
Abstract
Introduction
Stress has been previously defined as a stimulus (stressor) that causes a reaction in the brain that, in turn, activates a physiological response. 1 Previous literature has shown that stress on the body can affect the function and distribution of immune cells.1–5 In humans, psychosocial stress can elicit an inflammatory response, suggesting activation of the immune system. 6 Whereas it was once believed that acute stress suppressed inflammation, it is now known that stress can also increase the production of proinflammatory signals.7,8 Furthermore, sustained stress and inflammation can result in elevated levels of reactive oxygen species (ROS) and damage-associated molecular patterns (DAMPs).9,10 DAMPs stimulate an immune/inflammatory response and serve to maintain and restore homeostasis. However, DAMPs have been shown to initiate the posttraumatic immune response, which is normally induced by foreign bodies in order to activate and recruit cells of the immune system.11–13 Additionally, trauma can activate the release of DAMPs from dying cells. 14 These signals can have negative impacts on many systems and may contribute to mitochondrial dysfunction. 15
Whereas inflammation can lead to mitochondrial damage, the reverse is also true. Mitochondria generate or contain several types of potential mitochondrial DAMPs (mDAMPs) such as mitochondrial DNA (mtDNA).16,17 Each serves as a danger signal for the immune system and can function as drivers of inflammation.16,18 Extracellular mtDNA bind to Toll-like receptor 9 (TLR9), which, in macrophages causes the production of proinflammatory cytokines. 19 Proinflammatory cytokines have been shown to promote the generation of ROS in the mitochondria. 20 During acute and chronic inflammation, the production of ROS exceeds the capacity of the defense system’s ability to remove them, which in turn can lead to increased mitochondrial dysfunction as well as the release of mDAMPs. 21 Chronic mitochondrial dysfunction has been seen in a number of degenerative diseases, including Huntington’s, Alzheimer’s, Parkinson’s and Amyotrophic Lateral Sclerosis. 22 Sadly, for those suffering from the Human Immunodeficiency Virus (HIV), it is believed that both the HIV itself and many of the antiretroviral therapies (ART) used to treat HIV may contribute to mitochondrial dysfunction 23 and consequently result in proinflamamtory response.
HIV infection is characterized by a progressive depletion of CD4+ T-cell populations and a state of chronic inflammation and immune activation. As such, people with HIV (PWH) have a significantly higher chance of developing inflammatory-related comorbidities compared to older non-HIV+ individuals. 24 A mechanism implicated in the pathogenesis of HIV disease and its comorbidities is a pro-oxidative status associated with the infection and with ART. Previous literature shows that HIV infection induces oxidative stress by (1) deregulating pathways of oxidative stress, (2) increasing ROS production, and (3) inducing mitochondrial dysfunction.25–27 Activation of inflammatory pathways within the peripheral immune cells, acts as an adaptive response to the HIV-1 virus infections but could alter afferent nociceptive signaling, playing a role in the development of pain-related syndromes and hypersensitivity in PWH. 28
Chronic pain is often comorbid in those with HIV. Studies suggest that the percentage of PWH who suffer from some form of chronic pain throughout their lives varies from 25% to 90% with their chances increasing with age.28–30 Chronic pain concurrent with HIV includes neuropathic, inflammatory, and widespread musculoskeletal pain.29–32 Immune cells have been shown to contribute to all types of pain.33–36 Additionally, there is evidence to suggest that there is a relationship present between oxidative stress and one’s perception of pain as well as inflammation. 37 Physical stressors cause a physiological response that can increase oxidative stress and contribute to mitochondrial damage. A stressor can be any stimuli that evokes a physiological stress response and can be pain or non-pain related. 38 It is important to note that pain serves as a form of stressor. 38 Additionally, a maladaptive response experienced during acute pain may serve to intensify the pain that is experienced as well as condition a sensitized physiological stress response to a stimuli that provokes pain.39,40 For our study, we used Quantitative Sensory Testing (QST) as an acute physical stressor.
One possibility underlying the comorbidity of HIV and chronic pain is that the physiological response to stressors is enhanced, leading to greater mitochondrial damage and the development/maintenance of chronic pain. It is unknown whether an acute stressor (quantitative sensory testing; QST) would result in measurable changes in mitochondrial damage markers in PWH and whether comorbid chronic pain would exacerbate the effect. Therefore, we sought to examine whether PWH with/without chronic pain showed differential mitochondrial physiological responses to acute painful stimuli. We hypothesize the following: (1) PWH and chronic pain will have a statistically significant difference overall in their mitochondrial biomarkers following exposure to QST than PWH and no pain; (2) Specifically, PWH and chronic pain would have lower mitochondrial copy number (i.e., content), higher mitochondrial DNA damage, and higher levels of mitochondrial DNA DAMPs circulating in the blood in comparison to PWH and no pain, following exposure to QST.
Methods
The current study was a part of a larger parent study with the goal of investigating biobehavioral risk factors that are associated with HIV and chronic pain (NCT #04298658). The parent study collected data including self-report questionnaires, daily diaries, QST, and various biomarkers during one study visit. More details of the findings and procedures from this study can be found elsewhere.41–44 However, none of these published studies overlap with the current study.
Participants
All study participants were HIV-positive. Participants were recruited from Birmingham, Alabama via flyers, including the University of Alabama at Birmingham (UAB) 1917 Clinic, which provides care for PWH. Interested participants were instructed to call in order to join the study and eligibility was determined. Inclusion criteria included: 18 years or older and current HIV infection. High blood pressure (150/95 mmHg, systolic/diastolic) was an exclusion criteria as high blood pressure could lead to adverse effects with some of the procedures conducted during the study. Chronic pain was classified as having pain that lasted for 6 months, at least half of the days. After enrollment, participants were provided with a unique identification number for use throughout the study. No identifying information was maintained in the dataset. A total of 55 participants were enrolled in the study.
Following determination of eligibility, participants came to the testing facility and completed a single session that lasted approximately 180 min (Figure 1). After completion of informed consent, blood pressure was taken in addition to body temperature, height, and weight. Next, a nurse inserted an intravenous (IV) cannula into the antecubital fossa of the non-dominant arm in order to allow for a total of two blood draws across the session. Lastly, the participants completed QST. To not exacerbate any pre-existing conditions for our chronic pain participants, QST stimulation was performed on the opposite side of the body if pain was present at the location of the stimulus application. Timeline of session. The above figure illustrates the matriculation through the study session. QST tasks listed in timeline are heat pain, punctate, pressure, conditioned pain modulation, and cold pressor task. Heat pain represents all thermal tasks (i.e., threshold, tolerance and temporal summation). Pressure tasks refers to baseline pressure pain threshold task. Punctate refers to the punctate mechanical stimuli task.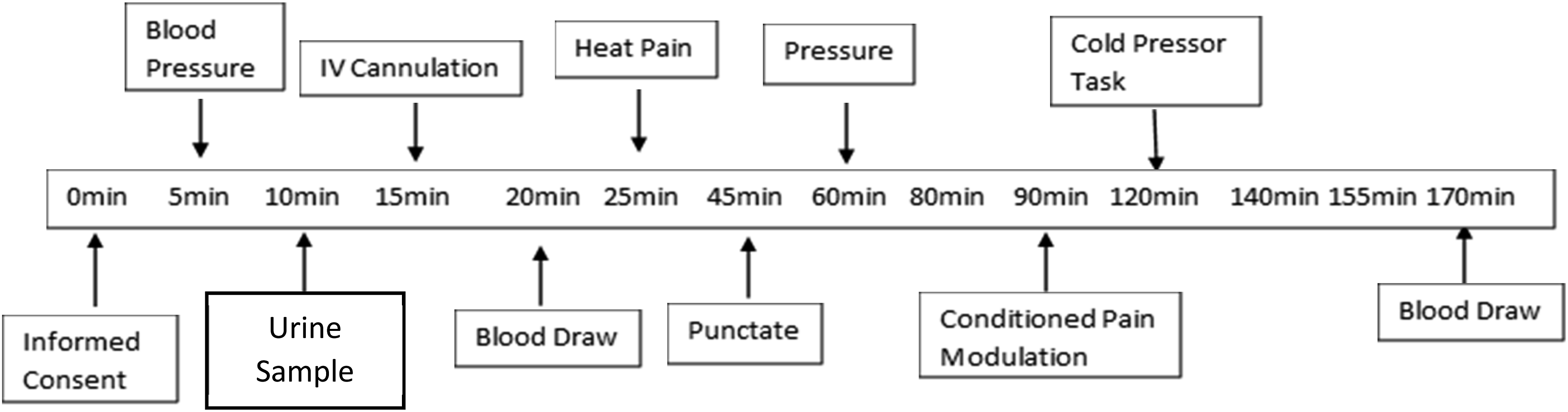
Medical record review
A Medical Record Review was completed in order to confirm prescribed medications and diagnoses. Medical records were also used in order to obtain the participants’ most recent CD4 T-cell count, a measure of their immune status, and the value of their recent viral load.
Questionnaires
Demographics questionnaire
A demographics questionnaire was completed in order to collect general participant background information. It was utilized to obtain the participants’ self-reported race, sex, ethnicity, and age.
Brief pain inventory-short form
The Brief pain inventory-short form (BPI-SF) is a nine item self-reported questionnaire that is used to examine the severity of a participant’s pain as well as the impact of the pain on the daily functioning of the participant. The BPI-SF is a modified version of the Brief Pain Inventory. 45 The questionnaire asked participants to rate their pain intensity during their worst, least, average, and current pain. It also examines the participants’ current medications and their effectiveness as well as the degree that their current pain interferes with their daily lives (i.e., mood, general activity, relationships, sleep, etc.). BPI-SF was shown to be a valid and reliable questionnaire.46–53
Quantitative sensory testing
Pain intensity ratings
The participants’ pain experience during QST was measured using the intensity of the painful sensation. The rating is measured on a scale from 0 to 100, with 0 considered no pain sensation and 100 reflecting the most intense pain imaginable. The participants provided ratings multiple times throughout the study.
Heat pain testing
Thermal pain task – threshold and tolerance: The thermal pain task was carried out via a computer-controlled Advanced Thermosensory Stimulator (TSA) using Medoc Software (Medoc, Advanced Medical Systems, Israel). The TSA thermode was placed in three separate spots on the ventral forearm in order to avoid the participant becoming sensitized or habituated to the stimuli. 54 The thermal pain task assessed two aspects: threshold and tolerance. The task was repeated three times for both threshold and tolerance in order to obtain an average reading. The first assessment was heat pain threshold. The baseline temperature of the thermode for each trial was 33°C and slowly increased to 50.5°C (approximately 1°C a second). The participant was then instructed, as a response to the stimulus, to press a button as soon as the stimulus first became painful. The second assessment, after threshold, was heat pain tolerance. Participants were then instructed to press a button when the pain became intolerable. Both the threshold and tolerance were repeated a total of 3 times and for analysis, the mean temperature for each measure was used.
Temporal summation (TS) heat pain task: Following the Thermal Pain Task, assessment, the participants took a two-minute break before going through a second thermal procedure. TS Heat Pain task was then assessed. The participants were then instructed to rate the intensity of their pain on a scale of 0 – 100, when each 5 repetitive, suprathreshold heat pulses reached their peak. Target temperatures were delivered using a computer-controlled TSA with the Medoc Software. The stimulus was applied on the dorsal forearm at 44°C, as a practice to show the participant how fast the peaks occurred. The remaining two trials were at two separate sites on the ventral forearm at 46 and 48°C. If the participant rated the task as “100”, it was immediately terminated. The average of the five peaks per TS temperature were used in analysis.
Punctate mechanical stimuli task
The Punctate Mechanical Stimuli Task used a fine nylon (300 g (6.65)) monofilament in order to test the participant’s sensitivity to fine pressure at two separate skin sites: the back of the non-dominant hand and the trapezius on the non-dominant side. Fine pressure was applied to the two separate sites by placing the monofilament gently on the skin without jabbing. After contact was made with the skin, pressure was applied until the tip of the monofilament bent slightly and held in place for 1 s before moving. For this study, we will refer to this action as tapping. The task began by tapping the site once and obtaining a rating of pain intensity. The task was then repeated, by tapping the site 10 times in a row (approximately a tap per every 2 seconds) and asking the participant to rate the highest level of pain they experienced during the trial. The task was repeated two times at each site. During the task, the participant was asked to close their eyes to prevent participant bias from seeing the pressure being applied.
Baseline pressure pain threshold task
Baseline Pressure Pain Threshold Task (PPT) was evaluated at two sites: the dominant forearm and ipsilateral trapezius. For both sites, a handheld pressure algometer (AlgoMed-Digital Algometer, Medoc, Advanced Medical Systems, Israel) and computer software (Medoc, Advanced Medical Systems) were used to assess the baseline pressure. Pressure was applied to the two sites at a constant rate (approximately 33 kPa per second) using a rubber tip probe with a diameter of 10 mm. The participant was asked to press a button when the pressure stimulus first became painful. The amount of pressure, measured in kPa, was recorded. The task was repeated three times for each testing site to create an average pressure pain threshold to be used for analysis. The maximum pressure applied on the two sites were 1000 kPa. If the participant did not report any pain before or at the maximum pressure applied, then the procedure was terminated and a score of 1000 kPa was assigned to the trial.
Cold pressor task
The Cold Pressor Task (CPT) involved the participant placing their non-dominant hand, up to their wrist, into cold water for up to 5 min. The participant was asked to keep their hand open and to avoid contact with the sides or bottom of the tank. Every 30 s, the participant was asked to rate the pain intensity in their submerged hand. The water temperature was maintained at 10°C (Thermo Fisher Scientific Haake SC150, USA). An average of the pain ratings was used for analysis.
Conditioned pain modulation task
The Conditioned Pain Modulation (CPM) Task serves as a marker for the endogenous inhibition of pain. 54 The CPT served as the conditioning stimulus for the CPM Task with a maintained temperature of 10°C. Pressure pain served as the testing stimulus and was applied to both the ipsilateral trapezius and dominant forearm. A total of four trials was conducted, twice at each site. During the cold immersion, the participant was instructed to place their non-dominant hand in the cold water for up to minute. They were asked to rate their pain intensity after 30 s. After a minute, or if their pain reached 100, the participant removed their hand from the water. Immediately, after the hand was removed from the cold water, the pressure stimulus was administered to one of the two sites. The participant was then instructed to press a button when they first felt pain at the pressure pain site. Following the pressure stimulus was a two-minute break before repeating the task and switching the pressure pain site. The CPM measure was calculated by subtracting the PPT when tested alone from the PPT when tested immediately following the CPT.
Mitochondrial reactivity measures
Mitochondrial DNA copy number
The following methods were previously described.55–63 Briefly, total DNA was isolated using the QIAamp DNA Mini Kit (Qiagen, Hilden, Germany) and quantified using the Quant-iT PicoGreen dsDNA Assay Kit (Invitrogen, Carlsbad, CA). A 236 bp (short) fragment of the mitochondrial genome was amplified from 15 ng of total DNA, in a GeneAmp PCR system 2400 using a quantitative polymerase chain reaction (QPCR), GoTaq DNA Polymerase (Promega, USA), 32P radiolabeled ATP (Perkin-Elmer, USA) and primers 1066F/N1007R. 20 µL of each QPCR product was resolved (vertical electrophoresis) on 8%–12% polyacrylamide gels at 90 volts for 2 h. Dried gels were exposed to phosphor screens for 1–2 h and quantified with IMAGEQUANT (Molecular Dynamics Inc., USA). Quantified bands were normalized to the control and mtDNA copy number (content) reported as a percentage of positive control. MtDNA copy number is used as an approximation of mitochondrial content. A higher number of mtDNA copy number is indicative of higher mitochondrial content.
Mitochondrial DNA damage
The entire mitochondrial genome (long) was then amplified by QPCR from 15 ng of total DNA, using AccuPrime Taq DNA Polymerase (Invitrogen, USA), 32P radiolabeled ATP (Perkin-Elmer, USA) and primers 1065F/1066R.55–63 25 µL of each “long” QPCR product was resolved (vertical electrophoresis) on 1.2% agarose gels at 90 volts for 3 h, dried and exposed to phosphor screens for 12 h, before being quantified with IMAGEQUANT (Molecular Dynamics Inc., USA). Quantified bands were normalized to the positive control. Each long was normalized to the short PCR product and these values were used to calculate the average mtDNA lesion frequency for the sample using the Poisson equation: –ln (treated/untreated sample). 56 The lesion frequency variable was used as a measure of mtDNA damage with lower scores representing greater mtDNA damage.
Mitochondrial DNA damage associated molecular pattern assay
The following methodology was previously described by Butts and associates. 64 mDAMPs were in assessed in serum from human subjects. Briefly, cell-free mitochondrial DNA (cfmtDNA) was extracted from 100 µl of serum using a MagMax™ Cell-Free DNA Isolation Kit (Applied Biosystems, USA) following the manufacturer’s instructions with minor adaptations (2.5 µL of the MagMax™ Cell-Free DNA Magnetic Beads per sample was used instead of 5 µL per sample). CfDNA was eluted in 20 µl volumes and aliquots were stored at −80°C. mDAMPs were assessed via amplification of sequences within the NADH dehydrogenase subunit 1 (ND1) and NADH dehydrogenase subunit 6 (ND6) regions of the mtDNA by Real-Time polymerase chain reaction (PCR) as previously described65,66 with minor modifications using a StepOne Plus Real-Time PCR system (Thermo Fisher Scientific, USA). The mDAMPs assay provided two variables to represent markers of damage: ND1 and ND6.
Data analysis
Chi-square was used to examine relationships between categorical variables. The inter-relationships among continuous study variables were evaluated using Pearson-product moment correlations. One-Way ANOVA were used to examine mean differences between pain groups across the baseline mitochondrial biomarkers (i.e. mtDNA copy number, mDAMPs, and mtDNA damage). Change scores of mtDNA copy number and mDAMPs (i.e. ND1 and ND6) along with mtDNA damage (i.e., lesion frequency) were evaluated using a Wilks’ Lambda Multivariate Analysis of Covariance (MANCOVA) while controlling for age, race, sex, and ethnicity to test our two hypotheses. The relationships between BPI-SF, pain reports, and the summary indicators of mitochondrial reactivity to pain were assessed using Pearson correlations. Data were analyzed using SPSS Statistical Software version 25 (International Business Machine Corporation, IBM).
Results
Participant characteristics
Demographics of PWH with and without chronic pain.

Numbers in the table are mean with standard deviations or percentages. For comparisons between chronic pain and no pain, chi-square test was used for categorical variables and t test for continuous.
Mean differences at baseline
There were no statistically significant differences between pain groups across each of the mitochondrial biomarkers at baseline: ND1 (F (1,46) = 0.292, p = .592), ND6 (F (1,45) = 0.281, p = .599), mtDNA copy number (F (1,46) = 2.235, p = .142), and mtDNA damage (F (1,46) = 0.267, p = .608).
Findings of MANCOVA
Differences in reactivity levels following pain testing
There was a statistically significant difference between PWH with and without pain on the various indicators of mitochondrial reactivity (i.e., mtDNA damage, copy number, ND1, and ND6) following acute pain testing, after controlling for age, race, sex, and ethnicity, (F (4,30) = 4.368, p = .007, Wilks’ = 0.632). Therefore, there is a statistically significant difference between the pain groups in terms of the combined mitochondrial biomarkers after controlling for age, race, sex, and ethnicity.
Differences in reactivity between groups
Mitochondrial biomarker differences between pain groups.

Numbers in the table are means with standard deviations. The mtDNA damage numbers represent the normalized differences between before and after exposure to quantitative sensory testing (QST).

Differences in ND6 levels between chronic pain and no pain. The above figure presents mean change scores for ND6 levels (copies per microliter of plasma) for PWH with chronic pain and without pain. * represents p < .05.

Differences in mtDNA damage levels between chronic pain and no pain. The above figure presents mean change scores for mtDNA damage levels (lesion frequency) for PWH with chronic pain and without pain. ** represents p < .01.
Correlations
Bivariate correlations of physiological reactivity variables.

aRepresents p < .05.
Note: Pain status coded as 1 = no chronic pain and 2 = with chronic pain. CPM = conditioned pain modulation (percent change from baseline).
Discussion
The results of the current study suggest that exposure to an acute stressor (QST) elicits a change in markers related to mitochondrial damage. Specifically, ND6 and mtDNA damage were the primary mitochondrial biomarkers shown to be significantly different between the groups (i.e., chronic pain and no pain) following QST. Our main finding shows that there is significant mitochondrial reactivity in PWH with pain following an acute physical stressor. For the collection of ND1 and ND6 we extracted cell-free mtDNA that was circulating in blood. Circulating cell-free mtDNA has been used as a potential indicator of mitochondrial health in a multitude of diseases such as brain diseases, inflammatory diseases and cancer. A study conducted by Zhang and associates, examining rats who were subjected to trauma and hemorrhagic shock (T/HS), found that rats who were exposed to T/HS have elevated mtDNA in their blood following exposure as compared to the naïve rats. 67 Similarly, we found greater change scores for ND6 following exposure to QST within our PWH and pain population. Importantly, there were no significant differences between pain groups across the mitochondrial biomarkers at baseline, suggesting that the reaction to the acute stressors (QST) were the primary distinction between groups. These results differ from previous literature that examines knee osteoarthritis (OA). Patients with knee osteoarthritis were found to have higher levels of mtDNA copy number and DAMPs in comparison to healthy controls.68–70
This study provided novel results about mitochondrial dysfunction, pertaining to mtDNA damage and mDAMPs in PWH. Additionally, there was an association between chronic pain and ND6 levels. However, it is important to note that there are several possible mechanisms that could be responsible for the mitochondrial reactivity (i.e. elevated ND6 and mtDNA damage) differences that are observed. One possible mechanism is the age difference between PWH with chronic pain and those without pain. Previous literature has shown that, as individuals age, there is a loss of tissue as well as organ function progressively over time. 71 The Free Radical Theory of Aging hypothesizes that functional losses due to aging are caused by the accumulation of oxidative stress created by both reactive oxygen and nitrogen species (RONS). The oxidative stress causes damage to macromolecules (i.e., DNA, proteins, etc.). 72 It is theorized that increased levels of RONS can lead to cellular senescence, which is a mechanism that stops the proliferation of cells as a response to damages that occur during the replication process. 73 Over time, this can lead to the development of age-related diseases. For example, the individuals in our chronic pain group were older by 5 years in comparison to our no pain group and had higher levels of ND6 change scores and lesion frequency (i.e., mitochondrial damage). Additionally, our older participants tended to have more comorbidities listed in their medical record review.
Another potential mechanism that could be responsible for the observed mitochondrial reactivity differences is the increase in passive cell death in PWH. HIV has a devastating effect on the immune system because it results in the infection and death of immune-regulating cells (i.e., CD4+ T lymphocytes), specifically through apoptosis. 74 Apoptosis is regulated cell death that occurs without the loss of the plasma membrane. 75 Additionally, DAMPs can be released during cell death, such as apoptosis. Traditionally, apoptosis was considered a non-immunogenic form of cell death. However, recent literature has shown that apoptosis can also be immunogenic when the body is experiencing stressful conditions. Immunogenic apoptosis is characterized by the release of DAMPs. 76 This mechanism has also shown that cell-free mtDNA as well as ATP are released during apoptosis.77,78 Different types of DAMPs can be released at different stages within during specific types of cell death. For example, ATP tends to be released during the pre-apoptotic stage whereas high mobility group box 1 (HMGB1) is released during the late stages of apoptosis.79,80 For PWH, they may have higher levels in comparison to healthy individuals since their disease results in increased programmed death. This could potentially explain the differences seen between our results and previous literature. However, we did not find significant differences in mtDNA copy number across groups at baseline or after exposure to the QST, as might be expected with older adults.81,82 Once again, these data suggest that the comorbid pain condition increased the responsivity to the acute stressors (QST).
Conclusions
Here, we show that the combination of HIV and chronic pain resulted in greater change in markers of mitochondrial dysfunction over the experimental timeline. Previous literature has shown that both HIV and pain can contribute to the presence of oxidative stress within the body and that both conditions are associated with mitochondrial damage.25–27,37 The present study sought to determine whether acute physical stressors would impact mitochondrial physiological responses in PWH and whether the added presence of chronic pain would enhance the reaction. Overall, there was a significant difference in mitochondrial reactivity levels following QST battery testing within the sample for those with chronic pain. Specifically, ND6 and mtDNA damage were shown to be significantly changed following the QST battery in PWH and chronic pain. It is possible that the high comorbidity of HIV and chronic pain is a result of the mitochondrial dysfunction due to HIV/ART that increases susceptibility to chronic pain. While we believe the minor (but significant) elevations in ND6 and mtDNA damage in PWH with chronic pain are transient and are unlikely to have long-term consequences, the data suggest that the comorbid conditions increase the physiological response to acute physical stressors at the level of the mitochondria. As such, persistent or repeated exposure to stressors is likely to exacerbate mitochondrial dysfunction and consequently enhance biological aging. Thus, further understanding of mitochondrial dysfunction in HIV and chronic pain is needed to facilitate development of effective therapies and reduce premature aging is this undertreated population.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Center for Advancing Translational Sciences (UL1TR003096), National Institute of Diabetes and Digestive and Kidney Diseases (P30DK056336, P30DK079626), National Heart, Lung, and Blood Institute (R01HL147603).