± SD 0.6892 ± 0.22 vs 0.6783 ± 0.31, n = 8). Taken together, these results suggest that resveratrol affects EMT.
Resveratrol inhibited LPS-induced metastasis in vivo. Groups of nine C3H/HeN mice were inoculated with K1735 cells (105) pre-incubated with or without LPS (5 µg/ml) or LPS plus resveratrol (16 µg/ml) for 16 h via the tail vein. (A) Representative examples of metastatic pulmonary nodules produced 20 d after intravenous injection of 105 K1735 cells. Histologic examination of pulmonary tumor nodules at d 20 after tumor inoculation. Note that H&E staining of 4 -µm paraffin-embedded lung tissue sections (40 ×) reveals the presence of larger tumor nodules from mice injected LPS-treated-K1735 cells compared with those from mice injected with LPS plus resveratrol-treated K1735 cells. (B) At d 20, the wet lung weight of the lungs was measured. Each value represents mean ± SD (n = 8). (C) Kaplan-Meier survival curves are shown (n = 9). *P < 0.05; **P < 0.01. (P < 0.05 for K1735 versus K1735 + LPS; P < 0.01 for K1735 + LPS versus K1735 + LPS + Resveratrol). Data are representative of two independent experiments.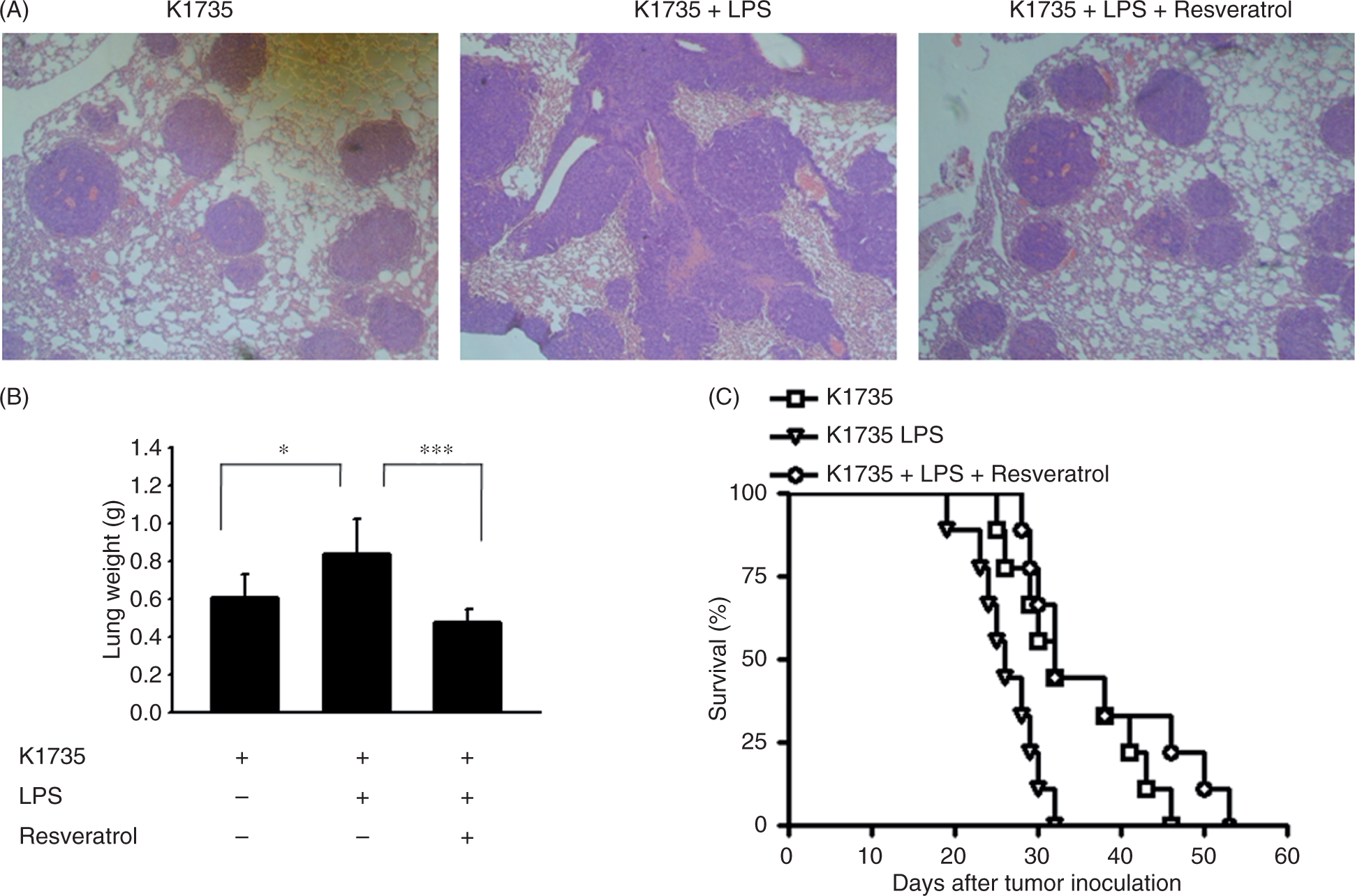
Discussion
In this study, we demonstrated that resveratrol inhibited LPS-mediated cell migration in melanoma. These findings provide important insights into the molecular mechanisms of grape seed extract in cancer prevention through suppression of tumor migration. Our results indicated that LPS acted as a stimulant for cellular migration. NF-κB activation by LPS induced the EMT phenotype with up-regulation of N-cadherin and Snail, and down-regulation of ZO-1. These results imply, by virtue of reduction NF-κB activities, that resveratrol could effectively hamper LPS-induced the EMT in melanoma.
The expression of TLR4 on tumor cells was reported to play a role in immune surveillance and facilitate tumor growth and chemoresistance. 16 TLR4 initiates signals through the sequential recruitment of MyD 88, which, in turn, activates downstream mediators, such as NF-κB, leading to the activation of genes encoding proinflammatory cytokines. The MyD88 signaling pathway downstream of TLR4 was demonstrated to induce hepatocarcinogenesis and intestinal tumorigenesis.17,18 TLR4-MyD88 signaling may function upstream of NF-κB in cells involved in inflammation-associated cancer and NF-κB is a multifunctional transcription factor that affects tumor growth and metastasis. 19 Increases in the NF-κB activity in the tumor microenvironment result in chronic inflammation and a substantial pro-tumorigenic effect. Herein, we demonstrated that LPS stimulated the activation of NF-κB in tumor cells and enhanced the EMT phenotype.
Many studies have demonstrated that resveratrol has a wide range of pharmacologic properties. Resveratrol exerts an anti-inflammatory response via inhibition of NF-κB. 20 Park et al. have suggested that the inhibition of the LPS-induced cell adhesion molecules by resveratrol was mediated by NF-κB. 21 Resveratrol inhibited LPS-stimulated nuclear translocation of the p65 subunit of NF-κB in endothelial cells. Resveratrol acts on NF-κB by the inhibition of IκB 8 which results in the prevention of translocation p65 into the nucleus or deacetylation of NF-κB. 20 Resveratrol may play an important role in anti-inflammation and targets for the NF-κB signaling pathway could be used as effective therapies for inflammatory-mediated disease, such as cancer.
Expression of Snail represses expression of E-cadherin and induces EMT in breast cancer cells, indicating that Snail plays a fundamental role in EMT and metastasis. 22 In addition, over-expression of Snail correlates with tumor grade, metastasis and tumor recurrence in patients.23,24 Tumor migration and invasion at the tumor-host boundary are induced by inflammation through Snail-mediated EMT induction. 5 Hererin, we found that LPS induced the expression of Snail in melanoma (Figure 1). Previous studies suggested that the activation of the NF-κB pathway is important for Snail stabilization. 5 In our study, we found that LPS may enhance the migration of tumor cells by inducing EMT programming through NF-κB-mediated Snail stabilization. We further demonstrated that blocking of the NF-κB pathway reverted LPS-induced metastasis by resveratrol treatment (Figure 3). Inflammation promotes tumor growth and metastasis; the benefit of resveratrol prevents metastasis through its anti-inflammation effects.
Conclusions
These data suggest that resveratrol inhibits EMT by inhibiting NF-κB signaling. In conclusion, resveratrol may be used as a therapeutic agent in the treatment of melanoma.
Footnotes
Funding
The authors wish to acknowledge support relevant to this manuscript from the National Science Council (NSC 100-2320-B-039-024) and China Medical University (CMU-99-N2-08). They had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Acknowledgements
We thank Dr MC Hung (The University of Texas M. D. Anderson Cancer Center) for generously providing K1735 cells.
Conflicts of interest
The authors declare that they have no conflicts of interests.