The interaction of lipopolysaccharide-primed murine peritoneal macrophages with ivermectin, an antiparasite drug which potentiates P2X4 receptors and dynasore which inhibits the GTPase activity of dynamin, a protein contributing to the internalization of plasma membrane proteins, was tested. Murine peritoneal macrophages express P2X4 receptors which are mostly intracellular. In cells from P2X7-knockout mice (KO mice), 10 µm adenosine triphosphate (ATP) provoked a transient increase of the intracellular concentration of calcium. Ivermectin had no effect by itself but potentiated the increase of the intracellular concentration of calcium by ATP. The combination of ATP plus ivermectin also decreased the intracellular concentration of potassium and promoted the secretion of IL-1β. Concentrations of dynasore above 50 µm affected the integrity of mitochondria (MTT test) and of the plasma membrane (release of lactate dehydrogenase, LDH). At a 10 µm concentration, dynasore had no effect on the responses to ATP and on the internalization of P2X4 receptors. By itself dynasore promoted the release of potassium and the secretion of IL-1β after activation of caspase-1. In conclusion, our results confirm that ivermectin potentiates the responses coupled to P2X4 receptors probably by interaction with an allosteric site. We also show that this potentiation triggers the release of IL-1β by macrophages. As opposed to ivermectin, dynasore has no effect on P2X4 receptors. This drug triggers a potassium efflux via a mechanism which does not involve purinergic receptors and generates, in consequence, the activation of caspase-1 and the secretion of IL-1β.
P2X receptors are plasma membrane proteins with two transmembrane domains. After binding nucleotides or nucleosides, they assemble to homo- or hetero-oligomeric structures which form a non-selective cation channel.1 Seven ionotropic purinergic receptors have been cloned. They differ with respect to their selectivity for various agonists or antagonists, or to their rate of desensitization, or to their tissular expression.2 The P2X7 receptors are the most different of these receptors. They have a long C-terminal domain, enabling this receptor to interact with various intracellular proteins.3 The P2X7 receptor has a very low affinity for adenosine triphosphate (ATP) and after sustained activation its permeability switches from a channel to a pore permeant to ions like ethidium or propidium.4 It is mostly expressed by immunocytes, but there is also large consensus for its expression by exocrine glands.5 In the murine and human genomes, the genes coding for P2X7 and P2X4 receptors are in tandem loci.6 The P2X4 receptor is usually co-expressed with P2X7 receptors by macrophages and nervous or glandular tissues and, apparently, the P2X4 receptor is the only P2X subunit able to form a heterotrimer with the P2X7 receptor.7 The lack of specific agonists or antagonists for P2X4 receptors and their rapid desensitization have hampered the physiological characterization of the P2X4 receptors. Very few P2X4 receptors are inserted in the plasma membrane. Most of them have an intracellular location (probably on the lysosomal membrane) where their role has not yet been determined.8 The C-terminal domain of the receptor contains a non-canonical tyrosine-based endocytic motif which interacts with the µ2 subunit of the adaptor protein AP2.9 This heterotetramer serves not only as a cargo for sorted proteins but also contributes to the coating of fragments of the membrane with polymerized clathrin.10 Dynamin, a GTPase, contributes to the pinching of these zones and the release of these clathrin-coated vesicles into the cytosol where they will eventually fuse with lysosomes.11 The transfer of P2X4 receptors is inhibited in cells expressing a dominant-negative mutant form of dynamin-1, confirming the role of dynamin in the internalization of these receptors.12 Brown and Yule recently reported that this pathway could be indirectly regulated by cyclic adenosine monophosphate (cAMP).13 According to these authors a protein phosphorylated by protein kinase A (maybe one component of AP2) blocks the internalization of the receptor. This inhibition accounts for the potentiation by cAMP of the responses coupled to P2X4 receptors. The internalization of P2X4 receptors via a clathrin-coated vesicle proceeds via a mechanism similar to other ionotropic receptors like the AMPA and GABAA receptors.14
Dynasore, a non-competitive inhibitor of the GTPase activity of dynamin, was isolated from 16,000 compounds.15 Exposure of eukaryotic cells to this experimental drug fully blocks the intracellular transfer of plasma membrane proteins mediated by clathrin.16 Dynasore has also been shown to potentiate the responses coupled to P2X4 receptors in microglia, but not in bone marrow derived macrophages (BMDM).17 In this study we compared the response of murine peritoneal macrophages coupled to P2X4 receptors after their pretreatment with either ivermectin or dynasore. The purpose of these experiments was to determine whether the responses coupled to these receptors were regulated by their internalization. To avoid any contribution of P2X7 receptors, the experiments were performed on macrophages isolated from P2X7-KO mice.18 We have previously reported that these cells do not express any functional P2X7 receptors but respond to P2X4 stimulation.19 In contrast to ivermectin, we show that a low, non-deleterious concentration of dynasore did not potentiate the response to a low (10 µm) concentration of ATP. We also show that by itself, dynasore did not modify the intracellular concentration of calcium ([Ca2+]i), but decreased the intracellular concentration of potassium ([K+]i). The effect of dynasore on the monovalent cation remained unexplained. It probably accounted for the activation of caspase-1 and the release of IL-1β in response to the drug and should be considered when studying the effect of dynasore on the responses of macrophages to various stimuli.
Materials and methods
Materials
Ethylene glycol-bis-(β-aminoethyl ether)-N,N,N’,N’-tetraacetic acid (EGTA), N-[2-hydroxyethyl] piperazine-N’-[2-ethanesulfonic acid] (HEPES), ivermectin, dynasore, nigericin, ATP disodium salt, fetal bovine serum (FBS), lipopolysaccharides (LPS) from Escherichia coli (Sigma catalog ref 2880), Tween-20, Triton X-100 the protease inhibitor cocktail, β-nicotinamide adenine dinucleotide (NADH), sodium pyruvate and 3-(4,5-dimethylthiazole-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) were obtained from Sigma-Aldrich (St Louis, MO, USA). Bovine serum albumin (BSA) fraction V was obtained from Roche Diagnostics (Mannheim, Germany). Trishydroxymethylaminomethane (Tris) was obtained from VWR (Leuven, Belgium). Fura2-AM, PBFI-AM, the glutamine-free amino acid mixture, phosphate-buffered saline (PBS), RPMI-1640, L-glutamine, the solution of penicillin-streptomycin, the gels (NuPAGE Novex Bis-Tris 4-12% and 12% gels), the various buffers for electrophoresis (NuPAGE LDS Sample Buffer; NuPAGE MOPS SDS Running Buffer; NuPAGE Transfer Buffer), the nitrocellulose and polyvinylidene fluoride (PVDF) membranes and the solution of molecular mass standards for the electrophoresis (SeeBlue Plus 2) were all obtained from Invitrogen (Groningen, the Netherlands). Thioglycollate was supplied by Becton-Dickinson (Franklin Lanes, NJ, USA). The biotinylated monoclonal anti-mouse IL-1β antibody and the mouse IL-1β were obtained from Endogen (Perbio Science, Erembodegem, Belgium). The polyclonal anti-mouse IL-1β antibody was from R&D Systems (Minneapolis, MN, USA). The HRP substrate 1 Step Ultra TMB-ELISA, the peroxidase-linked streptavidin, the sulfo-NHS-SS-biotin and the streptavidin agarose resins were bought from Pierce (Perbio Science, Erembodegem, Belgium). The polyclonal anti-mouse caspase-1 p10 antibody was purchased from Santa Cruz (Santa Cruz, CA, USA). Anti-P2X4 antibody was purchased from Alomone (Jerusalem, Israel). The chemiluminescent ECL Plus reagent and the peroxidase-linked anti-rabbit IgG were all obtained from GE Healthcare (Buckinghamshire, UK).
The experiments were carried out on P2X7R-/- null mice (KO) initially supplied by Pfizer Inc. (Groton, CT, USA). Breeding of P2X7 knockout mice males with females was used to maintain the colony of receptor-deficient animals. Mice used in the experiments were between 20–25 g. The lack of expression of P2X7 receptors in KO mice was tested by Western blot as previously described.20
Methods
Cell culture
Adult male P2X7-KO mice were used for these experiments. Peritoneal macrophages were prepared as described previously.21 Briefly, peritoneal exsudates were induced by injection of 1.5 ml thioglycollate solution (4% in sterile water) 3 or 4 days before harvesting the cells. The animals were fasted overnight and sacrificed by raising the CO2 concentration in accordance with the procedures of the Belgian Ministry of Agriculture under the supervision of the institutional ethical committee (protocol n° 221N). Ten milliliters of sterile PBS without calcium or magnesium (pH 7.2), containing heparin (10U/ml), were injected into the peritoneal cavity. The abdomen was gently massaged after its distension. The fluid was recovered and transferred to a sterile tube kept on ice. After centrifugation at 1500 g for 10 min at 4°C, the cellular pellet was resuspended in RPMI 1640 medium supplemented with 20 mm HEPES, 10% FBS, 2 mml-glutamine, 100 U/ml penicillin and 100 µg/ml streptomycin (RPMI medium). Ten million cells were added to culture flasks (usually two flasks per mouse) and incubated at 37°C for at least 2 h in a humidified atmosphere containing 5% CO2. Non-adherent cells and the medium were then aspirated and replaced with 10 ml fresh RPMI medium. The cells were incubated overnight at 37°C in the presence of 250 ng/ml lipopolysaccharides (LPS) before use.
Determination of the [Ca2+]i
The culture plates were washed twice with 10 ml PBS. The cells were gently scraped off the plates with a rubber policeman. They were incubated at 25°C for 45 min in a HEPES-buffered medium (HBM) containing: 24.5 mm HEPES (pH 7.4), 96 mm NaCl, 6 mm KCl, 2.5 mm NaH2PO4, 11.5 mm glucose, 5 mm sodium pyruvate, 5 mm sodium glutamate and 5 mm sodium fumarate. This medium also contained 1 mm CaCl2 and MgCl2, 1% (v/v) glutamine-free amino acid mixture, 1% (w/v) BSA (medium HBT) and 3 µm fura2-AM. At the end of this incubation, the cells were washed and diluted with HBM supplemented with 1 mm CaCl2. They were transferred to the cuvette of a fluorimeter and maintained at 25°C under constant stirring. The light emitted at 510 nm was measured every second after excitation at 340 and 390 nm (Photon Technology International, Birmingham, NJ, USA). At the end of the incubation, the cells were permeabilized with 100 µm digitonin and the autofluorescence of the samples was estimated by quenching fura2 with 100 mm manganese. The blank values were subtracted from the results and the [Ca2+]i was estimated as described.22
Determination of the [K+]i
Macrophages were incubated at 25°C for at least 1 h in HBT in the presence of 3 µm PBFI-AM. At the end of this incubation, the cells were washed and diluted with HBM medium with 1 mm CaCl2. They were transferred to the cuvette of a fluorimeter and maintained at 25°C under constant stirring. The light emitted at 490 nm (slitwidth 16 nm) was measured every second after excitation at 345 nm (slitwidth 16 nm) (Photon Technology International). Results were plotted as percentage of the initial fluorescence without any attempt to calibrate the traces.
Assay of IL-1β secretion
Peritoneal macrophages from one mouse were dispersed in RPMI medium at a 500,000 cells/ml dilution and 1-ml aliquots were transferred to a 12-well plate. After 3 h at 37°C, the medium was removed and the cells were incubated overnight in fresh RPMI medium containing 250 ng/ml LPS. The cells were washed twice with PBS. They were then incubated for 15 min at 37°C in 1 ml HBM medium without magnesium but containing 1 mm CaCl2, 0.1% BSA and the tested agent. Each condition was tested in duplicates. Incubation medium was collected, centrifuged at 10,000 g for 5 min to remove detached cells and the supernatant sonicated for 15 sec at 15 µm amplitude (Soniprep 150, MSE, London, UK). The IL-1β content was assayed by sandwich enzyme-linked immunosorbent assay (ELISA). A 96-well plate was coated overnight with 1 µg/ml polyclonal anti-mouse IL-1β antibody and blocked with 4% BSA in PBS for 1 h at 25°C. The wells were washed twice with Tris-Tween buffer (Tris-HCl 50 mm, pH 7.5 and 0.2% Tween-20). Aliquots of medium samples or recombinant IL-1β standards were diluted to 150 µl with HBS medium containing 1 mm CaCl2 and 0.1% BSA and mixed with 150 µl biotinylated monoclonal anti-mouse IL-1β antibody (0.4 µg/ml solution). One hundred microliters of this mix were added in duplicate to the wells. Within each experiment, each condition was tested in duplicates. The plate was incubated at 37°C for 2 h and then washed four times. The captured immune complexes were incubated for 30 min at 25°C with 100 µl streptavidin-HRP (0.2 µg/ml), washed four times and colorimetrically developed with 100 µl HRP substrate. After the addition of 100 µl stop solution (H2SO4 1.8 m), the absorbance was read at 450 nm in a Synergy HT plate reader (Bio-Tek, Winooski, VT, USA).
Western blot analysis of P2X4 receptors
Peritoneal macrophages from one mouse were dispersed in RPMI medium at a density of 2.0 × 106 cells/ml and 1-ml aliquots were transferred to a tissue culture dish (Nunclon 10x30; NUNC®, Rochester, NY, USA). After 3 h at 37°C, the medium was removed and the cells were incubated overnight in fresh RPMI medium containing 250 ng/ml LPS. The cells were washed twice with PBS before their incubation at room temperature for 5 min with either 3 µm ivermectin or 10 µm dynasore or diluted dimethylsulfoxide (DMSO) (control condition) followed by 15 min incubation with the association tested agent/sulfo-NHS-SS-biotin (freshly prepared; 1 mg/ml in PBS) and by 20 min incubation with sulfo-NHS-SS-biotin alone. Excess biotin was removed by quenching the unlinked biotin with 50 mm glycine made up in PBS and by washing the cells twice with cold PBS. Adherent cells were solubilised with lysis buffer (50 mm Tris HCl, 150 mm NaCl, 0.5% Na deoxycholate, 0.1% SDS and 0.1% Igepal CA-630; pH 8) containing a cocktail of protease inhibitors. The resulting cellular suspensions were incubated on ice for 30 min before sonication and clarifying centrifugation. Thirty µl of the supernatants were mixed with 10 µl LDS sample buffer and dithiothreitol (DTT) (50 mm) and kept at −80°C in order to assess total P2X4 receptors for each sample (‘homogenate fractions’). Another aliquot of the supernatants (100 µl) was incubated with immobilised streptavidin-coupled biotin-binding beads on a rotating rack at 4°C in order to extract biotinylated proteins. Eighteen hours later, biotinylated proteins linked to beads were spun for 1 min at 15,000 g at 4°C and the supernatants were removed. After washing the beads three times with 1 ml lysis buffer, the proteins were eluted from the beads by incubation in 40 µl LDS sample buffer containing 50 mm DTT (‘surface fractions’). Twenty µl of the homogenate and of the surface fractions were separated by SDS-PAGE performed on NuPAGE® 4–12% Bis-Tris gel, transferred onto a nitrocellulose membrane and detected by immunoblotting using an anti-P2X4 primary antibody from rabbit, a secondary anti-rabbit antibody coupled to peroxidase and a chemiluminescent substrate (ECL Plus).
Western blot analysis of caspase-1 cleavage
Peritoneal macrophages were plated in 12-well plates at a density of 1.5 × 106 cells per well. They were primed overnight in RPMI medium containing 250 ng/ml LPS. They were then stimulated for 15 min at 37°C in HBM medium without magnesium but with 1% (v/v) glutamine-free amino acid mixture, 0.05% BSA and 1 mm CaCl2, either in control condition or in the presence of dynasore (10 µm). The incubation medium was aspirated and the cells were incubated with a lysis buffer (25 mm HEPES, 300 mm NaCl, 1.5 mm MgCl2, 0.2 mm EDTA and 0.1% Triton X-100) containing a protease-inhibitor cocktail. Cell lysates were then collected and centrifuged at 1500 g for 10 min at 4°C to remove cell debris. The resulting supernatants were mixed with LDS sample buffer containing mercaptoethanol (5%) and were heated at 80°C for 1 h. Protein concentrations were determined using the BCA Protein Assay Kit (Pierce, Rockford, IL, USA), and equal concentrations of proteins were loaded for SDS-PAGE on a NuPAGE® 12% Bis-Tris gel. Proteins were then transferred to a PVDF membrane. The membrane was incubated overnight with a polyclonal anti-mouse caspase-1 antibody from rabbit. A secondary anti-rabbit antibody coupled to peroxidase and a chemiluminescent substrate (ECL Plus) were used for revelation.
Assay of the release of lactate dehydrogenase (LDH)
The LPS-primed macrophages were resuspended in HBM medium supplemented with 0.1% BSA, 1% (v/v) glutamine-free amino acids and 1 mm CaCl2, in the absence of magnesium. They were incubated for 1 h at 37°C under constant agitation both in control and in the tested condition. Each condition was tested in triplicate. At the end of the incubation the cellular suspension was centrifuged for 5 min at 10 000 g. To estimate the amount of LDH present in the medium at the beginning of the incubation (blank values), a similar procedure was applied to non-incubated aliquots of cells. Aliquots of cells were also sonicated to estimate the total LDH activity. The assay was performed in a 96-well microplate. Ten microliters of the enzyme solution (cellular supernatant) were transferred to the wells. Then, 190 µl Tris-EGTA-NADH (56 mm Tris, 5.6 mm EGTA and 170 µm β-NADH) and 20 µl sodium pyruvate (14 mm) were added to the wells. The oxidation of NADH was monitored at 340 nm for 20 min at 30°C (Synergy HT, BioTek, Winooski, VT, USA). Absorbance measurements were performed at time intervals of 45 sec. The decrease in absorbance per minute was estimated by linear regression and the blank value was subtracted from the results obtained with the incubated cells and the cellular homogenate.
MTT test
Peritoneal macrophages from one mouse were dispersed in RPMI medium at a 500,000 cells/ml dilution and 200 µl aliquots were transferred in a 96-well plate. Following overnight incubation in fresh RPMI medium containing 250 ng/ml LPS, the cells were washed twice with PBS. They were then incubated for 1 h at 37°C in the presence of various concentrations of dynasore. Each condition was tested in sextuplates. The effect of dynasore on the viability of the cells was assessed by measuring the ability of cells to reduce 3-(4,5-dimethylthiazole-2-yl)-2,5-diphenyl tetrazolium bromide (MTT).23 This reaction is catalyzed by oxidoreductases which use electrons from mitochondria-generated NADH as the primary reductants. After incubation of the cells with MTT for 3-4 h, formazan crystals develop in living and early apoptotic cells. These crystals are then solubilized with DMSO and the absorbance measured at 540 nm (Synergy HT, BioTek).
Statistical analysis
Results were analyzed using the Mann-Whitney non-parametric test. ***P < 0.005; **P < 0.01; *P < 0.05.
Results
Effect of dynasore on the viability of peritoneal macrophages
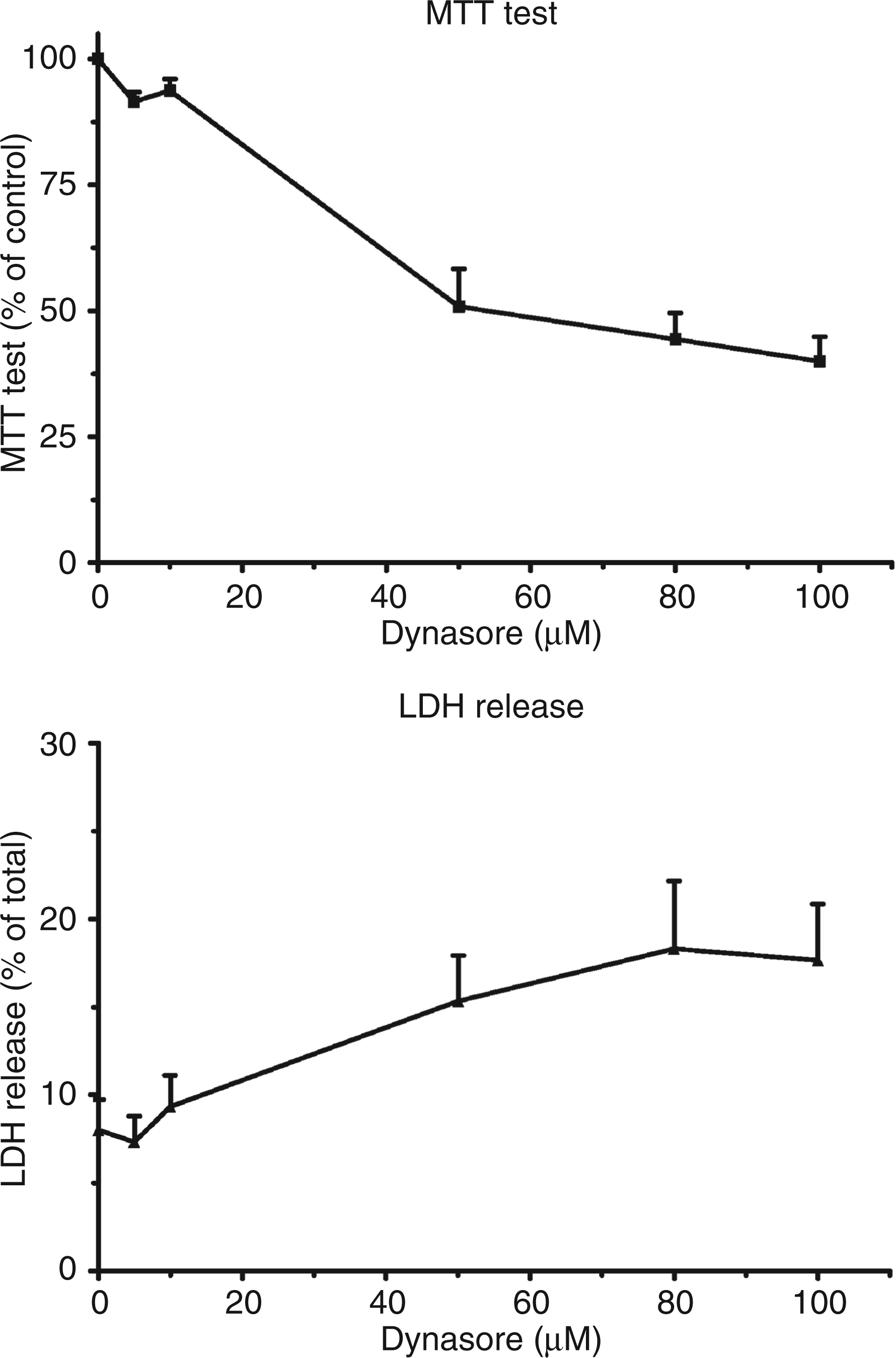
Taking into account that P2X4 receptors are located not only on the plasma membrane but also on intracellular membranes, we considered the possible contribution of dynamin, a GTPase involved in the internalization of vesicles coated with clathrin, to the trafficking of the receptors between these two compartments. To this end, we tested the response of macrophages to a low concentration of ATP when dynasore, a drug which inhibits dynamin, had been added to the medium. Initially, the toxicity of the drug on murine peritoneal macrophages was studied. The integrity of the mitochondrial function was evaluated with the MTT test while the release of LDH in the medium was an index of the leakiness of the plasma membrane. Concentrations of dynasore between 0 and 100 µm were tested. As shown in the upper panel of Figure 1, the drug dose-dependently affected the activity of mitochondria. The absorbance of the dye in the MTT test dropped by 50% at a concentration of dynasore between 10 and 50 µm. Similar results were obtained when measuring the release of LDH in the medium: at a 10 µm concentration dynasore did not significantly increase the activity of LDH in the incubation medium (Figure 1, lower panel, less than 10% total activity). Raising the concentration of dynasore to 50 µm doubled the release of LDH. From these results it was decided to test dynasore at a 10 µm concentration.
Study of the toxic effects of dynasore on LPS-primed murine peritoneal macrophages. LPS-primed macrophages from P2X7-knockout (KO) mice were incubated in the presence of various concentrations of dynasore for 1 h at 37°C. At the end of the experiment an MTT test (upper panel) was performed on the cells and the activity of the lactate dehydrogenase (LDH) released in the medium (lower panel) was measured. Results are the means + SEM of six experiments (MTT test) and of three experiments (LDH release).
Expression and localisation of P2X4 receptors in peritoneal macrophages
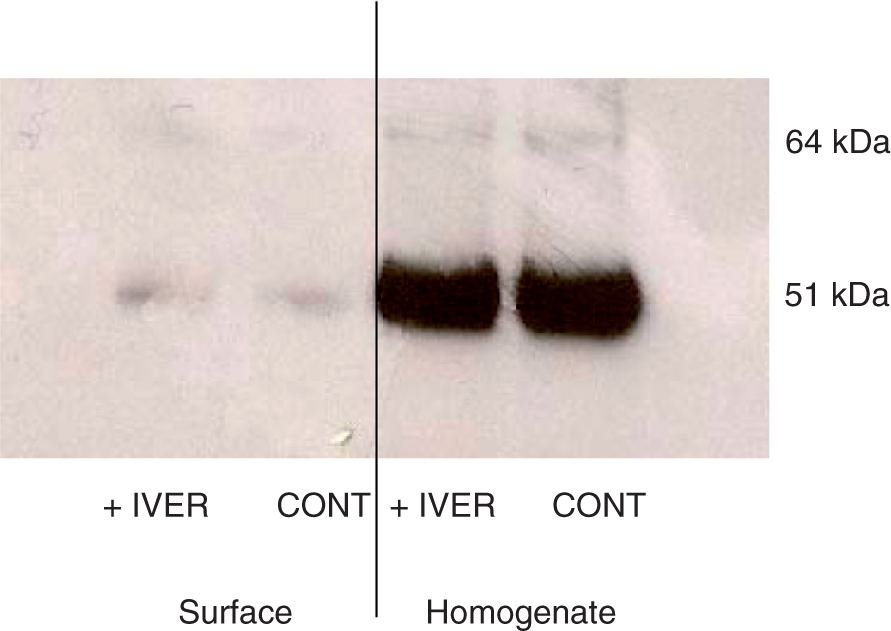
The expression of P2X4 receptors by macrophages and their location in the cells were examined next. The effect of dynasore was compared with the effect of ivermectin, a drug which potentiates the responses coupled to P2X4 receptors.24 LPS-primed macrophages were incubated for 10 min in control conditions or in the presence of 3 µm ivermectin or of 10 µm dynasore. Results obtained with cells incubated in control conditions and in the presence of ivermectin have been plotted in Figure 2. The strong signal observed after incubation of the homogenate fraction with anti-P2X4 antibodies confirmed that macrophages expressed the P2X4 receptor. In the surface fraction, the anti-P2X4 antibody reacted slightly with a protein of 50–60 kDa. The incubation of the macrophages with ivermectin did not increase the intensity of this band suggesting that the drug did not modify the location of the receptors (Figure 2). Similar negative results were obtained with 10 µm dynasore (data not shown). From these results it could be concluded that very few P2X4 receptors were inserted in the plasma membrane and that neither ivermectin nor dynasore increased the fraction of the receptors exposed at the surface of the macrophages.
Cellular localization of P2X4 receptors in LPS-primed murine peritoneal macrophages. LPS-primed macrophages from P2X7-knockout (KO) mice were incubated for 10 min in control conditions (CONT) or in the presence of 3 µm ivermectin (IVER). Proteins of the plasma membranes were covalently labelled with biotin as described in the Materials and Methods section. An immunoblot was performed either on 20 µl of total cellular extract (Homogenate) or on 20 µl solution of biotinylated proteins (Surface). This figure is representative of three experiments.
Interaction of dynasore and ATP on ion movements
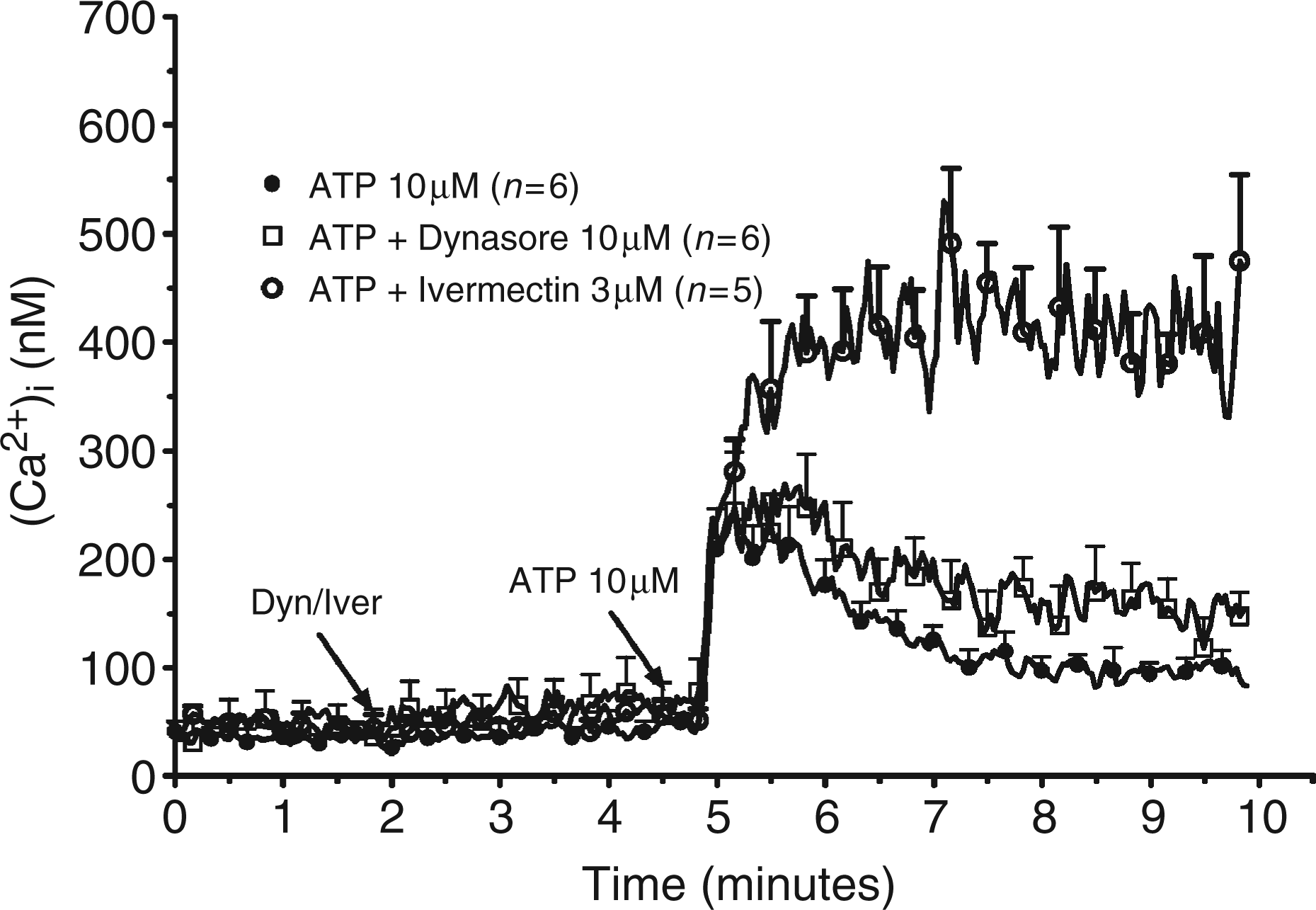
Exposure of the peritoneal macrophages of P2X7-KO mice to 10 µm ATP provoked a transient increase of the [Ca2+]i through activation of P2X4 receptors.19 The concentration of the ion peaked after 20 sec (from 41 ± 10 nm to 210 ± 37 nm, n = 6, Figure 3) and decreased at later times (down to 97 ± 13 nm after 3 min). The contribution of P2X4 receptors to the regulation of the [Ca2+]i was confirmed with the preincubation of the cells with ivermectin. By itself, 3 µm ivermectin had no effect on the [Ca2+]i. It strongly potentiated both the amplitude and the duration of the response to ATP: the [Ca2+]i reached 356 ± 62 nm (n = 5) after 20 sec and did not decrease at longer times (411 ± 56 nm after 3 min). This result confirmed that murine peritoneal macrophages express functional P2X4 receptors which desensitize rapidly and which are sensitive to regulation by ivermectin. Considering that ivermectin did not increase the fraction of receptors located in the plasma membrane (vide supra), our results confirmed that the potentiation exerted by ivermectin was secondary to an allosteric effect rather than inhibition of internalization. Preincubation of the cells with 10 µm dynasore did not modify the response to ATP. In the presence of the drug the [Ca2+]i increased to 244 ± 54 nm after 20 sec and decreased to 135 ± 22 nm after 3 min (n = 6).
Effect of dynasore on the [Ca2+]i of LPS-primed murine peritoneal macrophages. LPS-primed macrophages from P2X7-knockout (KO) mice were loaded with fura2 for 45 min at 37°C. After washing, the cells were incubated at 37°C in the cuvette of a fluorimeter under constant stirring. Two minutes after the start of the measurement the cells were either maintained in control conditions or exposed to 10 µm dynasore (Dyn) or to 3 µm ivermectin (Iver). Three minutes later, the cells were stimulated with 10 µm ATP. Results are the means + SEM of n experiments.
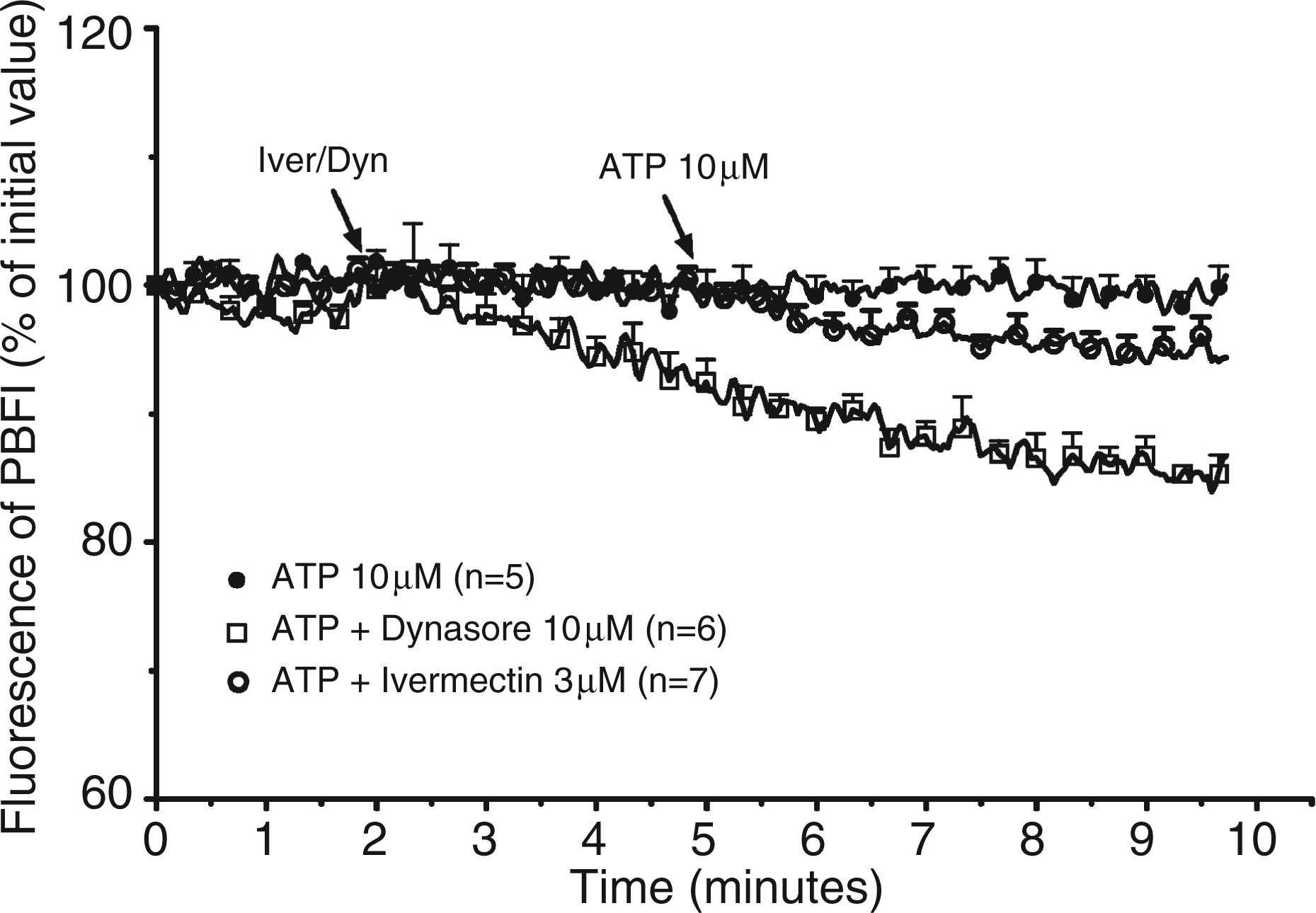
The variation of the [K+]i in response to ATP was evaluated next. As shown in Figure 4, 10 µm ATP or 3 µm ivermectin alone had no effect on the fluorescence of PBFI, a fluorescent dye sensitive to the concentration of potassium. The addition of ATP to ivermectin-treated cells provoked a slight but reproducible decrease of the fluorescence (94.3 ± 1.7 % after 5 min, n = 7). Dynasore (10 µm) provoked by itself a sustained decrease of the fluorescence of the dye which dropped to 85 ± 1 % after 7 min (n = 6). The addition of ATP to the medium did not provoke a further decrease of the signal (Figure 4). These results suggested that dynasore increased the permeability of the plasma membrane to potassium, which decreased the [K+]i. The lack of effect of dynasore on ATP-mediated ion flux was in accordance with our previous observation that showed that non-toxic concentrations of dynasore did not increase P2X4 surface expression in LPS-primed peritoneal macrophages.
Effect of dynasore on the [K+]i of LPS-primed murine peritoneal macrophages. LPS-primed macrophages from P2X7-knockout (KO) mice were loaded with PBFI for 90 min at 37°C. After washing the cells were incubated at 37°C in the cuvette of a fluorimeter under constant stirring. Two min after the start of the measurement, the cells were either maintained in control conditions or exposed to 10 µm dynasore (Dyn) or to 3 µm ivermectin (Iver). Three min later, the cells were stimulated with 10 µm ATP. Results are expressed as percent of the initial fluorescence and are the means + SEM of n experiments.
Effect of dynasore on the secretion of IL-1β
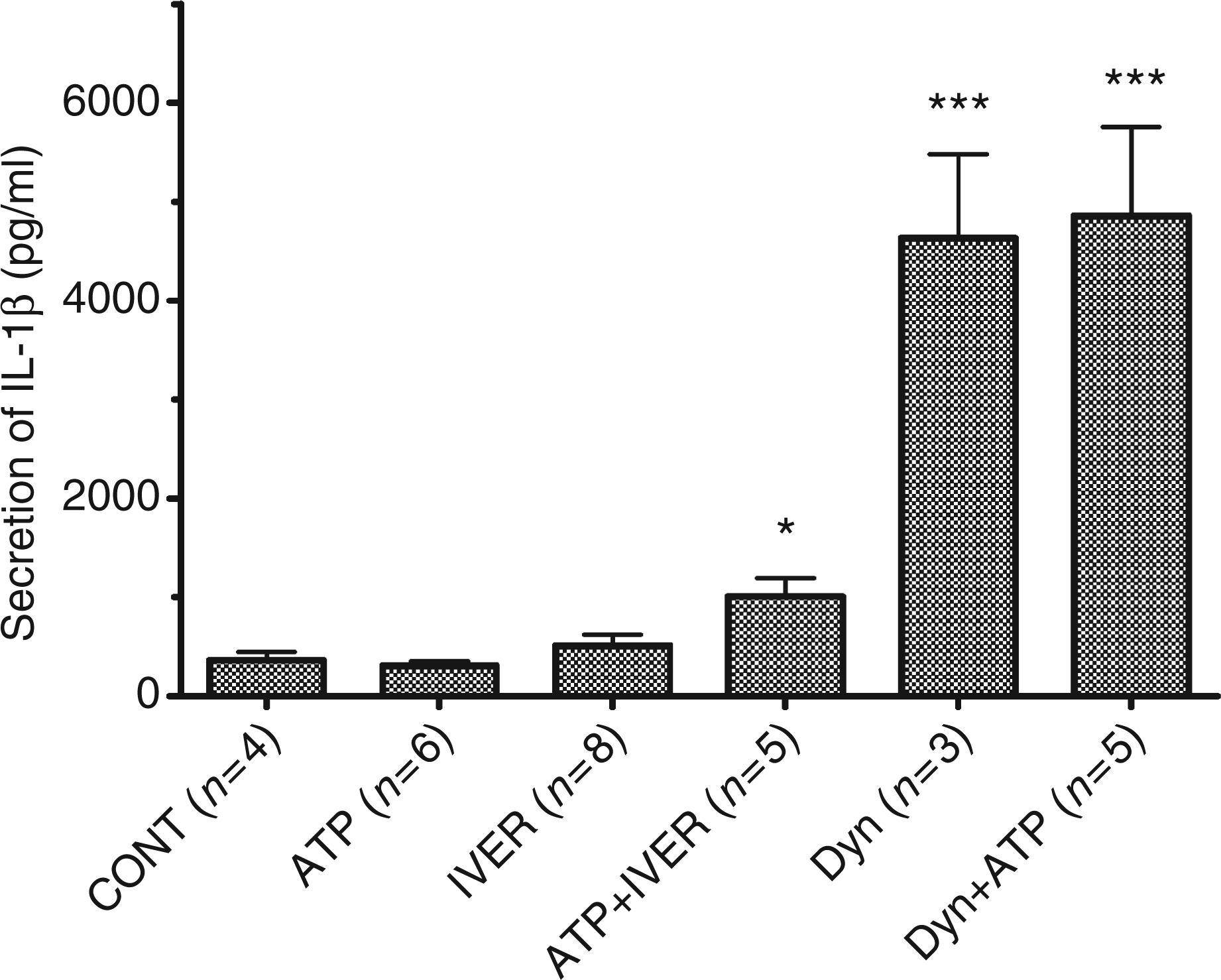
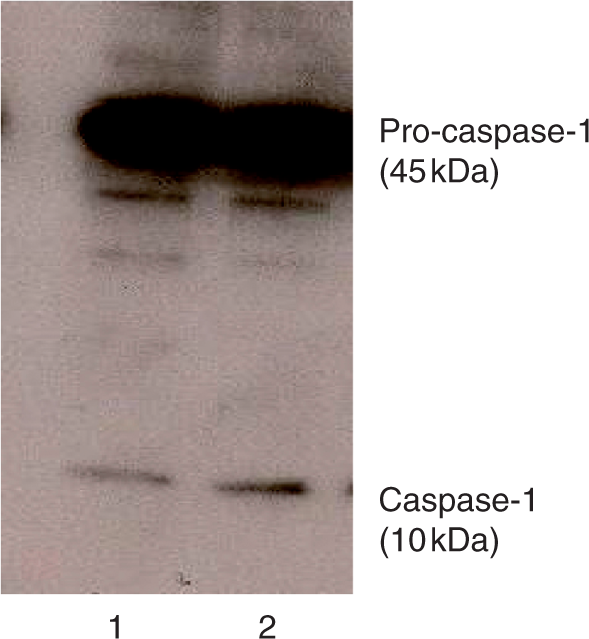
It is now clearly established that the drop in the [K+]i is essential to the formation of the NALP3 inflammasome, the activation of procaspase-1 and the secretion of IL-1β.25 The secretion of IL-1β in response to 10 µm ATP and/or ivermectin or dynasore was thus measured. As shown in Figure 5, the presence of ATP or ivermectin in the medium did not affect the basal release of immunoreactive IL-1β (from 365 ± 80 pg/ml to 312 ± 42 pg/ml and 513 ± 107 pg/ml respectively, P > 0.1) but the combination of the two drugs provoked the release of the immunoreactive cytokine (from 312 ± 42 pg/ml to 1005 ± 192 pg/ml, n = 5, P = 0.043). In the presence of dynasore, the secretion of IL-1β increased to 4634 ± 837 pg/ml (n = 3). The presence of ATP did not affect the response to dynasore (4855 ± 898 pg/ml, n = 5). By comparison, nigericin, a K + /H+ exchanger which decreases the [K+]i stimulated the release of IL-1β to 8972 ± 1608 pg/ml (n = 9, data not shown). These results suggested that dynasore stimulated the NALP3 inflammasome. In the following experiment, the effect of dynasore on the activation of caspase-1 was examined. We performed an SDS-PAGE to detect both the inactive (p45) and the active (p10) forms of caspase-1 in whole cell homogenates. The migration of immunoreactive proteins obtained from the intracellular content of treated cells was tested by immunoblotting. As shown in Figure 6, a strong band of 45 kDa was present in control and dynasore-treated samples. Caspase-1 antibody enabled us to detect another band of 10 kDa. This low-molecular mass form of caspase-1 most probably resulted from pro-caspase-1 cleavage upon NALP3 inflammasome formation.25 The p10 band was more pronounced in homogenate from cells treated with dynasore suggesting that the drug promoted the cleavage of pro-caspase-1.
Secretion of IL-1β by LPS-primed murine peritoneal macrophages. LPS-primed macrophages from P2X7-knockout (KO) mice were incubated at 37°C for 15 min in control conditions (CONT) or in the presence of 10 µm ATP or 10 µm dynasore (Dyn) or 3 µm ivermectin (IVER) or a combination of ATP + ivermectin/dynasore. The concentration of IL-1β was assayed in the supernatant. Results are the means + SEM of n experiments.
Immunoblot assay of intracellular caspase-1 in LPS-primed murine peritoneal macrophages. LPS-primed macrophages from P2X7-knockout (KO) mice were incubated at 37°C for 15 min in control conditions (1) or in the presence of 10 µm dynasore (2). Immunoblot of caspase-1 present in the lysate was performed as described in the Materials and Methods section.
Discussion
In this work we showed that a low concentration of ATP transiently increased the [Ca2+]i in macrophages from P2X7-KO mice. The addition of ivermectin to the incubation buffer strongly potentiated this increase of the [Ca2+]i. In the presence of this drug, ATP also decreased the [K+]i and promoted the release of mature IL-1β. These responses were mediated by a purinergic receptor with properties (responses of short duration; rather high sensitivity to the agonist; permeability to calcium and potassium; sensitivity to ivermectin) which characterize P2X4 receptors.26 These receptors are, indeed, expressed by murine peritoneal macrophages,27,28 as confirmed in this work. Treatment of the cells with a non-permeant biotinylating reagent showed that only a small population of these receptors was accessible to the cross-linking reagent. The major intracellular location of the receptor probably accounts for the small responses elicited by the interaction between ATP and the receptor. The intracellular localization of P2X4 receptors has been recently confirmed.29 Stokes and Surprenant showed that the P2X4 receptors were located on the lysosomal membrane and that they rapidly trafficked to the plasma membrane after induction of endolysosomal secretion.29 Ivermectin potentiated the responses coupled to this receptor without modifying its cellular distribution. This is at variance with the results of Toulmé et al. who reported that ivermectin increased the location in the plasma membrane of P2X4 receptors expressed in Xenopus oocytes.30 Using the same expression system, Asatryan et al. recently reported that 3 µm ivermectin did not affect the surface expression of P2X4 receptors.31 These results, and ours, are consistent with the current view that the drug interacts with an allosteric site on the receptor rather than on its cellular internalization.24,32 The stimulation of P2X4 receptors by the combination of ATP plus ivermectin not only potentiated the increase of the [Ca2+]i but also promoted the drop of the [K+]i and the release of IL-1β. These results confirmed the contribution of P2X4 receptors to the secretion of pro-inflammatory cytokines by macrophages.28
Dynasore proved to be toxic to murine peritoneal macrophages at concentrations above 10 µm. It was recently reported that the combination of LPS with 80 µm dynasore was toxic for bone marrow derived macrophages (BMDM).17 At a low, non-toxic concentration, dynasore did not potentiate the increase of the [Ca2+]i in response to a low concentration of ATP suggesting that it did not potentiate the response coupled to P2X4 receptors or the surface expression of the receptor. Higher concentrations of dynasore (50 and 80 µm) increased the surface expression of the receptor (data not shown), but considering the permeabilizing effect of these concentrations of dynasore, these experiments are difficult to interpret. The apparent increased fraction of the P2X4 receptors in the plasma membrane may, indeed, be secondary to a better access of the biotinylating agent to the intracellular pool of receptors. Boumechache et al. reported that in microglial cells, dynasore increased the surface expression of P2X4 receptors but that the drug had no effect on the surface biotinylation of P2X4 receptors of BMDM.17 These results suggest heterogeneity among tissues with respect to trafficking of P2X4 receptors. Marina-García et al. recently showed that a high concentration of dynasore (80 µm) inhibited the activation of murine BMDM by ATP.33 These BMDMs were obtained from wild-type mice and the response to ATP was probably mediated by P2X7 receptors, not by P2X4 receptors. At this high (80 µm) concentration, dynasore has also been reported to inhibit the expression of pro-IL-1β by macrophages stimulated with oxidized low-density lipoprotein (LDL)34 or the activation of caspase-1 by muramyl-dipeptide and ATP.33 Our results show that at a low concentration, dynasore did not inhibit, but rather stimulated, the secretion of IL-1β from macrophages primed with LPS. This secretion was probably secondary to the decrease of the [K+]i and ensuing activation of pro-caspase-1 as suggested by the increased intensity of the p10 subunit of caspase-1 in cells treated with dynasore. Macrophages express various potassium channels, some of which are internalized with a clathrin-mediated mechanism.35 Dynasore, by blocking the intracellular transfer of these proteins might facilitate the efflux of potassium. The stimulatory effect of dynasore on IL-1β secretion is reminiscent of the results reported by Chung et al.36 According to these authors, 10 µm dynasore stimulated the expression of plasminogen activator inhibitor-1 (PAI-1) by human pleural mesothelial cells. This observation led them to conclude that dynasore might be a pleurodesing agent in vivo.
Considering the stimulatory effect of dynasore on the maturation of caspase-1 and on the secretion of the pro-inflammatory cytokine IL-1β by macrophages after activation of potassium efflux, our results suggest that, at this concentration, this drug may also provoke an inflammatory syndrome independently of any purinergic receptors activation.
Footnotes
Abbreviations
Acknowledgements
This work was supported by grant n° 3.4.528.07.F from the Fonds National de la Recherche Scientifique to J.P. Dehaye. M. El Ouaaliti is supported by the Belgian Fund for ‘Formation à la Recherche dans l’Industrie et l’Agriculture’. The authors thank S. Dulanto, R. Antoine, V. Delpoux and S. Patel for their help.
References
1.
SurprenantANorthRA. Signaling at purinergic P2X receptors. Annu Rev Physiol2009; 71: 333–359.
2.
RobertsJAVialCDigbyHR. Molecular properties of P2X receptors. Pflugers Arch2006; 452: 486–500.
3.
KimMJiangLHWilsonHLNorthRASurprenantA. Proteomic and functional evidence for a P2X7 receptor signalling complex. EMBO J2001; 20: 6347–6358.
4.
Donnelly-RobertsDLNamovicMTHanPJarvisMF. Mammalian P2X7 receptor pharmacology: comparison of recombinant mouse, rat and human P2X7 receptors. Br J Pharmacol2009; 157: 1203–1214.
5.
NovakIJansIMWohlfahrtL. Effect of P2X7 receptor knockout on exocrine secretion of pancreas, salivary glands and lacrimal glands. J Physiol2010; 588: 3615–3627.
6.
NorthRA. Molecular physiology of P2X receptors. Physiol Rev2002; 82: 1013–1067.
7.
DubyakGR. Go it alone no more–P2X7 joins the society of heteromeric ATP-gated receptor channels. Mol Pharmacol2007; 72: 1402–1405.
8.
QureshiOSParamasivamAYuJCMurrell-LagnadoRD. Regulation of P2X4 receptors by lysosomal targeting, glycan protection and exocytosis. J Cell Sci2007; 120: 3838–3849.
9.
RoyleSJBobanovićLKMurrell-LagnadoRD. Identification of a non-canonical tyrosine-based endocytic motif in an ionotropic receptor. J Biol Chem2002; 277: 35378–35385.
10.
TraubLM. Tickets to ride: selecting cargo for clathrin-regulated internalization. Nat Rev Mol Cell Biol2009; 10: 583–596.
11.
MettlenMPucadyilTRamachandranRSchmidSL. Dissecting dynamin's role in clathrin-mediated endocytosis. Biochem Soc Trans2009; 37: 1022–1026.
12.
BobanovicLKRoyleSJMurrell-LagnadoRD. P2X receptor trafficking in neurons is subunit specific. J Neurosci2002; 22: 4814–4824.
13.
BrownDAYuleDI. Protein kinase A regulation of P2X4 receptors: requirement for a specific motif in the C-terminus. Biochim Biophys Acta2010; 1803: 275–287.
MaciaEEhrlichMMassolRBoucrotEBrunnerCKirchhausenT. Dynasore, a cell permeable inhibitor of dynamin. Dev Cell2006; 10: 839–850.
16.
NankoeSRSeverS. Dynasore puts a new spin on dynamin: a surprising dual role during vesicle formation. Trends Cell Biol2006; 16: 607–609.
17.
BoumechacheMMasinMEdwardsonJMGóreckiDCMurrell-LagnadoR. Analysis of assembly and trafficking of native P2X4 and P2X7 receptor complexes in rodent immune cells. J Biol Chem2009; 284: 13446–13454.
18.
SolleMLabasiJPerregauxG. Altered cytokine production in mice lacking P2X7 receptors. J Biol Chem2001; 276: 125–132.
19.
SeilMKabréENagantC. Regulation by CRAMP of the responses of murine peritoneal macrophages to extracellular ATP. Biochim Biophys Acta2010; 1798: 569–578.
20.
PochetSGarcia-MarcosMSeilMOttoAMarinoADehayeJP. Contribution of two ionotropic purinergic receptors to ATP responses in submandibular gland ductal cells. Cell Signal2007; 19: 2155–2164.
21.
McCarronRMGoroffDKLuhrJEMurphyMAHerscowitzHB. Methods for the collection of peritoneal and alveolar macrophages. Methods Enzymol1984; 108: 274–284.
22.
GrynkiewiczGPoenieMTsienRY. A new generation of calcium indicators with greatly improved fluorescence properties. J Biol Chem1985; 260: 3440–3450.
23.
DenizotFLangR. Rapid colorimetric assay for cell growth and survival. Modifications to the tetrazolium dye procedure giving improved sensitivity and reliability. J Immunol Methods1986; 89: 271–277.
24.
KhakhBSProctorWRDunwiddieTVLabarcaCLesterHA. Allosteric control of gating and kinetics at P2X4 receptor channels. J Neurosci1999; 19: 7289–7299.
25.
ArlehamnCSPétrilliVGrossOTschoppJEvansTJ. The role of potassium in inflammasome activation by bacteria. J Biol Chem2010; 285: 10508–10518.
26.
KawateTMichelJCBirdsongWTGouauxE. Crystal structure of the ATP-gated P2X4 ion channel in the closed state. Nature2009; 460: 592–598.
27.
UlmannLHirbecHRassendrenF. P2X4 receptors mediate PGE2 release by tissue-resident macrophages and initiate inflammatory pain. EMBO J2010; 29: 2290–2300.
28.
SeilMEl OuaalitiMFontanilsUGorronzo ExtebarriaIPochetSDal MoroGMarinoADehayeJ-P. Ivermectin-dependent release of IL-1beta in response to ATP by peritoneal macrophages from P2X7-KO mice. Purinergic Signal2011; 6: 405–416.
29.
StokesLSurprenantA. Dynamic regulation of the P2X4 receptor in alveolar macrophages by phagocytosis and classical activation. Eur J Immunol2009; 39: 986–995.
30.
ToulméESotoFGarretMBoué-GrabotE. Functional properties of internalization-deficient P2X4 receptors reveal a novel mechanism of ligand-gated channel facilitation by ivermectin. Mol Pharmacol2006; 69: 576–587.
SilberbergSDSwartzKJ. Ivermectin interaction with transmembrane helices reveals widespread rearrangements during opening of P2X receptor channels. Neuron2007; 54: 263–274.
33.
Marina-GarcíaNFranchiLKimYG. Clathrin- and dynamin-dependent endocytic pathway regulates muramyl dipeptide internalization and NOD2 activation. J Immunol2009; 182: 4321–4327.
34.
StewartCRStuartLMWilkinsonK. CD36 ligands promote sterile inflammation through assembly of a Toll-like receptor 4 and 6 heterodimer. Nat Immunol2010; 11: 155–161.
35.
HibinoHInanobeAFurutaniKMurakamiSFindlayIKurachiY. Inwardly rectifying potassium channels: their structure, function, and physiological roles. Physiol Rev2010; 90: 291–366.
36.
ChungCLSheuJRLiuHE. Dynasore, a dynamin inhibitor, induces PAI-1 expression in MeT-5A human pleural mesothelial cells. Am J Respir Cell Mol Biol2009; 40: 692–700.