
Abstract
Chronic low back pain (cLBP) is associated with insomnia and advanced age. Emerging evidence suggests that the severity of both sleep disorders (like insomnia) and chronic pain are associated with a faster pace of biological aging. We aimed to determine whether the pace of biological age mediates the relationship between insomnia and the impact of cLBP in a sample of community-dwelling adults ages 19 to 85 years. Participants (49 with no pain, 32 with low-impact pain, and 37 with high-impact pain) completed sociodemographic, pain, insomnia, and short physical performance battery assessments. We calculated the pace of biological aging using DunedinPACE from blood leukocyte DNA. On average, individuals with high-impact cLBP had significantly faster biological aging than those with low-impact and no chronic pain (p < .001). Bivariate associations of DunedinPACE scores with insomnia severity and functional performance were significant at p < .01 (rs = 0.324 and -0.502, respectively). After adjusting for race and sex, the association of insomnia severity and the impact of cLBP was partially mediated by the pace of biological aging (β = 0.070, p < .001). Also, the association of insomnia severity with functional performance was partially mediated by the pace of biological aging (β = -0.105, p < .001). Thus, insomnia remains strongly predictive of cLBP outcomes, and the pace of biological aging helps explain this association. Future prospective studies with repeated assessments are needed to uncover the directionality of these complex relationships and ultimately develop interventions to manage cLBP.
Keywords
Non-specific chronic low back pain (cLBP) is a well-documented and pervasive health problem among adults worldwide. 1 The underlying pathophysiology of cLBP points toward a complex interplay of biological and psychosocial factors.2–5 Besides the high economic cost to the healthcare system and society, cLBP negatively affects the individuals’ daily functioning, quality of life, and sleep.6,7 This is concerning because sleep plays an essential physiological protective and restorative homeostatic role in reducing vulnerability and promoting healthy aging. 8 Decades of research have shown a bidirectional relationship between sleep and pain, but more recently, it has been suggested that sleep may have a greater impact on the experience of chronic pain than vice versa.9,10 It has been suggested that poor sleep quality may be essential for the development and maintenance of chronic pain, and the prevalence of both insomnia and cLBP increases with age. 11 As a result, insomnia is becoming increasingly recognized as a clinically relevant comorbidity in individuals with cLBP. 12 Thus, understanding the relationship between insomnia and biological aging in the context of non-specific cLBP may enhance personalized pain care.
Advancing age is a strong predictor of cLBP. 13 However, unlike chronological aging, which refers only to the passage of time and is the same for everyone, biological aging reflects time-dependent variations in physiologic function resulting from changes in molecular and cellular functions, influencing disease risk. 14 Generally, the aging process is characterized by nine hallmark changes (genomic instability, telomere shortening, epigenetic modifications, loss of proteostasis, deregulated nutrient sensing, mitochondrial dysfunction, cellular senescence, stem cell exhaustion, and altered intercellular communication), 15 some of which are used to measure biological aging. For instance, DNA methylation (DNAm) is one of the most stable epigenetic changes associated with aging. Various DNAm clocks have been developed with predictive phenotypes such as chronological age,16,17 lifespan, and mortality. 18 Unfortunately, these clocks do not accurately predict the biological aging process. 19 Recently, 20 years of longitudinal data was used to develop the Dunedin Pace of Aging Calculated From the Epigenome (DunedinPACE), which provides a robust estimate of the rate of physiological decline. 19 We recently found that the pace of biological aging, as measured by DunedinPACE, highly predicts the odds of having cLBP compared to pain-free control (PFC) and correlates with non-specific cLBP severity and interference (UNDER REVIEW). The underlying mechanism explaining the relation between non-specific cLBP and the faster pace of biological aging still needs to be fully understood.
Growing evidence suggests overlapping mechanisms with sleep disturbances (e.g., insomnia) and cLBP.11,12,20 In 2011, a systematic review reported that insomnia is the most common sleep disturbance among individuals with cLBP. 21 More recently, a 2021 systematic review and meta-analysis reported a positive association between insomnia severity and cLBP. 22 Other studies have found that individuals with cLBP are 18 times more likely to experience insomnia than pain-free controls (PFCs). 23 Also, insomnia and sleep fragmentation are associated with accelerated epigenetic aging.8,24,25 However, the relationship between insomnia and the pace of biological aging has not been examined in individuals with non-specific cLBP. Examining this relationship is relevant because understanding the pace of biological aging is critical to developing effective therapeutic interventions to improve the quality of life and lifespan.
Given the essential role of sleep in physiological restoration and the relationship between insomnia, the pace of biological aging, and non-specific cLBP, an empirically driven study of the relationship between these variables may yield novel information concerning insomnia and non-specific cLBP. This study aimed to address this gap in our understanding by investigating how insomnia and the pace of biological aging affect cLBP. We hypothesize that the pace of biological aging mediates the relationship between insomnia and pain outcomes. Specifically, we examined whether more severe insomnia correlates with faster biological aging, which in turn correlates with greater pain intensity, pain-related disability, and poorer physical performance.
Methods
Participants
The study included 118 participants recruited from Birmingham, AL, USA, as part of ongoing projects examining the role of psychosocial and epigenomic differences between non-Hispanic Blacks and non-Hispanic Whites with cLBP. Details of the recruitment and study protocol have previously been published.2,26 All participants self-identified as non-Hispanic Black (African American) or non-Hispanic White (Caucasian), and data reported in this study was collected between June 2019 through May 2022.
Individuals could participate in the study if they were between 18 and 85 years, could understand English, and complete the questionnaires independently. A comprehensive list of inclusion and exclusion criteria has previously been published.2,26 Briefly, non-Hispanic Blacks and non-Hispanic Whites with non-specific cLBP were eligible to participate. We confirmed the diagnosis of non-specific cLBP through the medical record in accordance with the joint clinical practice guidelines from the American Colleges of Physicians and the American Pain Society. 27 Potential participants with any of the following criteria were excluded from the study: (1) low back pain attributable to infection, trauma, malignancy, or ankylosing spondylitis; (2) systemic infection; (3) chronic inflammatory disease (e.g., rheumatoid arthritis, systemic lupus erythematosus, fibromyalgia); (4) poorly controlled diabetes; (5) neurological disorders (e.g., Parkinson’s disease, multiple sclerosis, and epilepsy) and (6) severe psychiatric disease requiring hospitalization within the past 12 months. The same inclusion and exclusion criteria applied to PFC participants, except for the diagnosis of cLBP in the PFCs. All participants were instructed to continue their usual medications. This is because temporary withdrawal from these medications could have affected slepain perception.
All participants were carefully informed about the study and provided written informed consent before joining the study. The study was conducted following ethical guidelines and the protocol approved by the Institutional Review Board (IRB) at the University of Alabama at Birmingham. The study protocol adhered to the principles of the Declaration of Helsinki.
Measures
Eligible participants who met the inclusion criteria and provided informed consent completed several sociodemographic and pain questionnaires, including self-identified race, sex assigned at birth, household income after taxes, Insomnia Severity Index (ISI), Graded Chronic Pain Scale (GCPS), and Short Physical Performance Battery (SPPB). During the visit, a phlebotomist also collected venous blood for genotyping from each participant.
Short physical performance battery (SPPB)
The SPPB was used to assess lower extremity physical function (functional performance). The SPPB consists of three objective measures: balance, gait speed, and chair stands. 28 Each component is scored on a scale from 0 (worst performance) to 4 (best performance), with higher scores indicating better functional performance. We calculated the SPPB total score by summing the scores from each component. The total score ranged from 0 to 12, with a lower score indicating worse functional performance and a greater likelihood of disability. In our sample, the SPPB had an acceptable internal consistency reliability (Cronbach α = 0.81).
Graded chronic pain scale (GCPS)
The GCPS was employed to evaluate the intensity and impact of pain on participants’ daily functioning. The GCPS comprises two main components: pain intensity (items 1-3) and pain-related disability (items 4-6). 29 Participants reported their current, average, and worse pain in the past 6 months using a numerical rating scale ranging from 0 to 10, where 0 represents “no pain” and 10 corresponds to “worst possible pain”. The average of these ratings gave the pain intensity score. Similarly, participants rated (on a scale of 0 to 10) how much pain interfered with various aspects of daily life, including activity level, social activities, work, and enjoyment of life. The pain-related disability score was calculated as the average difficulties with activities of daily living.
Based on the pain intensity and pain-related disability scores, participants were assigned into one of three pain groups: no chronic pain, low impact chronic pain (mild or bothersome chronic pain), and high impact chronic pain. This classification was based on guidelines from the revised GCPS 29 and consistent with published works.30,31 The GCPS has previously been demonstrated to have good reliability and validity in various populations with chronic pain.32,33
Insomnia severity index (ISI)
The Insomnia Severity Index (ISI) was utilized to assess the severity and impact of insomnia symptoms. The ISI is a brief self-report questionnaire consisting of 7 items that measure the perceived severity of insomnia symptoms such as difficulty falling asleep, maintaining sleep, and early morning awakening. 34 Using a scale from 0 to 4, each participant rated the severity of each symptom over the last month. We summed each item to calculate a total score that ranged from 0 to 28, with higher scores indicating greater insomnia severity. The ISI has previously been demonstrated to have good reliability and validity in various populations. 35
DNA methylation analysis
Venous blood was collected from each participant into EDTA tubes and subsequently used for DNA extraction and methylation analysis. Genomic DNA was extracted using the Gentra Puregene DNA Purification Protocol (Qiagen, Valencia, CA, USA). Details of the DNA extraction have previously been published. 5 Extracted DNA was shipped on dry ice to the University of Minnesota Genomic Center, where sodium conversion and DNA methylation analysis was done using the Illumina Infinium MethylationEPIC BeadChip protocol. The MethylationEPIC platform identifies the methylation level in over 850,000 CpG sites (Illumina, San Diego, CA, USA).
Pace of biological aging calculation
Raw methylome data from the Illumina EPIC array (.idat files) was processed using commonly used R packages, including minfi36,37 and Enmix. 38 Following preprocessing and normalization, none of the samples were removed because of low quality (e.g., bisulfate intensity <4750, low-quality methylation values, or outliers for their methylation β-value distributions). Principal component analysis (PCA) was used to estimate the variance explained by circulating proportions of CD8 T lymphocytes, CD4 T lymphocytes, natural killer cells, B cells, monocytes, and granulocytes.
We used the DunedinPACE algorithm in the methylAge module of Enmix R package on Bioconductor to estimate each participant’s biological aging pace (DunedinPACE scores). 19 The DunedinPACE scores are always positive; scores > 1 indicate a faster pace of biological aging, and scores < 1 indicate a slower pace of biological aging.
Statistical analysis
After calculating DunedinPACE scores, all statistical analyses were conducted using SPSS version 29.0 (IBM; Armonk, New York). The demographic and pain-related characteristics of participants were calculated separately by pain impact classification and overall study sample. Bivariate correlations among variables were examined using Spearman’s Rho test. Next, regression-based mediation analyses were performed with PROCESS macro (model 4) to determine whether DunedinPACE scores mediated the relationship between insomnia and the impact of cLBP, with insomnia as the explanatory variable (X), DunedinPACE scores as the mediator (M), and pain impact classification as the outcome or dependent variable (Y). Additionally, the same model was used to evaluate whether DunedinPACE scores mediate the relationship between insomnia and functional performance (SPPB). We included race and sex as covariates in both models, and p-values of < 0.05 were considered significant.
Results
Characteristics of participants
Characteristics of participants and study variables.

Notes. * denotes direction of the within group significant differences based on Scheffe’s Post Hoc Analysis; sd = Standard deviation; HI = high impact group; LI = low impact group; NP = no pain; SPPB = short physical performance battery.

Box plot of differences in DunedinPACE score, functional performance, and insomnia divided by pain impact groups (no pain, low impact pain, and high impact pain). Within each box, the horizontal line in the middle is the median value of the scores, and the lower and upper boundaries indicate the 25th and 75th percentiles, respectively. SPPB = short physical performance battery.
Bivariate correlations
Descriptive Statistics and correlations of study variables.
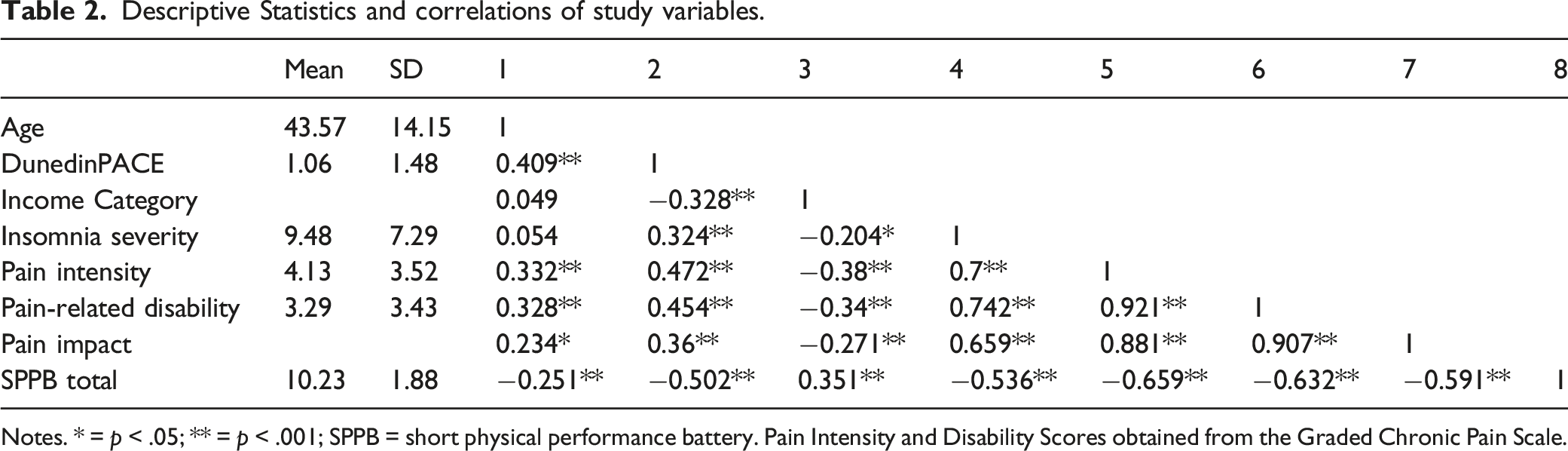
Notes. * = p < .05; ** = p < .001; SPPB = short physical performance battery. Pain Intensity and Disability Scores obtained from the Graded Chronic Pain Scale.
DunedinPACE mediates the association between insomnia and pain outcomes
We used mediation analyses to test whether the pace of biological aging (M) could explain the relationship between insomnia (X) and both intensity and pain-related disability (Y). We included race and sex as covariates in the mediation model. As shown in Figure 2, insomnia severity was positively associated with DunedinPACE scores (β = 0.006 (SE = 0.002), p = .001, R2 = 0.23) and pain intensity (β = 0.286 (SE = 0.034), p < .001). The relationship between DunedinPACE and pain intensity was significant (β = 5.493, (SE = 1.744), p = .002). The model with race, sex, insomnia severity, and DunedinPACE explained most of the variance in pain intensity (r = 0.734, p < .001). The total effect of insomnia severity reveals that a unit increase in insomnia severity positively correlates with a 0.32 increase in pain intensity (β = 0.320 (SE =0.034), 95% CI = 0.253; 0.387, p < .001). The bias-corrected bootstrap 95% CI for the total indirect effect based on 5,000 bootstrap samples did not include zero (β = 0.034 (SE = 0.016), Boots 95% CI = 0.008; 0.071), confirming that DunedinPACE partially mediates the influence of insomnia on pain intensity. A simple mediation model of the indirect relationship between insomnia severity and pain intensity through DunedinPACE, controlling for chronological age and sex. Coefficients presented (a, b, c, and c’) are unstandardized regression coefficients. *p < .05 **p < 01. Path c indicates the total effect, and path c’ indicates the direct effect, and a*b indicates the indirect effect.
Additionally, the indirect effect of insomnia on pain-related disability through DunedinPACE was statistically significant after controlling for race and sex (β = 0.032 (SE = 0.015), Boots 95% CI = 0.008; 0.066). Figure 3 depicts the schematic of the mediation model, including insomnia severity (X), DunedinPACE (M), and pain-related disability. Mediation Model of Associations Between Insomnia Severity and Pain-related Disability Through DunedinPACE scores, controlling for chronological age and sex. Coefficients presented (a, b, c, and c’) are unstandardized regression coefficients. *p < .05 **p < 01. Path c indicates the total effect, and path c’ indicates the direct effect, and a*b indicates the indirect effect.
DunedinPACE mediates the association between insomnia and functional performance
We also tested whether the pace of biological aging could explain the association between insomnia and SPPB scores. Age and sex were entered as covariates in the model. As depicted in Figure 4, insomnia severity was negatively associated with SPPB total scores (β = -0.105 (SE = 0.020), p < .001, R2 = 0.37). The total effect of insomnia severity on SPPB total scores revealed that a unit increase in insomnia severity decreased SPPB total scores by 0.125 units (β = -0.125 (SE = 0.020), 95% CI = -0.165 to -0.085, p < .001). The bias-corrected bootstrap 95% CI for the indirect effect DunedinPACE based on 5000 samples did not include zero (β = -0.020 (SE = 0.009), Boots 95% CI = -0.039 to -0.005). Supporting our hypothesis that greater insomnia severity correlated with lower functional performance, and the pace of biological aging partially mediates that relationship. Mediation model of the associations between insomnia severity and functional performance through DunedinPACE scores, for chronological age and sex. Coefficients presented (a, b, c, and c’) are unstandardized regression coefficients. *p < .05 **p < 01. Path c indicates the total effect, and path c’ indicates the direct effect, and a*b indicates the indirect effect.
Discussion
Pain and sleep are essential for normal physiological functions in humans, and poor sleep is critical for developing and maintaining chronic pain. 11 The current study is the first to investigate how insomnia and the pace of biological aging relate to non-specific cLBP. We found that insomnia severity positively correlates with both pain intensity and pain-related disability, and the pace of biological aging partially mediates these relationships. Specifically, individuals with more severe levels of insomnia had higher DunedinPACE scores, which in turn correlated with greater pain intensity and disability. The indirect relationship between insomnia and pain (via DunedinPACE) remained significant after controlling for the sex and race of the participants. These findings are consistent with other studies showing that short sleep durations and insomnia are associated with epigenetic age acceleration and faster aging in adults. 24 Additional studies have also reported a significant correlation between chronic low back pain and insomnia severity,20,22 corroborating the results revealed in this study.
One potential molecular mechanism by which insomnia increases the risk for cLBP severity and related disability is epigenetically mediated alterations of inflammatory gene expressions. Both insomnia and cLBP have been associated with inflammatory markers such as interleukin-6, C-reactive protein, and nitric oxide. 12 Additionally, a faster pace of aging has been associated with the accumulation of inflammatory markers. In fact, most epigenetic clocks correlate with DNA methylation changes in inflammatory markers.39,40 Emerging evidence suggest that more severe insomnia symptoms correlate with epigenetic age acceleration 25 and telomere shortening. 41 Other investigators have linked epigenetic age acceleration with chronic musculoskeletal pain such as knee osteoarthritis.42–44 Given that DNA methylation alterations of stress-related inflammatory process correlate with cLBP5,45,46 and insomnia, 47 it is possible that insufficient sleep quality that characterizes insomnia epigenetically induces inflammatory processes that accelerate aging, sustaining worse cLBP.
Other mechanisms have previously been proposed to explain the relationship between cLBP and insomnia, including the impact of pain on physical health, movement, as well as depressive symptoms. 48 Our study found that insomnia severity correlated with functional performance as measured by the SPPB, suggesting that individuals with more severe insomnia had worse physical health. We have previously reported that worse physical performance is positively associated with depressive symptoms, perceived injustice, internalized stigma, and response to experimentally evoked pain stimuli (temporal summation).26,49 Other investigators have suggested that increased cLBP symptoms in adults with insomnia may be related to altered pain modulation and central sensitization. 50 However, the molecular mechanism linking insomnia, central sensitization, and worse functional performance in cLBP remains under-explored.
There are several methodological limitations to our study. First, the findings of this cross-sectional study must be replicated in a prospective longitudinal design to determine causation. However, our mediation analyses suggest that insomnia may increase the impact of cLBP by accelerating the pace of biological aging. Should replication and prospective designs support these initial findings, this raises the possibility that interventions to improve sleep quality may slow the aging process and improve cLBP outcomes. Second, mediation analysis assumes a unidirectional relationship from insomnia to pain impact through DunedinPACE (i.e., insomnia → DunedinPACE → pain). While this directionality is biologically plausible, an alternative hypothesis could be that insomnia leads to high-impact cLBP, which results in faster aging (i.e., insomnia → pain → DunedinPACE). Future prospective studies with repeated assessments may uncover the directionality of these complex relationships. Thirds, the current sample comprised of self-identified non-Hispanic Blacks and non-Hispanic Whites only and is not representative of the population of the United States or adults with cLBP. Finally, we used blood samples to determine the pace of biological aging. We did not see any association between DunedinPACE and various cell compositions in our principal component analysis. Future studies should explore whether insomnia and the impact of cLBP are associated with the pace of aging in other tissues.
Despite these limitations, several strengths should be noted, particularly the rigorous quality of the measurements, sample collection, and bioinformatic processes. This study is the first to examine the complex relationship between insomnia, the pace of aging, and the impact of non-specific cLBP, providing a proof-of-concept. Another notable strength is the use of mediation analysis, which can inform future studies to establish causation.
Conclusion
Insomnia is very frequent in adults with cLBP, and the prevalence of both insomnia and cLBP increases with age. Our study aimed to examine how insomnia and the pace of biological aging correlate with cLBP intensity and disability. Based on our current knowledge, insomnia severity positively correlates with cLBP intensity, disability, and worse functional performance. Furthermore, the pace of biological aging mediates the relationship between insomnia severity and cLBP (pain intensity, pain-related disability, and functional performance), such that individuals with more severe insomnia have faster aging that correlates with high-impact pain and worse functional performance.
Footnotes
Declarations
The funder had no role in study design, data collection and analysis, the decision to publish, or the preparation of the manuscript.
Author contributions
ENA and BRG contributed to the conception and design. ENA, RES, PK, VS, and HKT helped in the experimental part, analysis, and interpretation of data. ENA, BRG, AMW, and TQ prepared the manuscript and performed additional analysis for revision. TQ, AMW, and JB collected the sample and maintained the patient database. ENA and BRG are the principal investigators of the project. All authors read and approved the final manuscript.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was funded by grants from the National Institute on Minority Health and Health Disparities R01MD010441 and R01MD017565), National Institute of Nursing Research (R01MD017565), and National Institute of Arthritis and Musculoskeletal and Skin Diseases (R01AR079178).