Low susceptibility of NC/Nga mice to the lipopolysaccharide-mediated lethality with d -galactosamine sensitization and the involvement of fewer natural killer T cells
Free accessResearch articleFirst published online February, 2012
Low susceptibility of NC/Nga mice to the lipopolysaccharide-mediated lethality with d -galactosamine sensitization and the involvement of fewer natural killer T cells
The LPS-mediated lethality of NC/Nga mice, having fewer NKT cells, was examined by using d-galactosamine (d-GalN)-sensitization. The NC/Nga mice were not killed by a simultaneous administration of d-GalN and LPS whereas all C57BL/6 (B6) control mice were killed. The injection of d-GalN and LPS failed to elevate the levels of serum alanine aminotransferase and caspase 3 in the liver tissues of NC/Nga mice. Further, the nitric oxide (NO) level of the d-GalN- and LPS-injected NC/Nga mice was much lower than those of the B6 mice. The expression of an inducible NO synthase (iNOS) was significantly reduced in the livers of NC/Nga mice. However, there was no significant difference in LPS-induced TNF-α production between B6 mice and NC/Nga mice. The NC/Nga mice had an impaired expression of IFN-γ protein and mRNA in response to d-GalN and LPS. The pretreatment with α-galactosylceramide (α-GalCer), which activates Vα14+ NKT cells and induces the production of IFN-γ, rendered NC/Nga mice more susceptible to the LPS-mediated lethality. The livers of NC/Nga mice had fewer NKT cells compared to B6 mice. Taken together, it is suggested that the resistance of NC/Nga mice to the LPS-mediated lethality with d-GalN sensitization depended on the impaired IFN-γ production caused by fewer NKT cells and reduced NO production that followed.
NC/Nga mice have been used for animal models for human allergic diseases.1,2 They spontaneously develop dermatitis associated with excessive IgE production under conventional conditions.3–6 The dermatitis is caused by a loss of the Th1/Th2 balance and the overproduction of Th2-type cytokines and chemokines.7–10 Moreover, the level of IFN-γ, the representative Th1 cytokine, in response to staphylococcal enterotoxin B or LPS is much lower than that of normal mice because of the lack of Vβ8+ T-cells and Vβ8+ NKT-cells.11 Thus, a systemic deficiency in the Th1 response to microbial stimulation may lead to a Th2-dominant immune response and have an impaired crosstalk in the cytokine network. This causes abnormal immune conditions in NC/Nga mice.
Recently, we reported that NC/Nga mice have a resistance to TNF-α-mediated lethality with d-galactosamine (d-GalN) sensitization.12 The resistance of NC/Nga mice to the TNF-α-mediated lethality is considered to be mediated by the sustained NF-κB activation. The sustained NF-κB activation in NC/Nga mice might be dependent on the insufficient level of IFN-γ, at least partially, caused by the lack of Vβ8+ NKT-cells. The LPS-mediated lethality with d-GalN-sensitization was mainly due to an excessive production of TNF-α and nitric oxide (NO).13–16 The LPS-mediated lethality is strongly regulated by IFN-γ, which augments the production of TNF-α and NO.17–19 In the present study, we examined the resistance of d-GalN-sensitized NC/Nga mice to LPS-mediated lethality. Here, we report that the reduced NO production caused by the impaired IFN-γ production may be responsible for the low susceptibility of NC/Nga mice to LPS-mediated lethality.
Materials and methods
Mice
Female C57BL/6 (B6) mice and NC/Nga mice were purchased from SLC (Hamamatsu, Japan) and used at 6–7 wk of age. Each experimental group contained 8–10 mice. The study protocol was approved by the Animal Care Committee and performed according to the Guide for the Care and Use of Laboratory Animals of Aichi Medical University.
Materials
Lipopolysaccharide from Escherichia coli O55:B5 was obtained from Sigma Chemical (St Louis, MO, USA) and d-GalN was from Wako Pure Chemical (Tokyo, Japan). Recombinant IFN-γ was obtained from Peprotech (Rock Hill, NJ, USA). Rabbit polyclonal antibody to an inducible type of NO synthase (iNOS) was purchased from ABR (Golden, CO, USA), and anti-nitrotyrosine antibody was purchased from Upstate (Lake Placid, NY, USA). A goat polyclonal antibody to actin was obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA). α-Galactosyceramide (α-GalCer) was obtained from Funakoshi (Tokyo, Japan).
Isolation of peritoneal cells
Mice were injected intraperitoneally with 1 ml of 10% thioglycollate (Difco Laboratories, Detroit, MI, USA). Three days after the injection, peritoneal exudate cells were collected by washing the peritoneum with 5 ml of ice-cold RPMI 1640 medium. The peritoneal cells were incubated in a 96-well culture dish (50,000 cells/well) with RPMI 1640 medium supplemented with 10% FCS at 37°C for 1 h, and non-adherent cells were removed by washing. The peritoneal adherent cells were used as macrophages.
Determination of alanine aminotransferase (ALT) level
Blood samples from individual mice were collected from the retro-orbital venous plexus, and the plasma ALT levels were determined with a Hitachi 7700 autoanalyzer.20
Determination of caspase 3 activity
The caspase 3 activity was determined by the amount of fluorochrome-labeled substrate cleaved by the enzyme with a commercially available kit (MBL, Nagoya, Japan). Briefly, livers were removed, rinsed in cold PBS at pH 7.2, homogenized, and lysed in a hypotonic buffer. An aliquot (200 µl) of the lysate was mixed with the substrate in wells of a black 96-well plate (Sumitomo Bakelite, Tokyo, Japan) for 90 min at 37°C, and the optical density was measured with the Wallace 1420 ARVO multilabel counter (Pharmacia Biotech, Buckinghamshire, UK).12
Determination of IFN-γ and TNF-α levels
The concentrations of circulating IFN-γ and TNF-α in the sera were determined by the corresponding enzyme-linked immunoabsorbent assay (ELISA) kits (R&D Systems, Minneapolis, MN, USA).20
Determination of NO
The nitrite level in supernatants was measured with a microplate reader through the Griess method as described elsewhere.21 Potassium nitrite diluted in PBS was used as the standard.22
Immunohistochemical staining
Livers were fixed in formalin, and liver sections were stained immunohistochemically as described previously.12 Briefly, the endogenous peroxidase activity in deparaffinized sections was blocked by methanol containing 0.3% hydrogen peroxide. The sections were washed in PBS containing 10% normal horse serum and incubated overnight at 4°C with anti-iNOS antibody (Spring Bioscience, Fremont, CA, USA). Horseradish peroxidase-conjugated goat anti-rabbit immunoglobulin (Ig) antibody was used at the 1:200 dilution level. Immune complexes were detected with a solution of 3,3-diaminobenzidine (0.2 mg/ml) and hydrogen peroxide in 0.05 m Tris-HCl buffer. The sections were counterstained with methyl green.
In situ specific labeling of fragmented DNA
The tissues were fixed with formalin and cut serially into 4–6 µm paraffin sections. The serial sections were deparaffinized for the in situ specific labeling of fragmented DNA. The details of the in situ specific labeling are described in our previous report.15
Immunoblotting
The immunoblotting procedures were conducted as described previously.12 The lysates of livers were diluted with an equal volume of 2× sample buffer and boiled for 5 min. Proteins were electrophoretically separated in an 8% polyacrylamide gel under reducing conditions and then transferred to a membrane by electroblotting. The membranes were blocked with 5% skimmed milk and treated with the antibody diluted to 1:1000 with PBS containing 0.05% Tween-20. After washing, the membranes were incubated with a 1:1000 dilution of peroxidase-conjugated secondary antibody. Finally, the immune complexes were detected with the supersignal west dura system (Pierce, Rockford, IL, USA) and visualized with a light-capture system (ATTO, Tokyo, Japan).
The PCR procedures for IFN-γ mRNA were conducted as described previously.23 Total RNA from liver tissues were isolated with Isogen (Nippon Gene, Tokyo, Japan) according to the manufacturer’s instructions. Reverse transcription-PCR was performed by the access quick single-tube RT-PCR system (Promega; Madison, WI, USA). The primer sequences for IFN-γ were listed in our previous report.23 The PCR products were visualized by ethidium bromide staining after separation in 2% agarose gels.
Laser flow cytometric analysis
Hepatic lymphocytes isolated from livers were stained with a 1:100 dilution of allophycocyanin (APC)-conjugated anti-CD3ε antibody and fluorescein isothiocyanate (FITC)-conjugated anti-pan-NK (DX5) antibody (eBioscience; San Diego, CA, USA) for 30 min at 4°C. To detect an intracellular cytokine, we referred to the manual of eBioscience and used their products. The isolated hepatic lymphocytes were cultured with LPS (100 ng/ml) for 6 h and incubated in the concentration of monensin (2 µm) for a further 2 h for the prevention of secretion of cytokines into the culture medium. The cells were harvested and stained with APC-conjugated anti-CD3 and FITC-conjugated anti-pan-NK antibody for 30 min. The cells were washed with PBS containing 10% fetal calf serum, fixed with the fixation buffer (eBioscience) and stained with phycoerythrin (PE)-conjugated anti-mouse IFN-γ antibody (eBioscience) in the permeabilization buffer (eBioscience) for 20 min at room temperature. The expression level was analyzed with a laser flow cytometer (FACS Calibur, Becton Dickinson; Palo Alto, CA, USA) by gating a lymphocyte fraction. The fluorescence intensity is expressed on a log scale.23
Statistical analysis
Experimental values are expressed as the mean ± SD of at least three independent experiments. Student’s t-test was used to identify significant differences between the experimental and control groups. A value of P < 0.05 was considered statistically significant.
Results
Low susceptibility of NC/Nga mice to LPS-mediated lethality with d-GalN sensitization
NC/Nga mice and C57BL/6 mice (B6) were intraperitoneally injected with d-GalN (8 mg) and LPS (1 µg) as described previously.24 The administration of d-GalN and LPS did not kill any NC/Nga mice within 24 h, whereas it killed all of the B6 mice within 12 h.
No development of hepatic lesions in NC/Nga mice by the administration of d-GalN and LPS
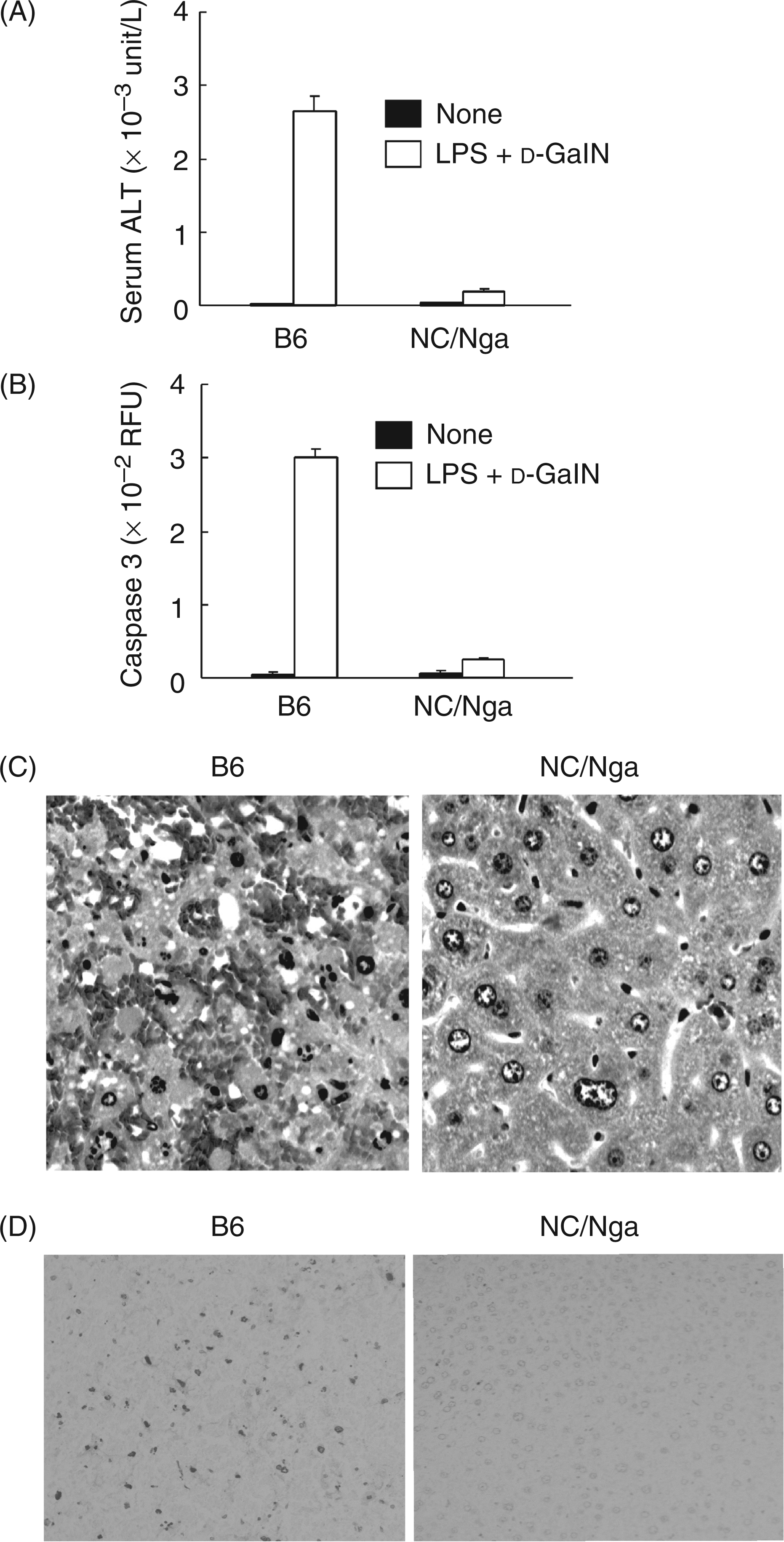
Lipopolysaccharide-mediated lethality with d-GalN sensitization is due to acute hepatic failure characterized by massive hepatocyte apoptosis.13 Therefore, we studied the histological changes in the livers of the NC/Nga mice and of the B6 mice after the d-GalN and LPS injection. To compare the NC/Nga and B6 mice in LPS-induced hepatic damages, we determined the activities of ALT and caspase 3 in sera and liver extracts, respectively. Higher levels of ALT and caspase 3 activities were observed in the B6 mice compared to the NC/Nga mice (Figure 1A,B). Histologically, no hepatic damage was found in the NC/Nga mice injected with d-GalN and LPS, while marked hepatic lesions with hemorrhage were seen in the B6 control mice (Figure 1C). The in situ specific labeling of fragmented DNA demonstrated that few apoptotic hepatocytes were found in livers from the NC/Nga mice injected with d-GalN and LPS, while a number of apoptotic cells were seen in livers from the B6 control mice (Figure 1D).
Hepatic injury in the livers of NC/Nga mice injected with D-GalN and LPS. Mice were intraperitoneally injected with D-GalN (8 mg) and LPS (1 µg). The serum ALT level (A) and caspase 3 activity in the liver extract (B) were determined 6 h after the injection. (C) Liver sections were stained with H&E. Magnification ×200. (D) Liver sections were stained with the in situ specific labeling of fragmented DNA. Magnification ×100.
Lower expression of iNOS in NC/Nga mice treated with d-GalN and LPS
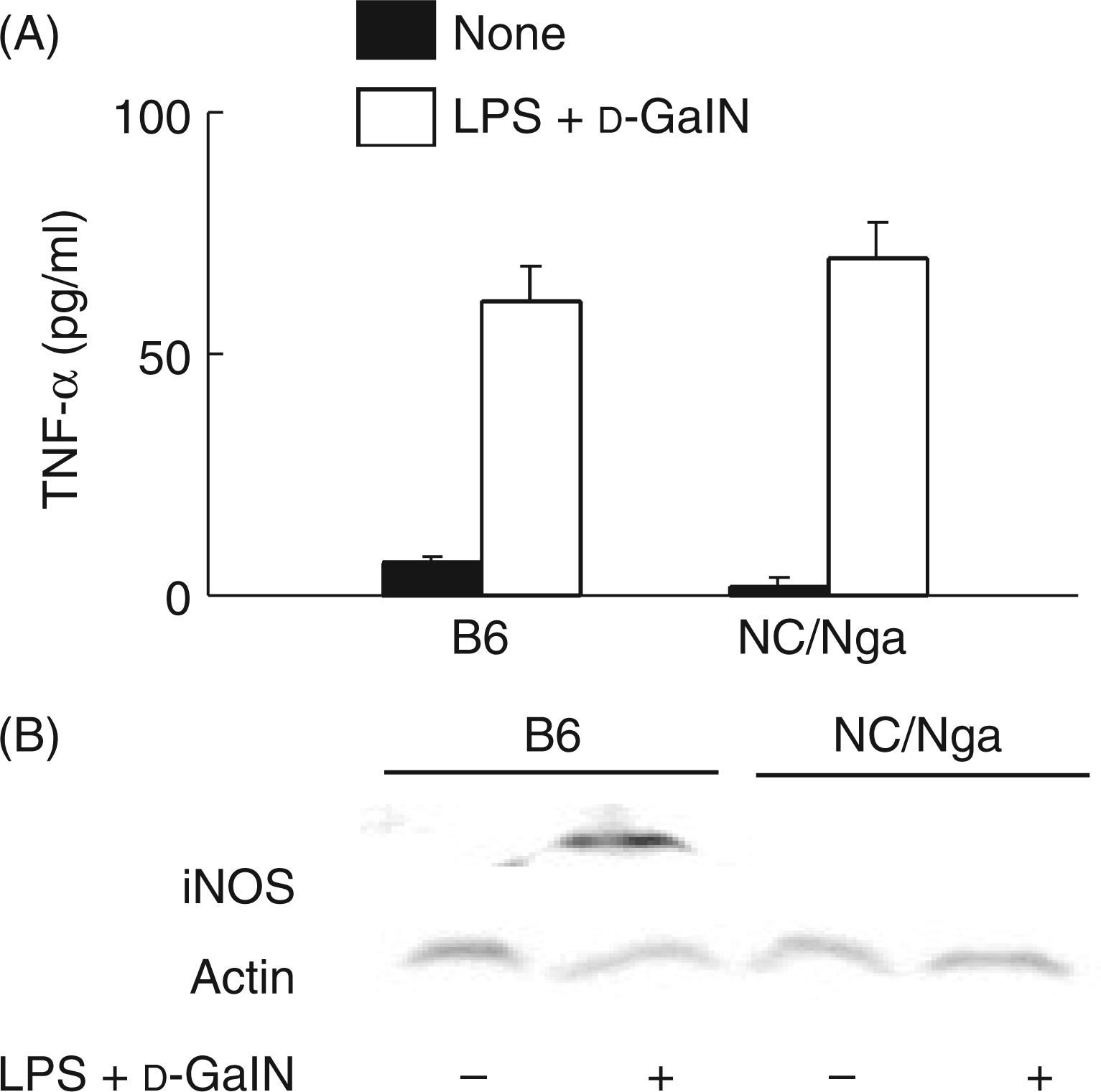
Tumor necrosis factor-α and NO are the major factors in the LPS-mediated, d-GalN sensitized lethality.13–16 First, LPS-induced TNF-α production was examined (Figure 2A). There was no significant difference in TNF-α production between the two d-GalN and LPS injected groups of the NC/Nga and B6 mice. Second, we examined the expression of iNOS in the two injected groups of NC/Nga and B6 mice LPS by immunoblotting. The immunoblot analysis showed that, 6 h after the LPS treatment, iNOS expression was significantly reduced in the NC/Nga mice, while not in the B6 mice (Figure 2B). Moreover, anti-iNOS or nitrotyrosine antibody positively stained a number of hepatocytes in the B6 mice but not in the NC/Nga mice (data not shown), suggesting the involvement of reduced NO production in the resistance of NC/Nga mice.
The expression of TNF-α and iNOS in NC/Nga mice injected with D-GalN and LPS. (A) Sera were collected 1 h after the treatment, and the level of TNF-α production determined by ELISA. (B) Liver tissues were collected 6 h after the treatment. The liver extract was analyzed by immunoblotting with anti-iNOS antibody.
No difference in LPS-induced NO production between peritoneal macrophages of NC/Nga and those of B6 mice
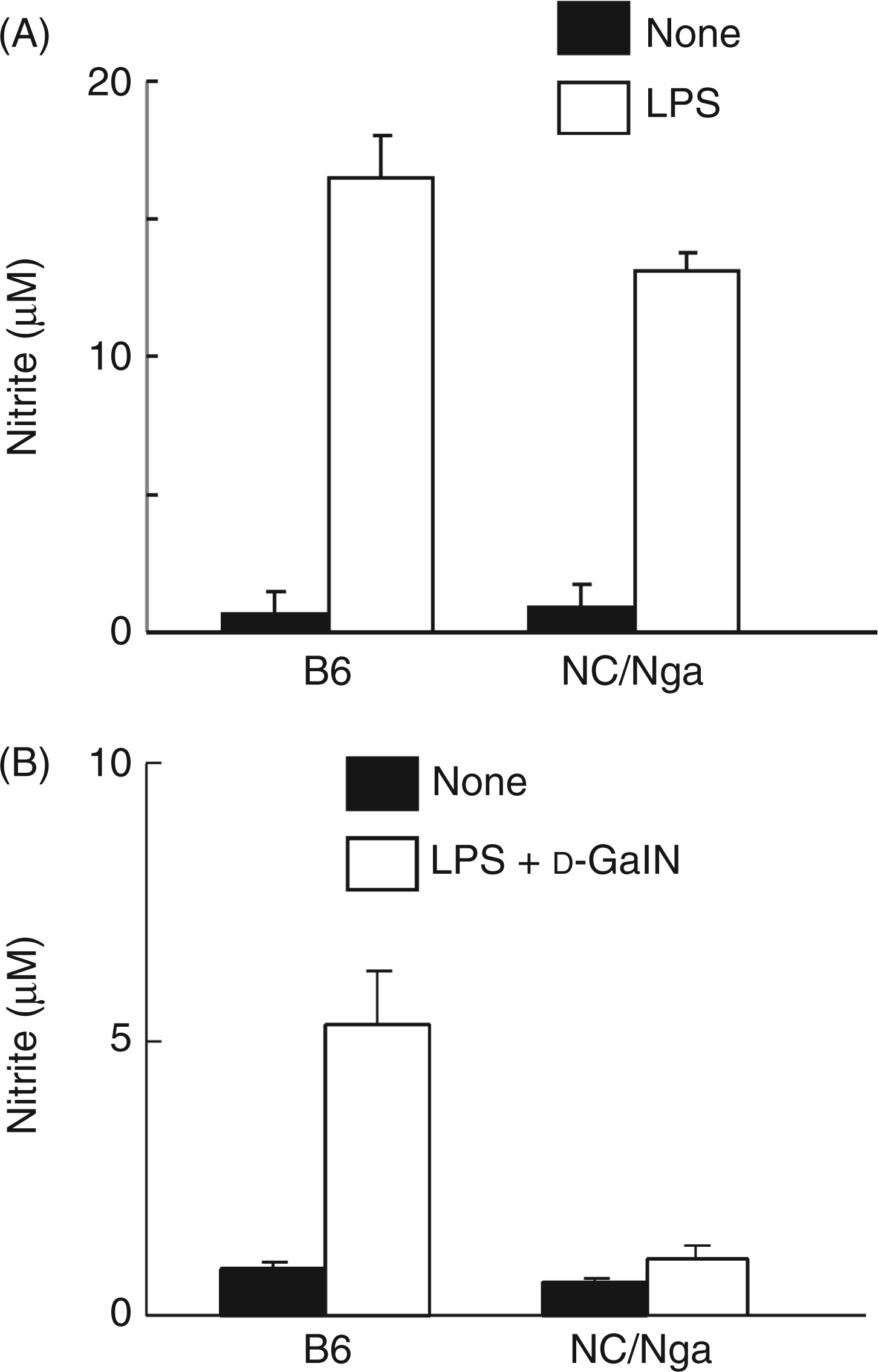
First, peritoneal macrophages of normal NC/Nga and B6 mice, which consisting of adherent cells, were stimulated by LPS (100 ng/ml) alone for 24 h. There was no significant difference in LPS-induced NO production between the peritoneal macrophages of the NC/Nga mice and those of the B6 mice (Figure 3A). In addition, there was no significant difference in LPS-induced NO production between the liver macrophages of the NC/Nga and those of the B6 mice although the NO amount was much less than that in the peritoneal macrophages (data not shown). Next, peritoneal macrophages were taken from NC/Nga and B6 mice injected with d-GalN and LPS 3 h before and cultured for the measurement of NO production (Figure 3B). The peritoneal macrophages of the injected B6 mice produced the higher levels of NO. On the other hand, the peritoneal macrophages from the injected NC/Nga mice produced only a small amount of NO. It was suggested that in vivo injection with d-GalN and LPS did not enhance NO production in the peritoneal macrophages from the NC/Nga mice, unlike the B6 mice. Thus, peritoneal macrophages from untreated NC/Nga and B6 mice, which contained few non-adherent cells, were shown to have normal NO productivity in in vitro stimulation with LPS. However, peritoneal macrophages from NC/Nga mice receiving d-GalN and LPS in vivo produced much less NO than those from B6 mice receiving them, suggesting peritoneal macrophages from NC/Nga mice, unlike B6 mice, could not become susceptible for NO production in in vivo response to d-GalN and LPS. It might be related to the fact that NC/Nga mice produce a lower level of IFN-γ because of the lack of Vβ8+ NKT cells.11
The production of NO in the peritoneal macrophages from NC/Nga mice. (A) Peritoneal macrophages from NC/Nga and B6 mice were stimulated with LPS (100 ng/ml) for 24 h. (B) Peritoneal cells were collected 3 h after the injection with D-GalN and LPS, and the peritoneal macrophages were incubated for 24 h. The nitrite level in supernatants was determined by the Griess method.
Impaired IFN-γ production in NC/Nga mice receiving an injection of d-GalN and LPS
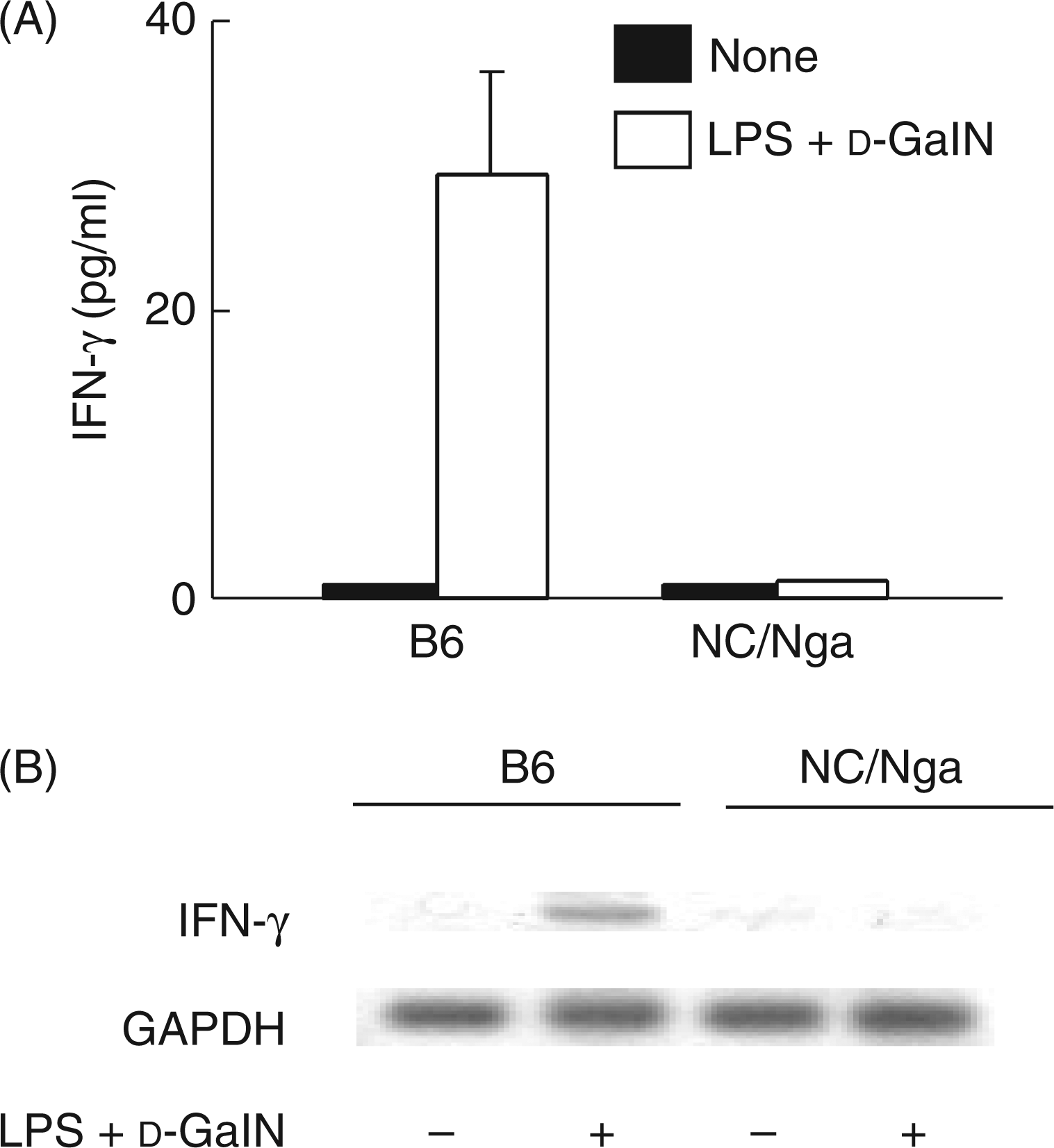
To confirm the lower level of IFN-γ in NC/Nga mice11, the levels of serum IFN-γ and IFN-γ mRNA in the liver extracts of the two d-GalN and LPS injected groups of NC/Nga and B6 mice were determined by ELISA and RT-PCR, respectively. The NC/Nga mice did not produce a significant level of IFN-γ 3 h after the d-GalN and LPS treatment (Figure 4A). Similarly, there was no IFN-γ mRNA expression in the NC/Nga mice 3 h after the d-GalN and LPS treatment (Figure 4B). However, treatment with d-GalN and LPS led to the expression of IFN-γ protein and mRNA in the B6 mice.
The expression of IFN-γ protein and mRNA in NC/Nga mice injected with d-GalN and LPS. The sera and livers were collected 3 h after the injection of d-GalN and LPS. The levels of IFN-γ protein in the sera and IFN-γ mRNA in the livers were determined by ELISA (A) or RT-PCR (B), respectively.
Presence of fewer number of NKT cells in NC/Nga mice
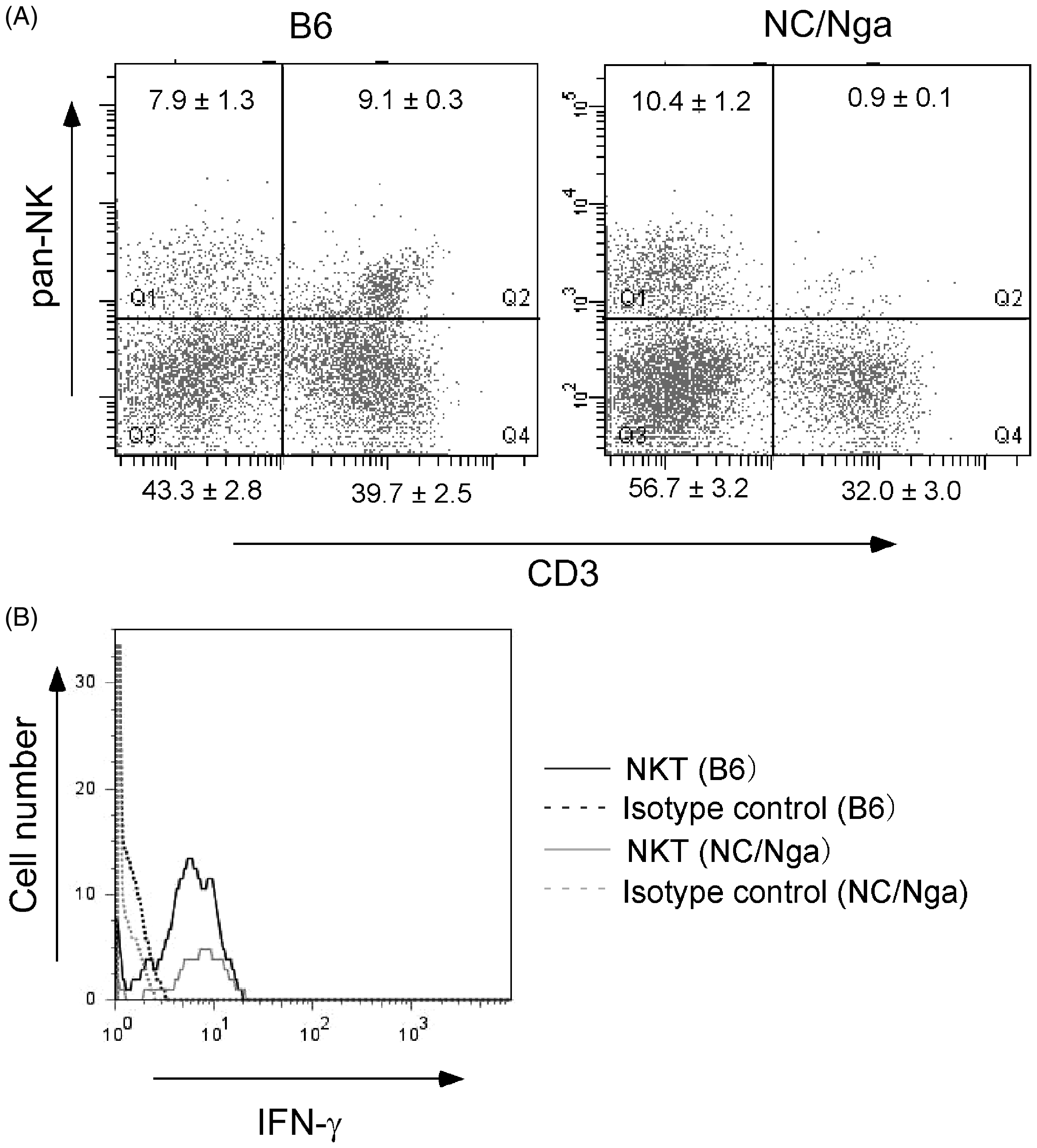
First, hepatic lymphocytes were prepared from livers of normal NC/Nga mice and B6 mice and stained with fluorescent antibodies to pan-NK and CD3ε. The frequencies of NKT cells expressing pan-NK and CD3 antigens in the livers of the NC/Nga mice and those of B6 mice were compared (Figure 5A). Laser flow cytometric analysis demonstrated the presence of fewer NKT cells (0.9%) in the livers of the NC/Nga mice. Next, we examined whether NKT cells of the NC mice can produce IFN-γ in response to LPS like NKT cells of the B6 mice. The NKT cells of the NC/Nga and B6 mice were cultured with LPS (100 ng/ml) for 8 h. Laser flow cytometric analysis demonstrated similar intracellular IFN-γ expression in both groups of the NKT cells, suggesting that the IFN-γ-producing ability of NKT cells of the NC/Nga mice was the same as that of NKT cells of the B6 mice (Figure 5B).
The frequency of NKT cells in NC/Nga mice and the IFN-γ-producing ability of NKT cells from NC/Nga mice and B6 mice. (A) Hepatic lymphocytes were isolated from NC/Nga and B6 mice and stained with APC-conjugated anti-CD3ε antibody and FITC-conjugated anti-pan-NK antibody. The value in each quadrant represents the percentage (mean ± SEM; n = 5) in the whole hepatic lymphocytes. (B) Intracellular IFN-γ was detected by PE-conjugated anti-mouse IFN-γ antibody.
Susceptibility of α-GalCer pretreated NC/Nga mice to d-GalN and LPS-mediated lethality
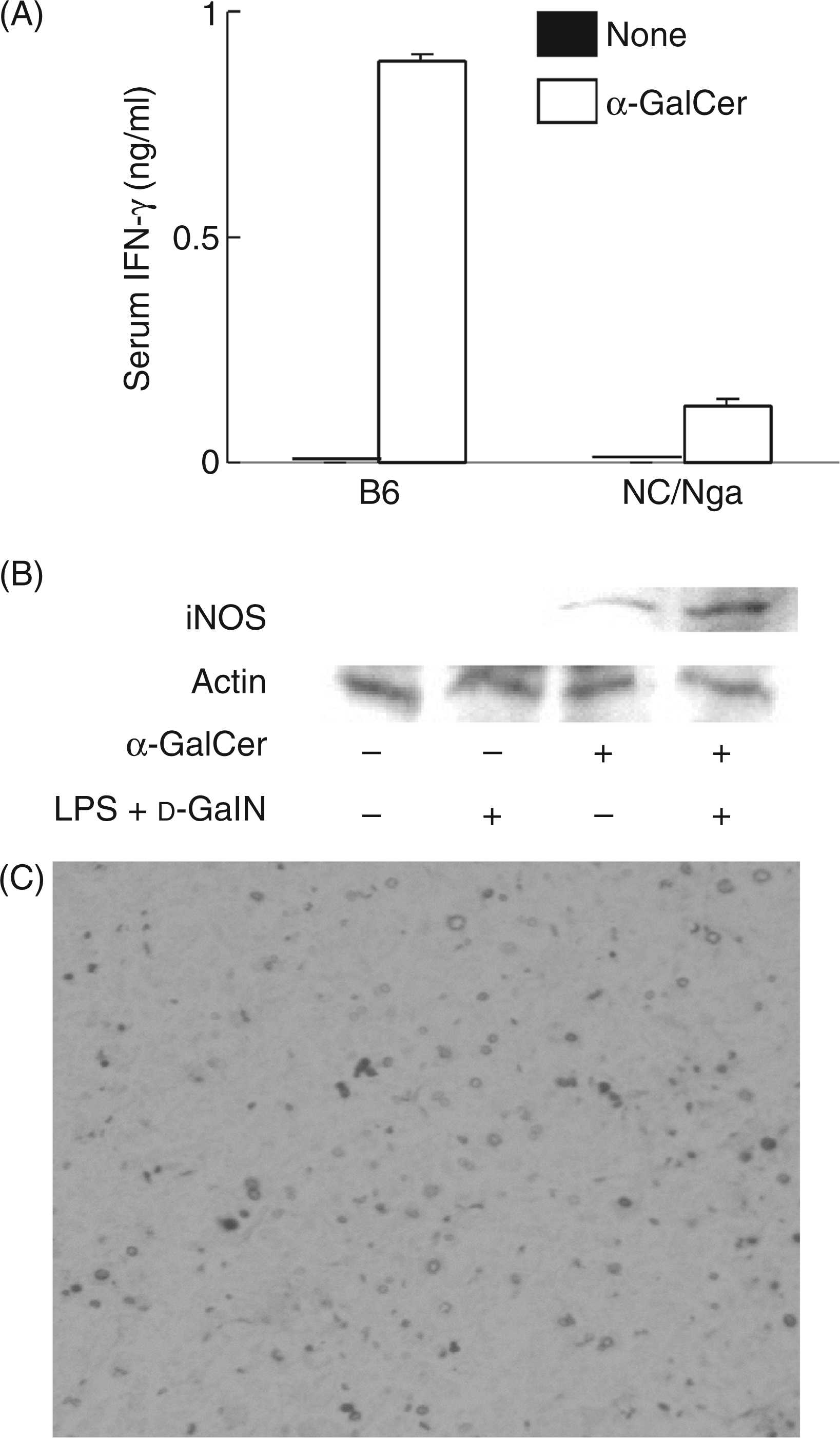
NC/Nga mice have a smaller number of Vβ8+ NKT cells and produce a lower level of IFN-γ.11 In order for endogenous IFN-γ production in NC/Nga mice to be augmented, 1 µg of α-GalCer, which induces IFN-γ production via activation of NKT cells, was intravenously administered into both groups of NC/Nga and B6 mice. Interferon-γ was produced in the NC/Nga and B6 mice 8–72 h after the α-GalCer treatment, and the peak was at 24 h. Therefore, serum IFN-γ levels of the NC/Nga and B6 mice 24 h after the treatment were compared (Figure 6A). A significant amount of IFN-γ was detected in the α-GalCer-injected NC/Nga mice, though the IFN-γ level in the B6 mice was much higher. The iNOS expression was also examined (Figure 6B). The pretreatment with α-GalCer induced definite iNOS expression in the d-GalN and LPS-treated NC/Nga mice. The in situ labeling of fragmented DNA demonstrated that the d-GalN and LPS treatment caused a number of apoptotic cells in livers from α-GalCer-pretreated NC/Nga (Figure 6C). Development of few apoptotic cells in livers of non-pretreated NC/Nga mice is shown in Figure 1D. In addition, α-GalCer did not increase the number of NKT cells in the livers (data not shown).
The production of IFN-γ and iNOS expression in NC/Nga mice injected with α-GalCer. NC/Nga and B6 mice were injected i.p. with α-GalCer (1 µg), and injected with d-GalN and LPS 18 h after α-GalCer sensitization. The livers of NC/Nga mice were collected 6 h after the injection of d-GalN and LPS. The production of IFN-γ (A) and iNOS expression (B) were analyzed by ELISA and immunoblotting, respectively. The liver sections of the α-GalCer-pretreated NC/Nga mice after the injection of d-GalN and LPS were stained with the in situ specific labeling of fragmented DNA (C). Magnification ×100.
The effect of pretreatment with α-GalCer on the LPS-mediated lethality was examined. Mice were intraperitonealy injected with α-GalCer (1 µg) 24 h before d-GalN and LPS treatment. The α-GalCer pretreated NC/Nga mice were all killed after the d-GalN and LPS treatment, whereas NC/Nga mice without the pretreatment were not killed. When α-GalCer was administered to the NC/Nga mice 72 h before d-GalN and LPS treatment, 6 of the 8 mice were killed. Next, we administered recombinant IFN-γ to NC/Nga mice 1 h prior to d-GalN and LPS treatment. The pretreatment with recombinant mouse IFN-γ (1 µg per mouse) sensitized the NC/Nga mice to d-GalN and LPS, and 8 of the 10 mice were killed.
Discussion
In the present study, we demonstrate that NC/Nga mice are resistant to the LPS-mediated lethality with d-GalN sensitization via the reduced NO production, and the reduction is caused by impaired IFN-γ production, which is, in turn, caused by fewer Vβ8+ NKT cells. The LPS-mediated lethality with d-GalN sensitization is induced by a number of pro-inflammatory mediators.13–16 Especially, TNF-α and NO are the main effector molecules in the lethality. The finding that there was no significant difference in TNF-α production between NC/Nga mice and B6 control mice suggests that the LPS resistance of NC/Nga mice is due to their impaired NO production. However, we have recently reported that NC/Nga mice are less sensitive to d-GalN and TNF-α-mediated lethality.12 Therefore, though there is no significant difference in serum TNF-α levels between NC/Nga and B6 mice, NC/Nga mice must be resistant to a toxic action of TNF-α. The LPS resistance of d-GalN-sensitized NC/Nga mice might be accounted for by the combination of reduced NO production and low TNF-α sensitivity.
Interferon-γ exhibits a priming effect on the LPS-mediated lethality and markedly augments the LPS sensitivity.25–27 Both IFN receptor-knockout mice and STAT1-knockout mice are resistant to d-GalN and LPS-mediated lethality.28,29 Therefore, it is suggested that IFN-γ plays a crucial role in the sensitization to the lethal reaction. Interestingly, NC/Nga mice have a lower level of IFN-γ production in response to d-GalN and LPS. The treatment with exogenous recombinant IFN-γ as well as the pretreatment with α-GalCer to enhance endogenous IFN-γ production renders NC/Nga mice more susceptible to the d-GalN and LPS-mediated lethality, suggesting a close relationship between the impaired IFN-γ production and the LPS resistance. Therefore, the impaired IFN-γ production in NC/Nga mice might correlate with reduced NO production.
There are several reports on the impaired IFN-γ production in NC/Nga mice. Habu et al.11 reported that IFN-γ levels in the sera of NC/Nga mice injected with LPS was several fold lower than those of normal mice, and that the low IFN-γ response to LPS correlated with an impaired IL-18 production of macrophages. Further, they reported that NC/Nga mice have partial resistance to the generalized Shwartzman model induced by IL-12 and LPS.30 The studies are consistent with our finding that NC/Nga mice have a low susceptibility against d-GalN/LPS-mediated lethality via the impaired IFN-γ production. Fujii et al.31 reported a new model of NKT cell-mediated hepatitis using α-GalCer and D-GalN. The hepatitis model seems to be caused by the hepatic toxicity of d-GalN to IFN-γ-sensitized mice. However, no hepatitis was induced in NC/Nga mice by using α-GalCer and d-GalN. This suggests that treatment with α-GalCer and D-GalN may not induce a sufficient amount of IFN-γ in NC/Nga mice for the development of the hepatitis. In addition, Matsumoto et al.4 reported that the addition of exogenous IFN-γ to IL-4 and LPS completely abrogated IgE production by B cells of BALB/c mice, but was insufficient to suppress it by B cells of NC/Nga mice. These findings suggest that NC/Nga mice may have a lower response to IFN-γ than normal mice. However, the present study demonstrates that IFN-γ pretreatment renders NC/Nga mice more susceptible to LPS-mediated lethality, despite the low response of NC/Nga mice to IFN-γ.
How are NKT cells involved in the d-GalN and LPS-mediated lethality, known as an endotoxin shock model? Previous studies11,30 reported the presence of fewer NKT cells in the peritoneal cavities and livers of NC/Nga mice as well as the present study. Interestingly, NOD mice, known as a non-obesity diabetes model, are also resistant to d-GalN/LPS-mediated lethality24 and have functional defects in NKT cells.32 Therefore, NKT cells are very likely to be involved in the lethal action of LPS. Further, NKT cells might be involved in the development of sepsis and septic shock. Once again, the present study suggests that the decreased number of NKT cells is responsible for the resistance of NC/Nga mice to d-GalN and LPS-mediated lethality. In addition, there are a significant number of NK cells that can produce IFN-γ in NC/Nga mice. It is still unknown why NK cells do not produce sufficient IFN-γ for the induction of the d-GalN and LPS-mediated lethal reaction in NC/Nga mice.
Conclusions
NC/Nga mice have fewer Vβ8+ NKT cells and produce a lower level of IFN-γ production in response to LPS. The impaired IFN-γ production fails to augment LPS-induced NO production. It might be of interest to study the importance of IFN-γ in clinical septic shock as well as experimental endotoxic shock.
Footnotes
Acknowledgements
This work was supported, in part, by a Grant-in-Aid for Scientific Research (Kakenhi: 21590499) from the Ministry of Education, Science, Sports and Culture of Japan. The authors are grateful to K. Takahashi for excellent technical assistance.
References
1.
GaoXKNakamuraNFusedaK, et al.. Establishment of allergic dermatitis in NC/Nga mice as a model for severe atopic dermatitis. Biol Pharm Bull2004; 27: 1376–1381.
2.
ShioharaTHayakawaJMizukawaY. Animal models for atopic dermatitis: are they relevant to human disease?. J Dermatol Sci2004; 36: 1–9.
3.
TeradaMTsutsuiHImaiY, et al.. Contribution of IL-18 to atopic-dermatitis-like skin inflammation induced by Staphylococcus aureus product in mice. Proc Natl Acad Sci USA2006; 103: 8816–8821.
4.
MatsumotoMItakuraATanakaAFujisawaCMatsudaH. Inability of IL-12 to down-regulate IgE synthesis due to defective production of IFN-gamma in atopic NC/Nga mice. J Immunol2001; 167: 5955–5962.
5.
MatsumotoMRaCKawamotoK, et al.. IgE hyperproduction through enhanced tyrosine phosphorylation of Janus kinase 3 in NC/Nga mice, a model for human atopic dermatitis. J Immunol1999; 162: 1056–1063.
6.
MiharaKKurataniKMatsuiTNakamuraMYokotaK. Vital role of the itch-scratch response in development of spontaneous dermatitis in NC/Nga mice. Br J Dermatol2004; 151: 335–345.
7.
HoriuchiYBaeSKatayamaINishiokaK. Therapeutic effects of streptococcal preparation OK-432 on atopic dermatitis-like lesions in NC/Nga mice: possible shift from a Th2- to Th1-predominance. J Dermatol Sci2004; 35: 187–197.
8.
ParkEJParkKCEoH, et al.. Suppression of spontaneous dermatitis in NC/Nga murine model by PG102 isolated from Actinidia arguta. J Invest Dermatol2007; 127: 1154–1160.
9.
HattoriKNishikawaMWatcharanurakK, et al.. Sustained exogenous expression of therapeutic levels of IFN-gamma ameliorates atopic dermatitis in NC/Nga mice via Th1 polarization. J Immunol2010; 184: 2729–2735.
10.
TanabeSHochiS. Oral administration of a galactooligosaccharide preparation inhibits development of atopic dermatitis-like skin lesions in NC/Nga mice. Int J Mol Med2010; 25: 331–336.
11.
HabuYSekiSTakayamaE, et al.. The mechanism of a defective IFN-gamma response to bacterial toxins in an atopic dermatitis model, NC/Nga mice, and the therapeutic effect of IFN-gamma, IL-12, or IL-18 on dermatitis. J Immunol2001; 166: 5439–5447.
12.
KoideNMorikawaANaikiY, et al.. Low susceptibility of NC/Nga mice to tumor necrosis factor-alpha-mediated lethality and hepatocellular damage with d-galactosamine sensitization. Clin Immunol2009; 130: 225–232.
13.
LeistMGantnerFBohlingerIGermannPGTiegsGWendelA. Murine hepatocyte apoptosis induced in vitro and in vivo by TNF-alpha requires transcriptional arrest. J Immunol1994; 153: 1778–1788.
MorikawaASugiyamaTKatoY, et al.. Apoptotic cell death in the response of d-galactosamine-sensitized mice to lipopolysaccharide as an experimental endotoxic shock model. Infect Immun1996; 64: 734–738.
16.
MorikawaAKatoYSugiyamaT, et al.. Role of nitric oxide in lipopolysaccharide-induced hepatic injury in d-galactosamine-sensitized mice as an experimental endotoxic shock model. Infect Immun1999; 67: 1018–1024.
17.
SassGKoerberKBangRGuehringHTiegsG. Inducible nitric oxide synthase is critical for immune-mediated liver injury in mice. J Clin Invest2001; 107: 439–447.
18.
ItoHKoideNMorikawaA, et al.. Augmentation of lipopolysaccharide-induced nitric oxide production by alpha-galactosylceramide in mouse peritoneal cells. J Endotoxin Res2005; 11: 213–219.
19.
MorikawaAKoideNKatoY, et al.. Augmentation of nitric oxide production by gamma interferon in a mouse vascular endothelial cell line and its modulation by tumor necrosis factor alpha and lipopolysaccharide. Infect Immun2000; 68: 6209–6214.
20.
ItoHKoideNHassanF, et al.. Lethal endotoxic shock using alpha-galactosylceramide sensitization as a new experimental model of septic shock. Lab Invest2006; 86: 254–261.
21.
GreenLCWagnerDAGlogowskiJSkipperPLWishnokJSTannenbaumSR. Analysis of nitrate, nitrite, and 15N nitrate in biological fluids. Anal Biochem1982; 126: 131–138.
22.
KoideNMuMMHassanF, et al.. Lipopolysaccharide enhances interferon-gamma-induced nitric oxide (NO) production in murine vascular endothelial cells via augmentation of interferon regulatory factor-1 activation. J Endotoxin Res2007; 13: 167–175.
23.
TumurkhuuGKoideNDagvadorjJ, et al.. The mechanism of development of acute lung injury in lethal endotoxic shock using alpha-galactosylceramide sensitization. Clin Exp Immunol2008; 152: 182–191.
24.
BahjatFRDharnidharkaVRFukuzukaK, et al.. Reduced susceptibility of nonobese diabetic mice to TNF-alpha and d-galactosamine-mediated hepatocellular apoptosis and lethality. J Immunol2000; 165: 6559–6567.
25.
KoideNNaritaKKatoY, et al.. Expression of Fas and Fas ligand on mouse renal tubular epithelial cells in the generalized Schwartzman reaction and its relationship to apoptosis. Infect Immun1999; 67: 4112–4118.
26.
HeremansHVan DammeJDillenCDijkmansRBilliauA. Interferon gamma, a mediator of lethal lipopolysaccharide-induced Shwartzman-like shock reactions in mice. J Exp Med1990; 171: 1853–1869.
27.
OzmenLPericinMHakimiJ, et al.. Interleukin 12, interferon gamma, and tumor necrosis factor alpha are the key cytokines of the generalized Shwartzman reaction. J Exp Med1994; 180: 907–915.
28.
CarBDEngVMSchnyderB, et al.. Interferon gamma receptor deficient mice are resistant to endotoxic shock. J Exp Med1994; 179: 1437–1444.
29.
KimWHHongFRadaevaSJarugaBFanSGaoB. STAT1 plays an essential role in LPS/D-galactosamine-induced liver apoptosis and injury. Am J Physiol Gastrointest Liver Physiol2003; 285: 761–768.
30.
HabuYShinomiyaNKinoshitaMMatsumotoAKawabataTSekiS. Mice deficient in Vbeta8+ NKT cells are resistant to experimental hepatitis but are partially susceptible to generalised Shwartzman reaction. Clin Exp Med2007; 7: 30–38.
31.
FujiiHSekiSKobayashiS, et al.. A murine model of NKT cell-mediated liver injury induced by alpha-galactosylceramide/d-galactosamine. Virchows Arch2005; 446: 663–673.
32.
PoultonLDSmythMJHawkeCG, et al.. Cytometric and functional analyses of NK and NKT cell deficiencies in NOD mice. Int Immunol2001; 13: 887–896.